. Introduction
The culinary popularity and high nutritional value of chili peppers (Capsicum annuum L.) are driving increasing demand worldwide. Chili peppers include phenolic acids, antioxidants, vitamins, carotenoids, and flavonoids (Anolisa et al., 2020; FAOSTAT, 2023; Hernández‐Pérez et al., 2020). There will be serious problems with food security and environmental sustainability due to the exponential growth in food consumption caused by the predicted doubling of the world’s population to around 10 billion by the year 2050. To keep up with the increasing demand, it is essential to increase yield (Wani et al., 2016).
The most effective strategy to increase crop yields is not to increase agricultural areas, but rather to improve crop production and management techniques (Islam & Mohammad, 2022; Subedi et al., 2023). The strategies include improving crop tolerance to both very low and severe abiotic stressors (Maqsood et al., 2023), addressing biotic restrictions such as pests and disease pressures, and managing flower and fruit drops caused by physiological and hormonal abnormalities in plants could eventually lead to significant reductions in chili output.
One potential approach to reducing pest load is to manipulate plant design by agro technological interventions, including plant pruning and adjusting crop density and spacing. This method could improve chili pepper development, canopy size, and harvest (Setiawati et al., 2022). Improving chili pepper production could be easily achieved by adjusting crop density. Resource availability, climatic conditions, and chili pepper yield potential are significantly affected by plant density. Plant density could enhance yield per unit area up to a certain point, while additional densities can result in a decrease in production (Haque & Sakimin, 2022; Li et al., 2019; Shi et al., 2016). The normal chili population density is between 20,000 and 40,000 plants ha-1 (Setiawati et al., 2021). However, under ideal growth circumstances and with compact cultivars, densities much higher than this are possible. However, 30,000 plants ha-1 were the sweet spot for plant density, which was associated with lower yields.
Pruning eliminates the principal cause of apical dominance and stimulates lateral branching, which in turn creates a bushier plant structure (Walsen et al., 2023) and leads to an increase in the number of fruiting sites, increasing fruit output by 19.87 to 30.92% and decreasing insect populations by 13.33 to 88.01% (Setiawati et al., 2021). Several physiological processes may be affected by this alteration to the plant’s architecture. Disease development and pest incidences are reduced with pruning because it changes light penetration and air circulation inside the canopy (Mussa & Shinichi, 2019; Parameshwar et al., 2024). Trimming leaves helps aphid control by attracting predatory insects without affecting crop productivity (Brenard et al., 2019). However, plants’ photosynthetic capacity is reduced and fruit development is delayed when pruning is done too heavily, which in turn reduces plant growth and production (Mardhiana et al., 2017). Thus, it is important to adjust pruning techniques based on the cultivar, the weather, and the production goals to achieve a balancing condition of vegetative development and fruit production.
A wide variety of hormones found in plants, known as plant growth regulators (PGRs), are responsible for precisely controlling how plants grow and develop and eventually improve agricultural output and quality. PGRs, which may be either synthetic or naturally occurring, have an impact on plant growth and development at very low concentrations (Verma et al., 2024). These substances act as chemical messengers, starting a web of signaling pathways that control a wide range of physiological events, such as cell proliferation and extension, tissue differentiation, blossoming, fruit set, ripening, and harvest quality (Jan et al., 2020; Kaur et al., 2021). PGRs may lessen the need for pesticides since they encourage plants to become stronger and more robust, which in turn makes them less vulnerable to pest infestations (Jan et al., 2020).
To what extent the application of PGRs and plant architectural modifications could improve pest and disease control and chili pepper production is not clearly understood. Thus, this study was carried out to investigate the complementary effects of foliar sprays of PGRs with changes to plant architecture on chili pepper production. By combining these agronomic approaches, the study offers a strategy for increasing production and effective pest control and enhancing the quality of chili pepper output.
. Materials and methods
Research data were gathered for the study from July 2022 to February 2023. The experiment was conducted in West Bandung, West Java, Indonesia (6°30'S107°30'E, altitude of 1,250 meters above sea level), which has the Andisol type of soils and soil pH values between 5.0 and 6.0. The calculated average temperatures and precipitation in this area during the study were 21.3 °C and 9.7 mm month-1 (NASA Power, 2025). In the site, around 36 plots measuring 13 m × 14 m each were arranged in an 88 m × 136 m experiment area, with a one-meter gap between blocks and a half-meter gap between plots. The planting distance for all treatments was maintained at 70 × 50 cm. However, for the treatment with a plant density of 30,000 plants ha-1, a specific arrangement was applied: planting holes were positioned in a zigzag pattern. In this configuration, two seedlings were placed in the first hole of each row, followed by a single seedling in the subsequent hole, and this sequence was repeated throughout the row. 30-t ha-1 of mature compost was applied as a basal fertilizer in addition to 1-t ha-1 of NPK composite fertilizers. Silver plastic mulch was applied in each bed after the fertilizer application.
A split-split-plot experiment with a randomized block design and three replications was carried out from July 2022 to February 2023 in West Java, Indonesia. The main plot contained two crop densities (20,000 plants ha-1 and 30,000 plants ha-1), the sub-plot contained two pruning applications (without pruning and with pruning), and the sub sub-plot contained five PGR applications (NAA (50 ppm), 2,4-D (5 ppm), Triacontanol (5 ppm), Ethephon (500 ppm), Tonic (1000 ppm)) and no PGR application. The seedlings of the ‘Kencana’ chili pepper cultivar were prepared in a well-protected nursery to avoid pests and disease attacks. Three-week-old seedlings were pruned just below the first shoot of leaves using sterile shears. The seedlings were transplanted to the field two weeks after pruning to follow the experimental design. Twice throughout the experiment, at 30 and 60 days after planting (DAP), PGRs were topically sprayed using a hand sprayer in the morning.
Agronomic parameters were observed every week starting from 30 DAP on randomly selected 10 crop samples in each plot. The plant height and canopy width were monitored weekly along with other growth-related variables. The canopy width was determined by measuring the longest and shortest canopy spans from one edge to the opposite edge. The two values were then summed and divided by two to obtain the mean canopy width. A portable chlorophyll meter, the SPAD-502 type, was used to measure the chlorophyll concentration.
Crop production was recorded throughout the growing season. At each harvesting time, the fruit number per sample plant was counted, the weight of the fruit per sample plant for each harvesting time was recorded, and the average yield per hectare was determined after the final harvesting time. Additionally, fruit morphological characteristics were documented. Chili fruits in each treatment plot were collected independently to collect phenotypic data, including fruit weight (g), length (cm), diameter (cm), pericarp thickness (cm), and seed weight. Seed weight was determined by taking all the seeds from the fruit container and weighing them.
Pest and disease attacks were observed every week starting from 30 DAP on randomly selected 10 crop samples in each plot. The severity of pest infestation-related plant damage was determined using Equation 1 (Moekasan & Prabaningrum, 2017).
The variables P, v, n, Z, and N denote the percentage of damage, the highest value of the category (n = 9), the number of plants seen, and the number of plants with the same v value. The variable v is defined as an ordinal scale representing the proportion of leaf area damaged, with scores assigned as follows: 0 (no damage), 1 (>0–20% damage), 3 (>20–40% damage), 5 (>40–60% damage), 7 (>60–80% damage), and 9 (>80–100% damage). Additionally, the incidence of fruit bored due to damage was calculated using Equation 2 (Moekasan & Prabaningrum, 2017).
The percentage of weight loss was determined to assess the treatment efficacy after two weeks of storage. Two kg per treatment of uniformly healthy chili peppers without detectable pests were put on plastic trays and placed in room temperature with a relative humidity (RH) of 80% to 90%. The following equation 3 was used to determine the percentage of weight loss (Amiri et al., 2021).
A two-way analysis of variance (ANOVA) was performed to evaluate the variations across all parameters. The post-hoc test called Tukey’s Honestly Significant Difference (HSD) was used to compare the means and examine variances between the treatments at a significance level of 5%. The arcsine-square root technique of transformation was applied prior to analyzing the percentage data.
. Results
The study shows that plant responses varied depending on population density, pruning, and PGR applications, with PGRs having the most significant impact. Applying PGRs notably enhanced various aspects related to growth, yield, and plant health, emphasizing their crucial role in affecting physiological activity and reproductive performance. Population density and pruning mainly influenced structural traits such as plant height, canopy width, and fruit shape, highlighting the importance of spacing and canopy management for maximizing light capture and overall plant vigor. Key yield factors like fruit size, weight, and total yield were affected by both individual elements and their interactions, demonstrating the complex nature of crop performance. Additionally, several pest variables responded to PGRs and population density, indicating improved pest tolerance or alterations in the microenvironment. Conversely, traits like leaf number, chlorophyll levels, and pericarp thickness remained significantly different among treatments. Combining PGRs with appropriate population density and pruning practices offers a promising strategy to enhance plant growth, yield, and resilience, supporting more sustainable crop management systems (Table 1).
Table 1
Main and interaction effects of population density, pruning, and plant growth regulators (PGRs) on the measured variables.
Chili growth response to population density, pruning, and PGR treatments
Population density exerted a considerable influence on chili vegetative performance. Plants cultivated at 20,000 plants ha-1 exhibited better plant height and canopy width than those at 30,000 plants ha-1, while branch number remained statistically identical. Pruning also altered height and canopy width, with pruned plants demonstrating superior growth at lower density. However, the number of branches was not considerably changed by pruning.
The plant growth regulators (PGRs) generated marked differences across all traits. Triacontanol (5 ppm) consistently produced the highest plant height, canopy width, and branch number, whereas Ethephon (500 ppm) resulted in the lowest values. The interaction between population density and PGRs was significant, with Triacontanol maintaining superior growth performance under both densities (Table 2).
Table 2
Main and Interaction Effects of Population Density, Pruning, and Plant Growth Regulators (PGRs) on Chili Growth Characteristics.
Pest infestations as affected by population density, pruning, and PGRs
Insect pests such as chili thrips, mites, fruit borers, and oriental fruit flies were the most common pests found during the experiment. Thrips and mites are major sucking insect pests that inflict severe leaf curl on chili plants. Pest infestation levels were greatly impacted by population density. T. parvispinus (11.27%) and P. latus (10.16%) caused more damage to plants at 30,000 plants ha-1 than at 20,000 plants ha-1 (9.32% and 7.78%, respectively). However, density did not significantly affect fruit damage by Bactrocera spp. or H. armigera. Pruning considerably reduced infestation across most pests. Pruned plants at both densities experienced decreased damage by T. parvispinus (8.21–9.07%) and P. latus (6.67–7.22%) compared with non-pruned plants, which exhibited much greater levels (up to 13.46% and 13.09%). Bactrocera spp. fruit damage was also reduced in trimmed plants (4.07–5.48%), although H. armigera damage was statistically constant across pruning treatments. The PGR treatment produced substantial variation in pest incidence. Triacontanol (5 ppm) resulted in the lowest infestation from all pests, including T. parvispinus (7.50%), P. latus (6.67%), Bactrocera spp. (2.47%), and H. armigera (3.52%) (Table 3).
Table 3
Effect of population density, pruning, PGRs, and their interaction on pest infestation.
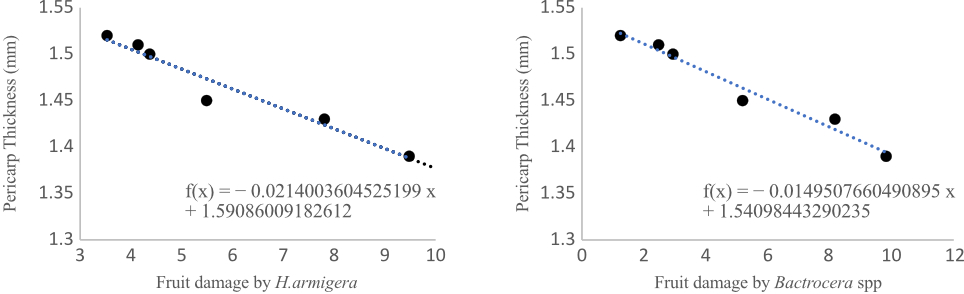
In contrast, the absence of PGRs resulted in the high pest damage, mainly from Bactrocera spp. (9.81%) and H. armigera (9.48%). The interaction between population density and PGRs was significant across all pest groups. Triacontanol consistently reduced infestation under both densities, whereas plants without PGRs experienced severe pest damage, especially at high density (up to 11.95% T. parvispinus and 10.94% P. latus; 9.18% Bactrocera spp.; 8.61% H. armigera). The beneficial effects of Triacontanol and 2,4-D were more pronounced at 30,000 plants/ha, suggesting density-dependent enhancement of PGR efficacy. Pericarp thickness and fruit damage were also shown to be negatively correlated (r2 = 0.6616 and r2 = 0.7471, respectively). Pericarp thickness makes oviposition (egg laying) and larval entry more difficult. This suggests that increasing pericarp thickness can effectively reduce fruit borer infestation and improve fruit resistance (Figure 1).
Effects of population density, pruning, and PGRs on crop yield and fruit characteristics
Population density, pruning, and plant growth regulators (PGRs) significantly affected chili yield and related traits (Table 4). Planting 20,000 plants per hectare with pruning consistently led to higher fruit numbers and weights compared to unpruned setups, indicating improved assimilate distribution with wider spacing. Among PGRs, Triacontanol (5 ppm) produced the highest fruit number (115.57 fruits plant-1), fruit weight (7.47 g), and seed yield (71.92 g), while Ethephon (500 ppm) significantly reduced all yield parameters. Yield per hectare showed a similar trend, with Triacontanol-treated plants at 20,000 plants ha-1 (pruned) achieving the highest yield (18.6 tons ha-1), followed by NAA (50 ppm). Conversely, 2,4-D and Ethephon resulted in the lowest yields.
Table 4
Effect of population density, pruning, PGRs, and their interaction on yield and yield characteristics.
The interaction between plant architecture and PGRs revealed that pruning combined with Triacontanol or NAA significantly enhanced fruit number and yield. Conversely, high-density unpruned plants treated with Ethephon exhibited the lowest performance across all yield parameters. These results demonstrate the importance of synchronizing canopy structure with hormonal treatments to optimize productivity. Pruned chili grown at 20,000 plants ha-1 also produced the highest flower count (45.56 flowers plant-1), while unpruned crops at the same density yielded the lowest (40.50). This suggests that high-density planting without pruning affects competition for resources, thereby suppressing flowering capacity. Overall, the pruned 20,000 plants ha-1 configuration yielded the highest production (15.61 t ha-1), whereas 30,000 plants ha-1 without pruning resulted in the lowest yield (14.76 t ha-1).
PGR treatments significantly influenced fruit size (length, diameter, weight), fruit quantity, and flower number. Triacontanol (5 ppm) outperformed all other treatments, producing the highest fruit count (115.6 fruits plant-1) and the heaviest fruits (16.60 g), confirming its effectiveness in promoting fruit set and development (Baghel & Singh, 2024). NAA (50 ppm) also performed well, with 103.4 fruits plant-1 and an average fruit weight of 15.8 g, indicating auxin-mediated enhancement of fruit growth. In contrast, Ethephon (500 ppm) greatly reduced flower and fruit numbers (28.00 and 34.22, respectively), likely due to its ethylene-releasing properties that cause early fruit drop and senescence (Iqbal et al., 2017). These findings highlight the importance of selecting suitable PGRs to improve reproductive success and yield stability.
The interaction among population density, pruning, and PGRs significantly influenced fruit diameter, length, and yield. Fruit diameter ranged 0.66–1.06 cm, with the highest value under 30,000 plants ha-1 + no pruning + Tonic 1000 ppm. Fruit length varied widely (7.99–17.13 cm), and the longest fruit occurred in 30,000 plants ha-1 + no pruning + Triacontanol 5 ppm. Yield ranged 10.13–18.60 t ha-1, with the highest yield produced by pruned plants + 20,000 plants ha-1 + Triacontanol 5 ppm. Ethephon consistently resulted in the smallest fruits and lowest yields across densities (Table 5).
Table 5
Three‑way interaction of population density, pruning, and PGRs on fruit morphological traits and yield on chili pepper.
Postharvest fruit weight loss was primarily driven by surface water evaporation through transpiration and respiration. Fruits from control plots experienced the highest weight loss (54.27%). In comparison, those treated with Triacontanol (18.71%) and NAA (17.88%) showed significantly lower losses (Table 4). This suggests that PGR application enhances fruit integrity and reduces deterioration during storage.
Overall, PGRs reduced postharvest weight loss by 12.03% to 50.39% compared to the control, indicating their role in preserving fruit quality during the 14-day storage period. These results reinforce the value of PGRs not only for yield enhancement but also for maintaining postharvest freshness and marketability.
. Discussion
Chili growth response to population density, pruning, and PGR treatments
The results show that lower planting density combined with pruning enhances vegetative growth by increasing light interception, air circulation, and reducing competition among plants. Pruning encourages a balanced distribution of nutrients between shoots and leaves, promoting both height and canopy expansion. These results align with Camacho-Rodríguez et al. (2022), who reported that optimized canopy management improved photosynthetic activity and uniformity in chili crops.
The superior performance under Triacontanol (5 ppm) indicates its strong effect in enhancing photosynthetic efficiency and chlorophyll biosynthesis. Triacontanol is known to activate enzymes associated with carbon fixation and promote cell division and elongation, leading to improved vegetative vigor (El-Mageed et al., 2022). The consistent positive influence of PGRs supports the view that exogenous regulators can effectively manipulate endogenous hormone balance, thereby improving growth and biomass accumulation (Backer et al., 2018).
The significant interaction between population density and PGRs indicates that hormonal responses are influenced by resource availability. The consistently positive effect of Triacontanol across different densities emphasizes its robustness, while the differing responses of the other PGRs point to density-dependent hormonal sensitivity. Together, these findings support that optimal vegetative growth results from combining lower population density with Triacontanol application.
Correlation analysis showed that yield was positively related to plant height (r = 0.635), canopy width (r = 0.681), number of branches (r = 0.714), number of leaves (r = 0.592), and chlorophyll concentration (r = 0.746). Among these, chlorophyll contents and the number of branches had the strongest connections with yield, highlighting their important role in productivity. The strong links between yield and vegetative traits suggest that vigorous canopy growth and photosynthetic activity help promote assimilate buildup for fruit development. Elevated chlorophyll concentration is a reliable indicator of improved photosynthetic efficiency, which directly boosts assimilate production and yield potential (Croce et al., 2024). These results indicate that optimizing plant architecture and physiological traits can effectively improve yield performance in chili production systems (Table 6).
Table 6
Correlation coefficients among growth, chlorophyll content, and chili yield.
Overall, the integration of pruning at 20,000 plants ha-1 with Triacontanol application provided the most favorable combination for vigorous vegetative development, optimizing canopy structure and photosynthetic potential of chili plants.
Pest infestations as affected by population density, pruning, and PGRs
Previous studies demonstrated the impact of PGRs on pest populations through their effect on pests’ reproduction, development, fecundity, and egg viability (Er & Keskin, 2016; Zhao et al., 2017). PGRs impede embryonic development, alter gametogenesis, or function as growth inhibitors. Another study found that crops treated with 2,4-D and Triacontanol developed systemic acquired resistance to herbivores (Gordy et al., 2015). The importance of pericarp thickness in providing resistance against pests such as fruit borers and fruit flies has been highlighted in previous studies (Gong et al., 2024; Mushtaq & Pathania, 2020). A thicker pericarp acts as a physical barrier, making it harder for pests to penetrate and harm the fruits. Tomatoes with a high pericarp thickness of 0.62 cm exhibited a 6.75 % reduction in fruit weight loss (Bisht et al., 2020), suggesting that the pericarp thickness is crucial for tomato resistance to H. armigera. Pericarp thickness was sensitive to the plant architectures and PGRs in chili peppers. Pericarp thickness and fruit weight changed as a consequence of the application of PGRs and certain plant architecture. Thus, by incorporating these measures, pest control could be improved, and chili pepper harvests can be boosted.
Correlation analysis revealed strong positive associations among the major pest species, particularly between H. armigera and Bactrocera spp. (r = 0.944**) and between Thrips parvispinus and Bactrocera spp. (r = 0.890**). In contrast, yield exhibited negative correlations with all pest infestations, most notably with P. latus (r = –0.732**) and H. armigera (r = –0.301*), indicating that increased pest pressure was associated with reduced productivity. The significant positive correlations among T. parvispinus, H. armigera, and Bactrocera spp. suggest potential overlapping infestation dynamics, where simultaneous pest occurrence amplifies crop damage. Similar patterns of pest interaction have been reported in solanaceous crops, where thrips and fruit borers often coexist and synergistically reduce fruit set and marketable yield (Hossain et al., 2021).
The negative correlations between pest populations and yield confirm that higher pest density leads to significant yield loss, as previously observed in chili and tomato systems (Jangra et al., 2017). Among pests, P. latus showed the strongest negative effect (r = –0.732**), consistent with reports that P. latus infestations impair photosynthesis and fruit development (Patavardhan et al., 2020). These results demonstrate that optimal pest suppression is achieved through a combination of lower population density, pruning, and Triacontanol application, offering an integrated approach to mitigating key chili pests. The data indicate that integrated pest management strategies targeting thrips, mites, and fruit borers simultaneously are essential to sustain high productivity in chili production (Table 7).
Effects of population density, pruning, and PGRs on crop yield and fruit characteristics
The findings reveal that plant growth regulators, pruning, and population density are key components of plant architectural management techniques that significantly influence chili pepper productivity and quality. These elements work synergistically to promote balanced development in chili fruits, enhance fruit quality, and improve postharvest performance. Consistent with these findings, Zhumanova et al. (2024) observed notable improvements in growth, fruit quality, and resilience to biotic stresses when PGRs were combined with plant architectural management. The interplay between plant design and PGRs enhances chili fruit performance. To optimize PGR use across different fruit crop systems and promote sustainable farming practices, further research is needed into the specific relationships between various PGRs and plant architecture strategies. PGRs and plant structure influence the size and weight of chili pepper seeds. A well-balanced plant with optimal height, branching, and leaf area promotes better light penetration and air circulation, leading to improved flowering and fruit set. Triacontanol, for example, boosts photosynthesis and carbon assimilation, resulting in increased fruit set and yield (Alharbi et al., 2021). Likewise, pruning or canopy-training that enhances light exposure and reduces intra-canopy competition improves fruit set efficiency (Anthony & Minas, 2021). Therefore, combining these PGRs with suitable plant architecture is likely to affect fruit yield, fruit weight, and seed production. PGRs improve key aspects of plant architecture critical for seed development, such as height, biomass, leaf area, and root growth, thereby potentially increasing seed yield and weight. Moreover, PGRs may enhance seed production and, in some cases, seed weight (Fatima et al., 2023). Additionally, the results indicate that PGRs significantly improve the postharvest quality of chili fruits, especially by reducing weight loss. They do so by enhancing enzyme activities, delaying nutritional decline, and maintaining fruit firmness and volatile compounds (Hayati et al., 2023). Optimized plant architecture supports better fruit development and resistance to diseases, which extends shelf-life and improves storage outcomes. While PGRs contribute to better postharvest qualities like firmness and nutrient retention, both plant structure and PGR application are most effective when implemented before harvest (Fatima et al., 2023).
The interaction of population density, pruning, and PGRs clearly shaped fruit morphology and yield in red chili. Triacontanol and NAA consistently enhanced fruit length, diameter, and yield, confirming their role in stimulating photosynthesis and reproductive efficiency. In contrast, Ethephon suppressed fruit development, reflecting ethylene’s inhibitory effect on growth and yield.
Lower plant density favored reproductive output, while higher density improved fruit size, highlighting a trade-off between number and weight. Pruning produced high flower initiation but low fruit filling, whereas unpruned plants produced fewer but heavier fruits (Table 4). The best outcomes were achieved when Triacontanol was combined with optimal density and pruning, yielding longer fruits and higher productivity with reduced postharvest losses.
Overall, integrating effective PGRs with proper spacing and canopy management provides a practical strategy to maximize chili yield and fruit quality under tropical production systems.
Correlation analysis (Table 8) revealed highly significant associations between flower number and fruit number (r = 0.933), fruit weight (r = 0.900), and yield (r = 0.954), indicating that floral productivity directly influences final yield potential. Fruit weight was strongly correlated with seed number (r = 0.932) and yield (r = 0.944). This emphasizes that yield largely depends on the extent of fruit mass accumulation. In contrast, fruit weight loss and morphological traits such as length or diameter showed nonsignificant relationships with yield, suggesting they have a secondary influence in field conditions. These results align with previous studies showing that Triacontanol and NAA enhance fruit set, seed yield, and overall productivity in chili and other Solanaceae (Sahu et al., 2017; Kumar & Pandey, 2018). Therefore, combining moderate plant density (20,000 plants ha-1), pruning, and Triacontanol seems to optimize canopy balance, leading to the highest chili yield and quality (Table 5). Overall, integrating PGRs with suitable population density and pruning practices can significantly improve chili growth, yield, and resilience. Such strategies optimize physiological activity, improve postharvest quality, and support sustainable chili production in tropical agroecosystems.
Table 8
Correlation coefficients between yield and yield‑attributing components of chili.
In conclusion, integrated crop management through optimal plant density of 20,000 plants ha-1, pruning, and Triacontanol application enhances chili pepper yield, reduces pest damage, and improves post-harvest shelf-life.
