. Wstęp
Współczesne lasy Polski są mozaiką zbiorowisk o różnym stopniu zachowania. Ocena zakresu ich przekształceń jest jednym z najważniejszych zadań w ochronie przyrody. Wokół naturalności lasu, począwszy od układu pierwotnego po antropogeniczny, było wiele dyskusji (np. Faliński, 1986, 1993; Dzwonko, 2017; Pawlaczyk, 1996; Peterken, 1996, Świerkosz, 2007). Za najważniejsze wydaje się zdefiniowanie pojęć lasu puszczańskiego (lasu pierwotnego; primary forest) oraz starego lasu (old-growth forest). Terminy te różnie były definiowane w literaturze (por. Cieśliński et al., 1996; Czyżewska & Cieśliński, 2003; Dzwonko, 2017). Ważnym głosem porządkującym zakres ich ujęcia był dokument Komisji Europejskiej z 2023 roku pt. Commission Guidelines for Defining, Mapping, Monitoring and Strictly Protecting EU Primary and Old-Growth Forests. Jako „las puszczański” rozumie się tutaj układ z rodzimymi gatunkami drzew, w którym nie ma wyraźnych śladów działalności człowieka, a procesy ekologiczne nie są znacząco zakłócone (w przeszłości nie było istotnej ingerencji człowieka lub ostatnia była na tyle dawno, że naturalny skład gatunkowy i procesy zdążyły się odtworzyć). W definicji starego lasu podkreśla się już widoczny aktualnie ślad działalności człowieka, który jednak wyraźnie zanika i nie wpływa na aktualne procesy ekologiczne. Obie kategorie obejmują lasy o najwyższym stopniu naturalności zajmujące stosunkowo duży obszar gwarantujący niezakłócony przebieg naturalnych procesów dynamiki lasu. Ważne z punktu widzenia występowania mszaków (i innych organizmów zarodnikowych) w lasach puszczańskich (i starych lasach) jest bogactwo i dostępność siedlisk dla epifitów i epiksyli (Rycina 1) tj. kora wielu gatunków różnowiekowych drzew (w tym bardzo starych i obumierających) oraz martwego drewna o różnym stopniu rozkładu – leżących kłód, złomów i pniaków (Czyżewska & Cieśliński, 2003). Do identyfikacji najlepiej zachowanych zbiorowisk wykorzystuje się gatunki wskaźnikowe określane jako relikty puszczańskie (relikty starych lasów). W polskiej geobotanice pojęcie to wprowadzili badacze Puszczy Białowieskiej i używali je początkowo w odniesieniu do roślin naczyniowych i porostów (np. Cieśliński & Tobolewski, 1988; Faliński, 1977). Pierwsza lista mchów i wątrobowców jako reliktów puszczańskich powstała dopiero z końcem lat 90. XX wieku w wyniku realizacji projektu badawczego pt. „Rola roślin zarodnikowych w zbiorowiskach leśnych Białowieskiego Parku Narodowego”, w skrócie CRYPTO (Cieśliński et al., 1996).
Rycina 1
Krajobrazy wnętrza lasu puszczańskiego w polskiej części Tatr Wysokich: 1–3 – Dolina Roztoki, 4 – Dolina Łężna; 1 – kłody porośnięte głównie przez Fuscocephaloziopsis catenulata, 2 – kłoda z trzema gatunkami reliktowymi: Crossocalyx hellerianus, Anastrophyllum michauxii i Fuscocephaloziopsis catenulata; 3 – nasadowa część pnia świerka z obficie występującym Lophoziopsis longidens, 4 – kłoda zawieszona w poprzek wąwozu wapiennego z bogatą populacją Fuscocephaloziopsis catenulata i Harpanthus scutatus (fot. P. Górski).

Wykaz wątrobowców wskaźnikowych dla starych lasów, w oparciu o zdefiniowane kryteria, przygotował Profesor Henryk Klama (Cieśliński et al., 1996; Klama, 2002a, 2002b). Wyjściową pulą gatunków do wyboru była hepatikoflora Białowieży. Jest ona stosunkowo bogata i liczy 67 taksonów (Klama, 2002a, 2017). Liczebnie przewyższa więc większość, za wyjątkiem Wigierskiego Parku Narodowego (79, Górski P. & Romański M., mat. npbl.), niżowych obiektów tego typu. Przez lata lista ta była pomocna w ocenie stopnia naturalności lasów niżowych. Jednak jeśli weźmiemy pod uwagę obszary górskie tj. Tatry, to pula gatunków tu występujących (w stosunku do białowieskiej puszczy) zwiększa się trzykrotnie (Górski & Váňa, 2014; Górski et al., 2023). Wydaje się oczywiste, że zestaw gatunków wskaźnikowych powinien ulec poszerzeniu, ale także status niektórych niżowych reliktów występujących jednocześnie w górach, może ulec zmianie. Dla mchów różnice w zestawie gatunków puszczańskich na niżu a w górach są bardzo duże. Lista „białowieska” zawiera ich 13 (J. Żarnowiec w Cieśliński et al., 1996), w Bieszczadach jest już ich 24 (Stebel & Żarnowiec, 2014). W Białowieży gatunków wskaźnikowych wśród porostów jest 43 (Cieśliński et al., 1996), ale już „niżowa” lista (a więc nadal bez gatunków górskich lasów) zawiera ich 71 (Czyżewska & Cieśliński, 2003). Dla grupy wątrobowców propozycji nowych regionalnych list nie było, za wyjątkiem niewielkiego obszaru jakim jest Ojcowski Park Narodowy (Stebel & Vončina, 2020).
Celem niniejszej publikacji jest podanie listy wątrobowców wskaźnikowych lasów puszczańskich (i starych lasów) Tatr wraz z charakterystyką ich rozmieszczenia i zasobów w Tatrach i w Polsce. Wykaz przygotowano na podstawie wieloletnich badań prowadzonych w tym masywie przez Autora. Gatunki reliktowe wytypowano z uwzględnieniem zdefiniowanych wcześniej kryteriów (por. dalej, rozdział 2) oraz – w niewielkim stopniu – subiektywnej oceny eksperckiej. Nazewnictwo łacińskie wątrobowców przyjęto według Klamy i Górskiego (2018), polskie – Szweykowskiego (2006). Pominięto fakt braku nowych polskich nazw dla taksonów powstałych w wyniku zmian nomenklatorycznych (tu dotyczy rodzajów Fuscocephaloziopsis i Lophoziopsis) i podano nazwę polską z wykazu Szweykowskiego (2006).
. Kryteria wyboru gatunków wskaźnikowych na niżu a w górach
W badaniach prowadzonych w Puszczy Białowieskiej przyjęto następujące kryteria opisujące gatunki wskaźnikowe lasów puszczańskich (Cieśliński et al., 1996; Klama, 2002b): 1) rosną tylko w zbiorowiskach leśnych (typowe leśne gatunki), 2) wykazują pełną żywotność na danym obszarze (tj. przechodzą pełny cykl życiowy, tworzą propagule, mają typowe dla gatunku rozmiary i barwę), 3) wymarły lub wymierają, albo są bardzo rzadkie na niżu Polski a rosną nadal w Puszczy Białowieskiej, gdzie mają liczne populacje, 4) zasiedlają mikrosiedliska charakterystyczne dla lasów puszczańskich (np. sędziwe drzewa, drewno kłód w różnym stadium rozkładu itp.), 5) nie wykazują tendencji do wkraczania na siedliska antropogeniczne. Lista białowieska liczy 7 gatunków wątrobowców. Są to: Anastrophyllum michauxii (F. Weber) H. Buch, Barbilophozia lycopodioides (Wallr.) Loeske, Bazzania trilobata (L.) Gray, Fuscocephaloziopsis catenulata (Huebener) Váňa & L. Söderstr., Lophoziopsis longidens (Lindb.) Konstant. & Vilnet, Plagiochila asplenioides (L.) Dumort. oraz Crossocalyx hellerianus (Nees ex Lindenb.) Meyl. (H. Klama w Cieśliński et al., 1996).
Wymienione powyżej kryteria, w odniesieniu do gatunków występujących w górach, należy dodatkowo skomentować. Obszary górskie z natury cechują się występowaniem skał w pasie reglowym. To powoduje, że niektóre wątrobowce związane z mikrosiedliskami leśnymi (np. epifity) mogą rosnąć dodatkowo na skałach (głazy i ściany skalne w obrębie lasu), a także na powierzchniach skalnych w piętrach wysokogórskich. Mamy tu więc sytuację, że gatunek reliktowy związany jest zarówno z formacją leśną jak i nieleśną, co jest w sprzeczności z kryterium 1 (por. powyżej). Przykładem jest uznany już wcześniej za reliktowy Lophoziopsis longidens, bardzo rzadki w lasach niżowych północno-wschodniej Polski (Klama, 1997, 2002a, 2002b), a w Tatrach występujący jako epifit (głównie świerka) i epilit (w borach) aż po piętro alpejskie. Wątrobowiec ten posiada 25 stanowisk naskalnych powyżej 1700 m n.p.m. (P. Górski, npbl.), a jego maksimum wysokościowe wynosi 2254 m n.p.m. (Górski et al., 2023). Identycznie będą występowały zaproponowane w niniejszej pracy nowe relikty tj. Frullania fragilifolia oraz F. jackii. Kolejne kryterium (nr 2) jest trudne w ocenie, w praktyce briologicznej należy je odrzucić. Nie ma badań pokazujących, że stopień naturalności lasu ma odzwierciedlenie w fakcie tworzenia sporofitów czy rozmnóżek. Dla wątrobowców (lub szerzej mszaków) raczej należy to wiązać z cechami biologicznymi gatunku. Wspominają o tym także Stebel & Żarnowiec (2014). Kryterium 3 odnosi się do rzadkości gatunków wskaźnikowych na danym obszarze. Uwzględniając je, w Tatrach jako reliktowe nie można uznać Bazzania trilobata, Barbilophozia lycopodioides i Plagiochila asplenioides, które są pospolitymi gatunkami związanymi także z lasami niewykazującymi cech puszczańskich. Wymienione gatunki posiadają odpowiednio 126, 459 oraz 308 stanowisk w całych Tatrach (Górski & Váňa, 2014; P. Górski npbl.). W ramach omawianego kryterium należy zwrócić uwagę raczej na częstość występowania gatunku reliktowego i ten fakt eksponować jako parametr waloryzacyjny. Za las bardziej naturalny uznalibyśmy występowanie gatunku wskaźnikowego np. na wielu kłodach lub drzewach (nawet położonych blisko obok siebie) niż jego jednostkowe wystąpienie. W Tatrach, gdzie stopień naturalności lasów jest dużo wyższy niż na niżu, odnalezienie jednego stanowiska gatunku wskaźnikowego może być wyrazem nie tyle „puszczańskości” kompleksu leśnego, ile korzystnych warunków mikroklimatycznych w danym miejscu. Przyjęcie tego nowego kryterium generuje bardziej pracochłonny zbiór danych terenowych, ale daje dobry obraz tego jak „czują się” w danym kompleksie leśnym najrzadsze gatunki lasu pierwotnego. Pozostałe kryteria nie wymagają komentarza i mają takie samo zastosowanie dla gatunków niżowych i górskich.
. Wykaz reliktów
Dotychczasowa lista gatunków wątrobowców lasów puszczańskich (Cieśliński et al., 1996; Klama, 2002a, 2002b) wymaga korekty w odniesieniu do Tatr. Trzy gatunki (Barbilophozia lycopodioides, Bazzania trilobata oraz Plagiochila asplenioides) jako pospolite należy odrzucić. Pozostałe wątrobowce tj. Anastrophyllum michauxii, Fuscocephaloziopsis catenulata, Lophoziopsis longidens oraz Crossocalyx hellerianus są nadal dobrymi wskaźnikami lasów puszczańskich. Wykaz ten należy uzupełnić o kolejne rośliny tj. Harpanthus scutatus (F. Weber & D. Mohr) Spruce, Frullania fragilifolia (Taylor) Taylor ex Gottsche, Lindenb. & Nees, F. jackii Gottsche, Metzgeria violacea (Ach. ex F. Weber & D. Mohr) Dumort., Porella arboris-vitae (With.) Grolle, Scapania apiculata Spruce oraz S. carinthiaca J.B. Jack ex Lindb. Stopień zagrożenia i zasoby wymienionych gatunków reliktowych przedstawiono w tabeli 1. Łącznie wykaz obejmuje 11 wątrobowców. Wszystkie są zagrożone w skali Polski, w tym 5 z najwyższym stopniem zagrożenia (CR). Liczby epiksyli (6) i epifitów (5) wśród reliktów są zbliżone. Wszystkie epifity, za wyjątkiem Metzgeria violacea, mają równocześnie swoje populacje naskalne w pasie reglowym lub wysokogórskim. Jako wskaźnikowe należy uznać jedynie wystąpienia tych gatunków na korze żywych drzew (stanowiska epifityczne).
Tabela 1
Wykaz gatunków wątrobowców reliktów lasów puszczańskich Tatr, ich siedliska, zagrożenie oraz aktualne zasoby
| Gatunek | 1 Siedl. | 2 Zagr. PL | 3 Zagr. Tatry PL+SL | 4 Liczba stan. lit. PL | 5 Liczba stan. lit. SL | 6 |
|---|---|---|---|---|---|---|
| Anastrophyllum michauxii | K,f,l,g | VU | NT | 17 | 31 | Cieśliński et al., 1996 |
| Crossocalyx hellerianus | K,f | EN | VU | 4 | 8 | Cieśliński et al., 1996 |
| Frullania fragilifolia | F,L | CR | EN | 12 | 3 | Górski, oryg. |
| Frullania jackii | F,L | CR | EN | 3 | 7 | Górski, oryg. |
| Fuscocephaloziopsis catenulata | K | VU | NT | 5 | 18 | Cieśliński et al., 1996 |
| Harpanthus scutatus | K | EN | VU | 2 | 13 | Górski, oryg. |
| Lophoziopsis longidens | F,L,k | VU | VU | 9 | 13 | Cieśliński et al., 1996 |
| Metzgeria violacea | F | CR | CR | 2 | 1 | Górski, oryg. |
| Porella arboris-vitae | F,L | VU | CR | 3 | 0 | Górski, oryg. |
| Scapania apiculata | K | CR | CR | 2 | 4 | Górski, oryg. |
| Scapania carinthiaca | K | CR | RE | 1 | 2 | Górski, oryg. |
[i] Objaśnienia: 1 Siedl. – typ zajmowanych siedlisk: F – epifit, G – epigeit, K – epiksyl, L – epilit (pisane wielką literą – najczęstsze występowanie, pisane małą literą – rzadko obserwowane), 2 Zagr. PL – zagrożenie w Polsce, według listy Klamy & Górskiego (2018); 3 Zagr. Tatry PL+SL – zagrożenie w skali całych Tatr, według listy Górskiego (2020); 4/5 Liczba stan. lit. PL/SL – liczba znanych stanowisk z literatury opublikowanych w latach 1814–2024 w Polskich Tatrach/Słowackich Tatrach; 6 – nadanie statusu gatunku wskaźnikowego lasów puszczańskich.
. Charakterystyka gatunków wskaźnikowych lasów puszczańskich Tatr
Anastrophyllum michauxii (F. Weber) H. Buch
Liczba stanowisk publikowanych: Słowacja (SL): Tatry Bielskie TB – 31, Tatry Wysokie TW – 14, Tatry Zachodnie TZ – 7, Polska (PL): TW – 8, TZ – 9.
Zgiętolist Michauxa jest gatunkiem epiksylicznym, rzadko rośnie na korze żywych drzew oraz skałach i głazach śródleśnych, wyjątkowo na glebie. W Polsce jego stanowiska skupione są głównie w obszarach górskich, w Sudetach (Góry Stołowe i Bialskie; Koła, 1968; Szweykowski 1953a, b) oraz Karpatach (Tatry, Babia Góra, Gorce, Bieszczady; Górski & Váňa, 2014; Klama, 2004, 2013a, b; Mierzeńska, 1994; Szweykowski & Buczkowska, 1996). Bardzo rzadko notowany był na niżu, w północno-wschodniej Polsce (Puszcza Białowieska; Karczmarz & Kornijów, 1981; Karczmarz & Sokołowski, 1981; Klama, 2002a, 2002b) oraz na Dolnym Śląsku (stanowisko z XIX wieku, por. Szweykowski, 2006).
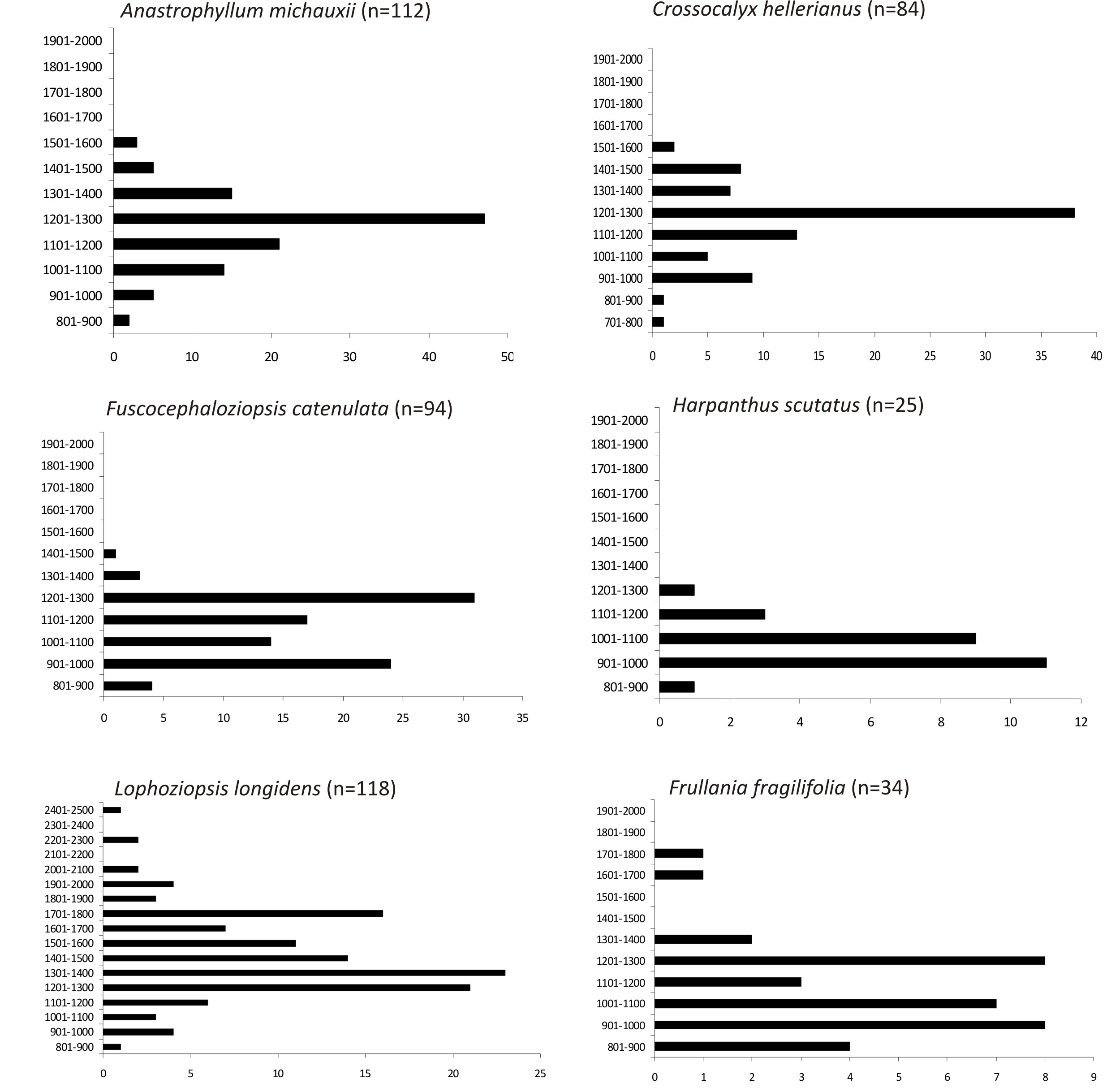
Spośród wszystkich gatunków wskaźnikowych lasów puszczańskich Anastrophyllum michauxii posiada najwięcej stanowisk publikowanych w Tatrach – 55 (Blackburn et al., 1997; Duda, 1962, Duda & Váňa, 1971, 1984; Górski & Váňa, 2014, Górski & Rusińska et al., 2016; Górski & Staniaszek-Kik et al., 2017; Górski & Vončina et al., 2016; Górski et al., 2020, 2023; Hazslinszky, 1885; Krupa, 1882, 1888; Limpricht, 1877a, 1877b; Rehman, 1864; Szweykowski, 1958, 1960; Szweykowski & Buczkowska, 2000; Szyszyłowicz, 1884; Šmarda, 1938, 1958, 1961, 1976). Maksimum wysokościowe tej rośliny przypada na 1575 m n.p.m. (Słowacja, Tatry Wysokie, Grúnik, leg. J. Šmarda, 1947; Šmarda, 1961), a minimum – 895 m n.p.m. (Słowacja, Tatry Zachodnie, Trnovská dolina, leg. P. Górski, 2015; Górski & Vončina et al., 2016). W profilu pionowym Tatr, najwięcej wystąpień Anastrophyllum michauxii (47 na 112) odnotowano w pasie wysokościowym 1200–1300 m n.p.m. (Rycina 3). W terenie dość często obserwuje się peryancja i osobniki męskie tego dwupiennego gatunku. Rzadziej roślina wytwarza pędy z rozmnóżkami. Omawiany wątrobowiec w skali kraju jest zagrożony (VU, Klama & Górski, 2018), jednak regionalnie, w odniesieniu do całego masywu tatrzańskiego, jest niezagrożony (NT; Górski, 2020).
Crossocalyx hellerianus (Nees ex Lindenb.) Meyl. [= Anastrophyllum hellerianum (Nees ex Lindenb.) R. M. Schust.]
Liczba stanowisk publikowanych: (SL): TB – 7, TW – 1, TZ– 0, (PL): TW – 2, TZ – 2.
Zgiętolist nadrzewny jest obligatoryjnym epiksylem (jeden raz obserwowano go na korze żywego świerka w Tatrach Wysokich; P. Górski, 2024, npbl.). W Polsce stanowiska tej rośliny rozproszone są na Pomorzu Zachodnim (Szweykowski & Koźlicka, 1969) i w północno-wschodniej części (Puszcze – Białowieska, Romincka, Knyszyńska, Borecka, Wigierska; Górski et al. 2014, Górski & Rusińska et al., 2016, Karczmarz & Sokołowski, 1995; Klama, 2002a, 2002b; Szweykowski, 1956; Wiśniewski & Rejment, 1935). Ponadto, wątrobowiec ten zbierany był w Górach Świętokrzyskich i Kotlinie Sandomierskiej (Karczmarz & Bloch, 1985; Stebel et al., 2013). W obszarach górskich, poza Tatrami, obserwowany był w Górach Stołowych (Szweykowski, 1953a, b), Śnieżniku Kłodzkim (Szweykowski, 1956), Pieninach (Szweykowski, 1961; Vončina et al., 2023), Gorcach (Mierzeńska, 1994), Beskidzie Żywiecko-Orawskim (Klama, 1996), Beskidzie Sądeckim (Mamczarz, 1977), Beskidzie Śląskim (Rejment-Grochowska, 1950).
W całym masywie tatrzańskim Crossocalyx hellerianus ma 12 stanowisk (Duda & Váňa, 1984; Górski & Váňa, 2014; Klama, 2008; Limpricht, 1877a, b; Szweykowski, 1956, 1960, 1971). Maksimum wysokościowe gatunku przypada na wysokości 1522 m n.p.m. (nowe stanowisko: Polska, Tatry Wysokie, Żabie, lasy na NE od Morskiego Oka, bór limbowo-świerkowy Vaccinio-Pinetum cembrae , próchniejąca kłoda, 49.209387°N, 20.085690°E, MGRS 34UDV3351, leg. P. Górski 1916/2024, 6.09.2024, POZNB 5553), minimum zaś położone jest na 800 m n.p.m. (Tatranská Lomnica, leg. A. Boros; Duda & Váňa, 1984). Zgiętolist nadrzewny posiada najwięcej notowań w Tatrach (38 na 84) w pasie wysokościowym 1200–1300 m n.p.m. (Rycina 3).
Crossocalyx hellerianus jest bardzo drobnym gatunkiem (Rycina 2), trudnym w obserwacji terenowej. Możliwość jego dostrzeżenia związana jest z odpowiednią wilgotnością wnętrza lasu i liczbą gametoforów na kłodzie. Pojedyncze wystąpienia, w suchym powietrzu praktycznie są przeoczane. Roślina najczęściej wytwarza pędy rozmnóżkowe i wtedy jest najłatwiejsza do odnalezienia. Rzadko tworzy peryancja. Wytwarza także pędy pozbawione rozmnóżek i łatwa jest do pomylenia z gatunkami epiksylicznymi z dwoma łatami liści (i przeoczana). W rzeczywistości zasoby populacyjne tego wątrobowca w Tatrach są większe niż wynika to z danych publikowanych, na co wskazują badania w polskiej części Tatr Wysokich (P. Górski, 2024, npbl.).
Rycina 2
Wybrane gatunki wątrobowców wskaźnikowych lasów puszczańskich: 1 – Crossocalyx hellerianus, 2 – Lophoziopsis longidens, 3 – Anastrophyllum michauxii, 4 – Frullania fragilifolia; 1 – Wigierski Park Narodowy (fot. M. Romański), 2-4 – Tatry Wysokie, Dolina Roztoki (fot. P. Górski).

Frullania fragilifolia (Taylor) Taylor ex Gottsche, Lindenb. & Nees
Liczba stanowisk publikowanych: (SL): TB – 2, TW – 0, TZ– 1, (PL): TW – 1, TZ – 9.
Miedzik krucholistny jest w Tatrach gatunkiem epifitycznym oraz epilitycznym. Notowany był tutaj na korze buka, jodły, jaworu i świerka (Szweykowski, 1960; P. Górski, npbl.) oraz skałach w pasie reglowym (P. Górski, npbl.) i piętrze alpejskim (leg. J. Szweykowski, 1960; Górski, 2013a). W Polsce roślina ta podawana była z Pomorza Zachodniego, przedgórza Sudeckiego, Tatr i Bieszczad (Koppe & Koppe, 1940; Krawiec, 1938; Krupa, 1878; Limpricht, 1876; Müller, 1912–1916; Szweykowski, 1958, 1960; Szyszyłowicz, 1884). Dane literaturowe i materiały zielnikowe zgromadzone w polskich herbariach (KRAM, LBL, POZW, POZNB, WA) wskazują, że Frullania fragilifolia posiada w Polsce 20 stanowisk historycznych (Górski, 2013a), z tego najwięcej notowań pochodzi z XIX wieku.
Po 2000 roku opublikowano w Tatrach 2 stanowiska Frullania fragilifolia : z doliny Kościeliskiej (po polskiej stronie; Górski, 2013a) oraz doliny Juraniowej (Słowacja, leg. P. Górski, 2013; Górski & Váňa, 2014). W profilu pionowym Tatr miedzik krucholistny zanotowany był najniżej na wysokości 880 m n.p.m. (Tatry Bielskie, Dolina Hučáva, na korze jodły, leg. J. Szweykowski, 1957; Szweykowski, 1960), a najwyżej na mylonitach zachodnich stoków Jarząbczego Wierchu – 1710 m n.p.m. (Polska, Tatry Zachodnie, leg. J. Szweykowski, 1959; Górski & Váňa, 2014). Najwięcej stanowisk tatrzańskich odnotowano w pasach wysokościowych 901–1000 i 1201–1300 m n.p.m. (Rycina 3).
W ocenie Szweykowskiego (2006) Frullania fragilifolia prawdopodobnie wyginęła w Polsce. W ostatnich latach została jednak odnaleziona na kilku stanowiskach w Tatrach Polskich i Słowackich, zarówno jako epifit jak i epilit (P. Górski, 2024, npbl.). Prawdopodobnie masyw ten może być jedynym miejscem występowania tego miedzika w Polsce. Na Pomorzu Zachodnim ostatnio widziany był 85 lat temu (Koppe & Koppe, 1940), w Bieszczadach w 1956 roku (Szweykowski & Buczkowska, 1996) a w Polsce południowo-zachodniej (Pogórze Karkonoskie) z końcem XIX wieku (Górski, 2013a).
Rycina 3
Rozmieszczenie pionowe w Tatrach w 100-metrowych pasach wysokościowych wybranych wątrobowców wskaźnikowych lasów puszczańskich [na osi X zaznaczono liczbę stanowisk, na y – zakres wysokości w m (n.p.m.); n – liczba stanowisk ustalona na podstawie danych publikowanych począwszy od 1814 roku i niepublikowanych (P. Górski, 2002–2024)].
Frullania jackii Gottsche
Liczba stanowisk publikowanych: (SL): TB – 2, TW – 3, TZ– 2, (PL): TW – 0, TZ – 3.
W Polsce Frullania jackii występuje tylko w Tatrach i ma status gatunku krytycznie zagrożonego (Klama & Górski, 2018; Szweykowski, 2006). Większość stanowisk z masywu tatrzańskiego (7 na 10) udokumentowana była po słowackiej stronie (Duda, 1955a; Duda & Váňa, 1976; Szweykowski, 1960; Šmarda 1960a, b, 1961) i te dotyczą siedlisk naskalnych. Jedyne epifityczne stanowiska F. jackii pochodzą z polskiej strony Tatr Zachodnich i były obserwowane z końcem XIX wieku („Dolina do Dziury na korze drzew, 891–1026 m”; por. Szyszyłowicz, 1884, s. 37). Te dane kwestionuje Szweykowski (2006), jednak to tylko przypuszczenie, gdyż nie zachował się materiał zielnikowy do weryfikacji. Od tego czasu nie obserwowano F. jackii jako epifita. Z tego samego okresu, w roku 1876, F. jackii zebrał Tytus Chałubiński w dolinie Strążyskiej i oznaczył jako F. dilatata. Szczęśliwie zbiór się zachował (rev. P. Górski, 2017, WA 37642, por. Górski & Graniszewska, 2017), natomiast na etykiecie brakowało szczegółowych danych lokalizacyjnych, wysokości i siedliska. Był natomiast dopisek „i ślady Lejeunea calcarea Lib.”, co sugeruje, po obecności Cololejeunea calcarea, siedlisko naskalne. Trudno obecnie przesądzić, na ile miedzik górski jest dobrym wskaźnikiem lasów o puszczańskim charakterze i czy jego brak jako epifita wiązać można ze stopniem naturalności ekosystemów leśnych Tatr, biologią gatunku czy niewystarczającym stanem badań. Zasoby genowe Frullania jackii w Tatrach nie są duże, ale stanowiska naskalne (także położone niżej) są obecne. W latach 2017–2024 roślinę tę odnotowano na skałach mylonitowych Tatr Wysokich i Zachodnich na 6 stanowiskach, w zakresie wysokościowym 1302–1795 m n.p.m. (P. Górski, npbl.). Minimum wysokościowe F. jackii przypada na wysokość 891 m n.p.m. (podane jako 891–1026 m; polskie Tatry Zachodnie, Dolina ku Dziurze; Szyszyłowicz, 1884), a maksimum na 1795 m n.p.m. (nowe stanowisko: Polska, Tatry Zachodnie, Dolina Starorobociańska, Wielkie Jamy, między Siwym Zwornikiem a Gaborową Przełęczą, mylonity, 49.202564°N, 19.827578°E, MGRS 34UDV1450, leg. P. Górski 486/2021, 14.09.2021, POZNB 4000).
Fuscocephaloziopsis catenulata (Huebener) Váňa & L. Söderstr. [= Cephalozia catenulata (Huebener) Lindb.]
Liczba stanowisk publikowanych: (SL): TB – 12, TW – 3, TZ– 3, (PL): TW – 3, TZ – 2.
Głowiak łańcuszkowaty jest obligatoryjnym epiksylem. Jego stanowiska w Polsce znane są z prawie wszystkich masywów górskich oraz północnej części (głównie NE) kraju (Szweykowski, 2006). Na południu Fuscocephaloziopsis catenulata podawany był z Gór Izerskich, Karkonoszy, Gór Sowich, Kotliny Jeleniogórskiej, Babiej Góry, Tatr (i Podtatrza), Gorców, Bieszczad, Beskidu Śląskiego, Żywiecko-Orawskiego i Sądeckiego (Górski & Váňa, 2014; Klama, 1996, 2004, 2008; Mamczarz, 1977; Mierzeńska, 1994; Szweykowski 1958, 1960; Szweykowski & Buczkowska, 1996). Niżowe stanowiska F. catenulata obejmują Pojezierza i Pobrzeża Południowobałtyckie, Pojezierza Wschodniobałtyckie, Nizinę Północnopodlaską, Wyżynę Kielecką (Góry Świętokrzyskie) oraz Roztocze (Fudali et al., 2015; Górski 2013b; Górski et al., 2014; Klama, 2002a, 2002b; Stebel et al., 2013; Szweykowski, 1958). Mapę rozmieszczenia F. catenulata w Polsce na podstawie danych literaturowych przedstawiają Górski & Romański (2016).
W Tatrach Fuscocephaloziopsis catenulata posiada zaskakująco mało stanowisk publikowanych (Duda & Váňa, 1986; Górski & Váňa, 2014; Hazslinszky, 1885; Klama, 2008; Krupa, 1882; Limpricht, 1877a; Schiffner, 1911; Szweykowski, 1960; Szyszyłowicz, 1884), w stosunku do rzeczywistych zasobów (P. Górski, npbl.). Z tego względu, w skali całego masywu roślina nie jest zagrożona (NT, Górski, 2020) i potencjalnie może mieć najwięcej stanowisk ze wszystkich tatrzańskich gatunków wskaźnikowych. W terenie często tworzy peryancja, rzadziej rozmnóżki, jeśli porasta kłodę to zazwyczaj tworzy duże populacje. Najwięcej wystąpień gatunku udokumentowano w pasach wysokościowych 901–1000 i 1101–1200 m n.p.m. (Rycina 3). Minimum wysokościowe F. catenulata w Tatrach przypada na 860 m n.p.m. (słowackie Tatry Zachodnie, Juráňova dolina, leg. P. Górski, 2013; Górski & Váňa, 2014), a maksimum na 1420 m n.p.m. (nowe stanowisko: Słowacja, Tatry Wysokie, Kôprová dolina, Niedźwiedzia Perć, rozkładająca się kłoda świerka, 49.17307°N, 19.98492°E, MGRS 34UDV2647, leg. P. Górski 521/2015, 16.09.2015, POZNB 2018).
Harpanthus scutatus (F. Weber & D. Mohr) Spruce
Liczba stanowisk publikowanych: (SL): TB – 10, TW – 2, TZ – 1, (PL): TW – 1, TZ – 1.
Płozikowiec tarczkowaty jest głównie epiksylem, rzadko notowany był na skałach. Występuje w obszarach górskich (wyłącznie w reglu dolnym) i na pogórzu. Najwięcej polskich stanowisk (19) zlokalizowanych jest w Bieszczadach Zachodnich (Klama, 2013b; Szweykowski, 1959; Szweykowski & Buczkowska, 1996). Ponadto, na pojedynczych lokalizacjach, roślinę tę odnotowano na Babiej Górze, w Beskidzie Niskim, Sądeckim, Śląskim Żywiecko-Orawskim, Gorcach, Górach Sanocko-Turczańskich, Pieninach i Górach Stołowych (Karczmarz, 1987; Klama, 1996, 2004, 2013a; Mamczarz, 1975, 1977, 1979; Mierzeńska, 1994; Mierzeńska & Drewniok, 2000; Rejment-Grochowska, 1950; Stebel et al., 2016; Szweykowski, 1953b, 1961, 1971). Poza obszarami górskimi, Harpanthus scutatus notowany był na Pogórzu Kaczawskim (Warta Bolesławiecka, leg. G. Limpricht; por. Müller, 1906–1911, s. 840) oraz w województwie świętokrzyskim (okolice Niekłania; por. Błoński 1890, s. 44). Warte podkreślenia jest, że oba ostatnie stanowiska są naskalne.
W Tatrach najwięcej stanowisk Harpanthus scutatus przypada na Tatry Bielskie (10; Duda & Váňa, 1978; Górski & Váňa, 2014; Limpricht, 1877a, b; Schiffner, 1911). W pozostałych częściach masywu notowany był bardzo rzadko (Klama, 2008; Duda, 1955a; Peciar, 1985; Szyszyłowicz, 1884). Najwięcej wystąpień płozikowca przypada na pas wysokościowy 901–1000 m n.p.m. (11 na 25 notowań; Rycina 3). Minimum i maksimum wysokościowe wynoszą odpowiednio 800 m (Słowacja, Tatry Zachodnie, Mihulčia dolina; Peciar, 1985) i 1200 m n.p.m. (Tatry Bielskie, stoki Hawrania, leg. L. Vajda, 1963; Duda & Váňa, 1978).
Lophoziopsis longidens (Lindb.) Konstant. & Vilnet [= Lophozia longidens (Lindb.) Macoun]
Liczba stanowisk publikowanych: (SL): TB – 0, TW – 13, TZ– 2, (PL): TW – 6, TZ – 3.
Czubek długozębny jest gatunkiem epifitycznym i epilitycznym, rzadziej epiksylicznym. W obrębie pasa reglowego porasta nasady pni żywych (i martwych stojących) świerków, zaś w pobliżu potoków często spotykany jest na głazach i martwym drewnie (przy wodzie). Stanowiska wysokogórskie są naskalne. W Polsce Lophoziopsis longidens występuje w Sudetach i Przedgórzu Sudeckim (Góry Stołowe, Góry Bialskie, Śnieżnik Kłodzki, Masyw Ślęży), Karpatach (Tatry, Babia Góra, Gorce, Skalice Nowotarskie i Spiskie Beskid Żywiecko-Orawski, Beskid Śląski, Bieszczady Zachodnie) oraz na niżu, w północno-wschodniej części (Górski & Váňa, 2014; Klama, 1996, 2002a, 2002b, 2004, 2013a, b; Koła 1963, Ochyra & Cykowska, 2008; Rejment-Grochowska, 1950; Szweykowski, 1953b, 1956; Szweykowski & Buczkowska, 1996; Wojterski, 1954).
W profilu pionowym Tatr Lophoziopsis longidens posiada najwięcej notowań w zakresach wysokościowych 1301–1400 i 1201–1300 m n.p.m. (w pasie reglowym), a obszarze wysokogórskim – 1701–1800 m n.p.m. (Rycina 3). Maksimum wysokościowe wynosi 2480 m n.p.m. (Słowacja, Tatry Wysokie, południowe stoki Krywania, leg. P. Górski, 2013; Górski & Váňa, 2014), minimum – 876 m (nowe stanowisko: Słowacja, Tatry Zachodnie, Trnovská dolina, w pobliżu potoku Trnovečanka, 49.13798°N, 19.74278°E, MGRS 34UDV0843, leg. P. Górski 546/2015, 19.09.2015, POZNB 3349).
Metzgeria violacea (Ach. ex F.Weber & D.Mohr) Dumort.
Liczba stanowisk publikowanych: (SL): TB – 0, TW – 1, TZ– 0, (PL): TW – 0, TZ – 2.
Widlik krzaczkowaty jest bardzo rzadkim i zagrożonym wyginięciem (CR; Klama & Górski, 2018) gatunkiem epifitycznym rosnącym na korze buków. W Polsce występuje na Pomorzu Zachodnim (Górski, 2013b; Koppe & Koppe, 1940; Szweykowski, 1968; Szweykowski & Koźlicka, 1966), Beskidzie Sądeckim (Mamczarz, 1975, 1977), Tatrach (Duda, 1965; Górski & Váňa, 2014; Mickiewicz, 1965), Pogórzu Dynowskim (Armata, 2009) i Bieszczadach Zachodnich (Szweykowski, 1959; Szweykowski & Buczkowska, 1996; Tobolewski & Glanc, 1960).
W Tatrach, jako pewne uznać można tylko trzy stanowiska Metzgeria violacea: 1) Tatry Wysokie, Słowacja, bez lokalizacji i wysokości, leg. V. Greschik, 4.1883, det. J. Duda (Duda, 1965), 2) Dolina Olczyska, brak szczegółowej lokalizacji i wysokości (Mickiewicz, 1965), 3) przy drodze Oswalda Balzera pomiędzy Chłabówką a Cyrhlą, na korze buka, wys. 960 m n.p.m., leg. P. Górski, 2012 (Górski & Váňa, 2014). Kilka stanowisk tego gatunku podaje Szyszyłowicz (1884), które jednak były błędnie oznaczone (por. Szweykowski, 2006).
Porella arboris-vitae (With.) Grolle
Liczba stanowisk publikowanych: (SL): TB – 0, TW – 0, TZ– 0, (PL): TW – 0, TZ – 3.
Parzoch pieprzowy jest gatunkiem epifitycznym i epilitycznym. Jego stanowiska znane są głównie z obszarów górskich (Sudety i Karpaty), nieliczne rozproszone są na niżu (por. Szweykowski, 1958). Poza Tatrami podawany był z Gór Stołowych (Szweykowski, 1953b), Babiej Góry (Klama, 2004), Beskidu Sądeckiego (Mamczarz, 1970), Beskidu Śląskiego (Mickiewicz, 1965; Rejment-Grochowska, 1950), Gorców (Górski et al., 2015; Mierzeńska, 1994), Pienin (Szweykowski, 1961; Vončina et al., 2023), Beskidu Niskiego (Karczmarz, 1987), Bieszczad (Buczkowska & Bączkiewicz, 2010; Klama, 2013b; Mickiewicz, 1965; Szweykowski, 1959; Szweykowski & Buczkowska, 1996), Pogórza Karpackiego (Armata, 2011), Roztocza (Karczmarz, 1967; Szweykowski, 1956, 1957), Ojcowskiego Parku Narodowego (Pałkowa, 1961) i Mazowsza (Rejment, 1937).
W Tatrach znane są tylko trzy stanowiska: z polskiej części Tatr Zachodnich, z doliny Kościeliskiej (Krupa, 1882) oraz Chochołowskiej (Szweykowski, 1960; Szyszyłowicz, 1884). Wszystkie wystąpienia tej rośliny są naskalne. W 2021 odnaleziono kolejne, 2 stanowiska tej rośliny, także epilityczne (Tatry Zachodnie, P. Górski, npbl.). Jak dotychczas w Tatrach nie ma wystąpień tego gatunku na korze drzew pomimo dostępności potencjalnych siedlisk (głównie buka i jawora).
Scapania apiculata Spruce
Liczba stanowisk publikowanych: (SL): TB – 2, TW – 0, TZ– 2, (PL): TW – 0, TZ – 2.
Skapanka spiczasta jest obligatoryjnym epiksylem, bardzo rzadkim i zagrożonym w Polsce (CR; Klama & Górski, 2018). Poza Tatrami znana jest zaledwie z kilku stanowisk położonych w Beskidzie Śląskim i Białowieży (leg. T. Wiśniewski; Hausbrandt, 1949), Beskidzie Sądeckim (Mamczarz 1975, 1977), Bieszczadach (Szweykowski, 1959; Szweykowski & Buczkowska, 1996) oraz okolicach Giżycka na Mazurach (Koppe, 1931). W polskiej części Tatr znane są tylko 2 stanowiska o bardzo ogólnie określonej lokalizacji: Nosal (leg. J. Żebrowska; Hausbrandt, 1949) oraz Dolina Strążyska (Krupa, 1888). Po stronie słowackiej Tatr Scapania apiculata ma 4 stanowiska (Szyszyłowicz, 1884; Šmarda, 1961). Stanowisko podawane przez Hausbrandt (1949) z Białowieży nie zostało potwierdzone w późniejszych badaniach (Klama, 2002a). W 2013 roku nie odnaleziono tego gatunku w słowackich Tatrach Zachodnich w miejscu opisywanym przez Szyszyłowicza (1884) (por. Górski, 2020). Natomiast w 2022 roku potwierdzono (w odniesieniu do doliny górskiej) stanowisko Krupy (1888) [nowe stanowisko: Dolina Wielkiej Równi (górna część Doliny Strążyskiej), w środkowej części, martwe drewno powyżej okresowego potoku, 49.258202°N, 19.934172°E, MGRS 34UDV2256, wys. 1183 m n.p.m., leg. P. Górski 632/2022, 11.08.2022, POZNB 4971].
Scapania carinthiaca J.B.Jack ex Lindb.
Liczba stanowisk publikowanych: (SL): TB – 1, TW – 0, TZ– 1, (PL): TW – 0, TZ – 1.
Skapanka karyncka jest obligatoryjnym epiksylem. W Polsce podawana była tylko z jednego stanowiska odnalezionego w Tatrach Zachodnich przez J. Szweykowskiego w 1959 roku (Szweykowski, 1960). Po słowackiej stronie masywu są jeszcze dwa historyczne notowania tej rośliny – w Tatrach Bielskich (Šmarda, 1961) i Zachodnich (Duda, 1955a, 1955b). Poszukiwania w terenie na dawnych lokalizacjach wskazują, że gatunek ten nie występuje w tym masywie (por. Górski, 2020). W skali Polski Scapania carinthiaca jest krytycznie zagrożona (CR; Klama & Górski, 2018).
. Uwagi końcowe
Zaproponowana lista 11 gatunków wątrobowców wskaźnikowych lasów zawiera mszaki spełniające przyjęte kryteria dla reliktów lasów puszczańskich. Ocenę naturalności kompleksu leśnego z wykorzystaniem wątrobowców należy prowadzić w połączeniu z dobrą znajomością biologii (autekologii) poszczególnych gatunków, a tym samym umiejętnością ich odnalezienia. Tylko odpowiednio rozległy las zapewni mozaikę zmiennych mikrosiedliskowych, w których należy poszukiwać poszczególnych gatunków. W lasach górskich dodatkowym czynnikiem wpływającym na obecność reliktów jest wyniesienie nad poziom morza. Inaczej wygląda ich obecność w dolnej części regla dolnego, inaczej przy górnej granicy lasu. Wydaje się, że z wysokością maleje liczba gatunków reliktowych. Nie należy więc oczekiwać licznej reprezentacji roślin tej grupy, a tym samym z jej bogactwa wyciągać wniosków o naturalności lasu. Na podstawie wstępnych badań w wybranych kompleksach leśnych polskiej części Tatr Wysokich zauważyć można, że w obrębie jednego obszaru leśnego może występować od 2 do 5 gatunków wskaźnikowych. Dobre różnice w stopniu naturalności oddaje analiza częstości występowania gatunków reliktowych, w której uwzględniamy wszystkie wystąpienia (nawet w bliskiej odległości, np. leżące obok siebie kłody, czy rosnące blisko drzewa) wszystkich gatunków wskaźnikowych. W Tatrach Wysokich są kompleksy leśne, które mają ten parametr na poziomie 26 (łączne notowania wszystkich gatunków wskaźnikowych ze wszystkich mikrosiedlisk), a są też takie, gdzie ta wartość sięga 90 (P. Górski, npbl.). To pokazuje, że w lasach górskich pobieżne badania nastawione tylko na stwierdzenie obecności lub braku cennych gatunków niewiele pokazują.
Z zaproponowanych 11 gatunków wątrobowców aż 5 będzie miało aktualnie niewielki walor praktyczny ze względu na swoją wyjątkową rzadkość. Są to: Frullania jackii, Metzgeria violacea, Porella arboris-vitae, Scapania apiculata oraz Scapania carinthiaca. Według Szweykowskiego & Buczkowskiej (1996) Porella arboris-vitae była w Bieszczadach, w latach 50-tych XX wieku, pospolitym gatunkiem na korze buków, razem z Metzgeria violacea i Frullania fragilifolia. Natomiast w latach 90-tych, w momencie redagowania pracy dotyczącej flory wątrobowców tego masywu, rośliny te niemal całkowicie ustąpiły (Szweykowski & Buczkowska, 1996). Z wymienionych epifitów, po kilku latach odnaleziono w Bieszczadach tylko P. arboris-vitae (Buczkowska & Bączkiewicz, 2010). Pytanie o występowanie tego gatunku oraz Frullania jackii na bukach lub jaworach w Tatrach jest otwarte. W tej chwili wyciąganie wniosków dotyczących występowania najrzadszych epifitów jest przedwczesne ze względu na brak szerzej zakrojonych badań i poszukiwań. Ostatnie były przeprowadzone w II połowie XX wieku, obejmowały buka w całym obszarze zasięgu, a Tatry były jednym z wielu regionów objętych inwentaryzacją (Mickiewicz, 1965).
Prezentowana lista 11 gatunków reliktowych lasów puszczańskich może być stosowana do oceny stopnia naturalności kompleksów leśnych we wszystkich obszarach górskich i podgórskich Polski. Na pozostałym terenie można dodatkowo wykorzystać gatunki, które w Tatrach są dość częste, ale poza nimi występują dużo rzadziej i związane są z lasami o wyższym stopniu naturalności. Do grupy tej zaliczyć można następujące wątrobowce: Bazzania trilobata (L.) Gray [zagrożenie w Polsce (PL): LC, zagrożenie w Tatrach (T): LC; Klama & Górski, 2018; Górski, 2020], Calypogeia suecica (Arnell & J. Perss.) Müll.Frib. [PL: VU, T: LC], Fuscocephaloziopsis leucantha (Spruce) Váňa & L. Söderstr. [PL: NT, T: LC], Frullania tamarisci (L.) Dumort. (stanowiska epifityczne) [PL: VU, T: LC], Lophozia ascendens (Warnst.) R.M. Schust. [PL: VU, T: LC] oraz Syzygiella autumnalis (DC.) K. Feldberg, Váňa, Hentschel & Heinrichs [PL: LC, T: VU]. Prezentowana powyżej grupa (umownie określona jako lista B, w przeciwieństwie do A – por. Tabela 1) powinna być konstruowana każdorazowo z uwzględnieniem lokalnych uwarunkowań w rozmieszczeniu i zasobach poszczególnych gatunków. Trudno nadal uwzględniać, także na niżu, wystąpienia Plagiochila asplenioides za wskaźnikowe dla lasów puszczańskich. Roślina ta związana jest z siedliskami o różnym stopniu naturalności i nie jest rzadkim gatunkiem. Wydaje się także, że waloru wskaźnikowego nie posiadają epifityczne notowania Porella platyphylla (L.) Pfeiff. Wątrobowiec ten uznawany był blisko 20 lat temu za wymierający (kategoria E; Klama, 2006), a obecnie ma wiele stanowisk (por. Górski & Fudali et al., 2017 oraz Górski & Smoczyk et al. 2016 i cytowana tam literatura; portal iNaturalist). Część z nich stwierdzono na siedliskach antropogenicznych tj. parki czy nawet drzewostany z Robinia pseudoacacia na hałdach pogórniczych (leg. M.K. Dyderski, 2016, por. Górski & Fudali et al., 2017). Jako gatunek wskaźnikowy lasów o wysokim stopniu naturalności nie powinien być wskazywany także wątrobowiec Nowellia curvifolia (Dicks.) Mitt. (por. Stebel & Vončina, 2020; Ojcowski Park Narodowy). I podobnie jak w przypadku P. platyphylla, także N. curvifolia była wcześniej rośliną zagrożoną wyginięciem (kategoria V; Klama, 2006). Aktualnie wątrobowiec ten jest dość częsty (regionalnie pospolity) na niżu i w górach, także w leśnych zbiorowiskach zastępczych, a więc układach antropogenicznych (por. New distributional data on bryophytes of Poland pod redakcją P. Górskiego i A. Rusińskiej, części 1, 2, 3, 4, 8, 13, 16 i cytowana tam literatura). Trudno określić przyczyny tak odmiennego obrazu rozmieszczenia obu gatunków w ciągu ostatnich lat. Z całą pewnością kluczowym problemem jest brak współczesnych danych terenowych, które byłyby kontynuacją intensywnych eksploracji całego obszaru Polski jakie miały miejsce w II połowie XX wieku.
Podziękowania
Badania były finansowane w ramach Funduszu Leśnego PGL Lasy Państwowe. Serdecznie dziękuję dr. inż. Antoniemu Ziębie (Tatrzański Park Narodowy, Dział Badań Naukowych i Planowania Ochrony Przyrody) za wskazanie najcenniejszych obszarów lasów puszczańskich w Tatrach oraz pomoc w organizacji badań terenowych.