. Introduction
Drought limits plant growth and development, posing a significant challenge to crop productivity (Bakır et al., 2022). Under drought stress, crop yields are adversely affected (Hasan et al., 2023), and the production of harmful reactive oxygen species (ROS) escalates rapidly. If not efficiently neutralized by the plant antioxidant defense mechanisms, these ROS can lead to extensive cellular damage (Hasan et al., 2021a; Alabdallah et al., 2021). This damage includes harm to cellular membranes and components in plants (Jahan et al., 2022; Hasan et al., 2021b). For example, during drought stress, elevated levels of MDA are often utilized as markers of oxidative damage to the plant cell (Khan et al., 2017; Abdelaal et al., 2022). Moreover, plants have evolved antioxidant defense mechanisms, particularly by enhancing the activities of SOD and CAT enzymes, to control ROS levels during drought stress (Rahman et al., 2022; Contreras-Oliva et al., 2022).
Coffee is a widely enjoyed beverage, ranking as the second most traded commodity worldwide (Contreras-Oliva et al., 2022). However, coffee yields have come under threat due to an escalation in both the frequency and severity of drought conditions (Silva et al., 2022). This situation poses potential risks not only to the quantity of coffee produced but also to its quality (DaMatta et al., 2018). Drought is a significant environmental challenge, and its adverse effects have been substantial, resulting in an alarming 80% reduction in global coffee production (Merga et al., 2023). Therefore, it is important to formulate strategies aimed at bolstering coffee’s resilience to drought and enhancing its overall productivity.
The application of titanium dioxide nanoparticles (TiO2NPs) and ascorbic acid (AsA) has demonstrated the ability to enhance the drought stress tolerance in plants (Celi et al., 2023; Karvar et al., 2022). Titanium dioxide (TiO2), found in everyday items like plastics and cosmetics, poses environmental risks, potentially leading to human health concerns (Menard et al., 2011). Toxicity is a major issue in the environment, directly related to various factors, including chemical compounds (Melo et al., 2022). Human exposure to TiO2 can occur through various routes, such as consuming contaminated water from rivers where individuals have used sunscreen. In agriculture, TiO2 in pesticides and fertilizers enhances plant growth and soil fertility, with its photocatalytic properties aiding in pesticide breakdown (Menard et al., 2011). Its agricultural use may result in widespread environmental distribution, posing risks through contaminated food crops or fish (Jovanovc, 2015). However, TiO2NPs have garnered significant attention due to their excellent compatibility with living organisms, low toxicity, strong capacity to neutralize harmful free radicals, and high availability for biological processes (Wang et al., 2022). TiO2NPs are considered to have the potential to enhance crop productivity (Li et al., 2018). For instance, TiO2NPs with melatonin increase the rate of photosynthesis, the activity of rubisco, the potential of antioxidant enzymes, and the production of chlorophyll in plants (Sheikhalipour et al., 2023). Conversely, AsA serves as a crucial growth regulator that safeguards plant tissues from the harmful impacts of oxygen radicals (Celi et al., 2023). For instance, AsA acts as a co-factor for enzymes responsible for hormone production and the regeneration of enzymatic antioxidants in plants (Castro et al., 2023).
However, despite these advancements, there has been limited research into the combined use of ascorbic acid and TiO2NPs in coffee plants facing drought stress. Therefore, it is urgent to explore these novel findings by investigating the impact of ascorbic acid and TiO2NPs on coffee plants. This has led to the hypothesis that applying ascorbic acid along with TiO2NPs could be a valuable approach to enhance the drought tolerance of coffee plants. Consequently, our objective was to assess the following: (i) The effectiveness of ascorbic acid and TiO2NPs in mitigating the detrimental effects of drought stress on coffee plant growth. (ii) The influence of ascorbic acid and TiO2NPs on physicochemical characteristics that contribute to improved crop yield. (iii) The possibility to cultivate coffee plants with reduced water consumption without experiencing a significant reduction in yield.
. Materials and methods
. Growth conditions
This study was designed to evaluate the effect of TiO2NPs and AsA on the drought tolerance of coffee plants (Coffea arabica cv. Khawlani) in controlled conditions. The experiments were conducted in a controlled greenhouse at the Department of Biology, Imam Abdulrahman Bin Faisal University, while the biochemical analysis was performed at Umm Al-Qura University, Saudi Arabia. Coffee seeds used in the study were sourced from the Jazan area in Saudi Arabia (latitude 16.8894°N, longitude 42.5706°E). To prepare the coffee seeds for planting, they were first sterilized with a 1% sodium hypochlorite (NaOCl) solution for 10 minutes and then rinsed with tap water for an additional minute. The treated seeds were then transferred into plastic pots, each filled with a mixture of garden soil.
The greenhouse maintained day/night temperatures of 22°C/16°C and relative humidity in the range of 60–70%. The plants were watered twice a week to reach a soil moisture level corresponding to 100% field capacity (FC), which served as the control condition. Drought stress was induced by adjusting watering to achieve 50% FC and 20% FC, simulating moderate and severe drought conditions, respectively, for two weeks. TiO2NPs and AsA were applied through foliar spray using a hand-held sprayer to ensure even distribution. The spray was applied starting in the 25th up to 30th weeks during the drought period, with concentrations of 10 ppm for TiO2NPs and 50 ppm for AsA, combining 10 ppm TiO2NPs and 50 ppm AsA for each application per plant. Each plant received a total spray volume of 30 mL per application, applied twice a week during the drought period. The coffee plants representing a mature growth stage of seedlings were harvested at 30 weeks for physiological and biochemical analyses.
. TiO2NP preparations and experimental treatments
We synthesized TiO2NPs using sol-gel techniques. Titanium tetraisopropoxide (TIP), with a purity level of 99.9%, was combined with acetic acid (purity ≥ 99.7%) and ethanol (purity ≥ 99.9%) at ambient room temperature. The mixture was prepared in a specific molar ratio of 0.5:0.05:4.5 for TIP, acetic acid, and ethanol, respectively. The solution was then continuously stirred for 1.5 hours to ensure proper blending and homogenization. Following this, the resulting mixture was subjected to drying in an oven set at 100°C, where it was allowed to dry for 12 hours. Subsequently, the dried material underwent thermal treatment, reaching temperatures in the range of 500 to 550°C in a furnace for 2 hours, with a heating rate of approximately 0.16°C per minute. The size and shape of the heated samples were characterized as spherical with dimensions ranging from 30 to 55 nm using a scanning electron microscope (SEM). In this experiment, we employed four different treatments: the control group, 10 ppm TiO2NPs, 50 ppm AsA, and a combination of 10 ppm TiO2NPs with 50 ppm AsA.
. Measurements of growth parameters
Coffee plant shoot and root lengths were measured using the metric system, and the units were reported in centimeters (cm). On the days of harvest, length-related attributes were determined using a measuring tape after the plants were gently uprooted. The roots and shoots were rinsed with tap water, and their lengths were measured in centimeters (cm). The weight of fresh coffee plant shoots and roots were determined using a digital balance (HR–200). The dry weights of the shoots and roots were recorded after they had been dried at 60°C.
. Relative water content (RWC)
The RWC in the coffee plants was measured following the protocol described by Barrs and Weatherley (1962). It was determined using the following formula: RWC (%) = [(Fresh weight, FW – Dry weight, DW)/(Turgid weight, TW − Dry weight, DW)] × 100. To obtain the TW, the leaves were submerged in distilled water for 6 hours, allowing them to reach full turgidity. The DW was measured after the leaves were thoroughly dried to remove all moisture content.
. MDA content measurements
The MDA concentration in the sample was quantified by following the protocol established by Heath and Packer (1968). To begin, 0.5 g of coffee fresh leaf tissue were ground and homogenized with 5 mL of a 10% trichloroacetic acid (TCA) solution. The homogenate was then subjected to centrifugation at 15,000×g for 10 minutes to separate the solid and liquid phases. After centrifugation, 2 mL of the supernatant were carefully collected and combined with 4 milliliters of a 0.6% thiobarbituric acid (TBA) solution. The resulting mixture was thoroughly blended to ensure uniformity. The mixture was subsequently heated to 100°C for 15 minutes to promote the reaction between MDA and TBA, which produces a colored complex. After heating, the reaction was immediately stopped by rapidly cooling the mixture on ice. The absorbance of this complex was measured at 532 nm to quantify the MDA concentration, as this wavelength is specific for the MDA–TBA complex. In addition, absorbance at 600 nm was measured to correct for any non-specific turbidity and light scattering in the sample, ensuring accuracy. The extinction coefficient used for the MDA–TBA complex was 155 mM-1 cm-1.
. Proline content determination
Proline content was determined by the protocol of Bates et al. (1973). 0.2 g of fresh leaf tissue was first homogenized with 10 mL of a 3% (w/v) sulfosalicylic acid solution. The homogenate was then centrifuged at 15,000×g for 10 minutes to separate the solid and liquid phases. After centrifugation, 2 mL of the clear supernatant were carefully transferred and mixed with 2 mL of glacial acetic acid and 2 mL of an acid ninhydrin solution in test tubes. The tubes were then placed in a boiling water bath for 1 hour to allow the reaction to take place, which facilitated the formation of a chromophore. Following the heating step, 4 mL of toluene were added to extract the reaction product into the organic phase. The absorbance of the toluene phase containing the chromophore was measured at 520 nm.
. Determination of total soluble carbohydrates (TSC)
TSC were determined following the protocol established by Dubois et al. (1956). In this procedure, anthrone was prepared by dissolving it in 45 mL of 95.5% (v/v) sulfuric acid. For the reaction, a mixture was created by combining 50 µL of the alcoholic extract with 950 µL of deionized water, resulting in a total volume of 1000 µL. To this reaction mixture, 2000 µL of the prepared anthrone solution were gradually added while keeping the reaction vessel in an ice bath to prevent overheating. After the addition, the mixture was incubated in a water bath at 100°C for 3 minutes to promote the reaction between anthrone and the carbohydrates in the extract. Once the incubation was complete, the mixture was allowed to cool to room temperature naturally. Glucose was used as the reference standard for this procedure. The absorbance of the resulting solution was measured at 620 nm using a spectrophotometer to quantify the carbohydrate content in the sample.
. Antioxidant activity measurements
A fresh coffee leaf sample weighing 0.3 g was placed in 5 mL of phosphate buffer, which had a concentration of 50 mmol/L and a pH of 7.0. To enhance the extraction efficiency, 10 g/L of polyvinylpyrrolidone (PVP) was added. The mixture was then subjected to centrifugation at 15,000×g for 20 minutes to separate the solid plant material from the liquid extract. After centrifugation, the supernatant was carefully collected and used for the preliminary analysis of enzyme activities.
The activity of SOD was determined based on the method described by Hasan et al. (2021b), with slight modifications. The reaction mixture for SOD activity included 0.1 mL of 60 μM riboflavin, 1.5 mL of 26 µM L-methionine, 1.0 mL of 225 µM nitroblue tetrazolium (NBT), 0.75 mL of 1% Triton-X, and 15 µL of 2 mM EDTA, all dissolved in 27.0 mL of sodium-potassium phosphate (NaKPi) buffer at pH 7.8. To this reaction mixture, 0.5 mL of the enzyme extract was added to make a total volume of 3.0 mL. The tubes containing the mixture were incubated at 25°C for 10 minutes, shielded from direct light. The reaction was initiated by exposing the tubes to light from a 15-watt fluorescent lamp (pre-heated, 6500 K). The reaction was allowed to proceed for 20 minutes under light and the SOD activity was quantified by measuring the absorbance of the reaction at 560 nm, which reflects the enzyme ability to inhibit the photochemical reduction of NBT.
CAT activity was evaluated following the method outlined by Aebi (1984). In this procedure, 20 µL of the enzyme extract was mixed with 830 µL of potassium phosphate buffer (50 mM, pH 7.0). The enzymatic reaction was initiated by adding 150 µL of 0.05M hydrogen peroxide (H2O2). The reaction progress was monitored by measuring the decrease in absorbance at 240 nm over one minute, reflecting the CAT activity as it breaks down H2O2 into water and oxygen.
. Statistical analysis
In this study, a Completely Randomized Design (CRD) was implemented for the treatment application. A one-way analysis of variance (ANOVA) was conducted to evaluate the data, ensuring that any differences between groups were statistically analyzed. To further explore and compare the specific group differences, Fisher’s Least Significant Difference (LSD) was employed, with a significance level set at p ≤ 0.05. All data processing and statistical calculations were carried out using Minitab-17 software.
. Results
Scanning electron microscopy (SEM) analysis was carried out to verify the successful synthesis of TiO2NPs and provided detailed insights into the morphology, size, and surface structure of the nanoparticles, confirming that the desired characteristics were achieved. After validation through SEM imaging, as illustrated in Figure 1, the synthesized TiO2NPs were incorporated into experimental studies aimed at exploring their effectiveness in reducing the adverse effects of drought stress on coffee plants.
Figure 1
SEM techniques visualize the successful synthesis of TiO2NPs by providing high-resolution imaging.

The growth of the coffee plants was observed to decrease significantly (p ≤ 0.05) when subjected to the drought stress conditions, specifically at 50% and 20% field capacity (FC). For instance, all morphological parameters of the coffee plants exhibited a notable decline (p ≤ 0.05) in response to the drought stress treatment (50% and 20% FC) (Figure 2, 3).
Figure 2
Impacts of TiO2NPs and AsA on the shoot and root lengths of coffee plants during drought stress conditions (50% FC and 20% FC). Statistically significant differences at the p ≤ 0.05 level are indicated by bars with different letter annotations.
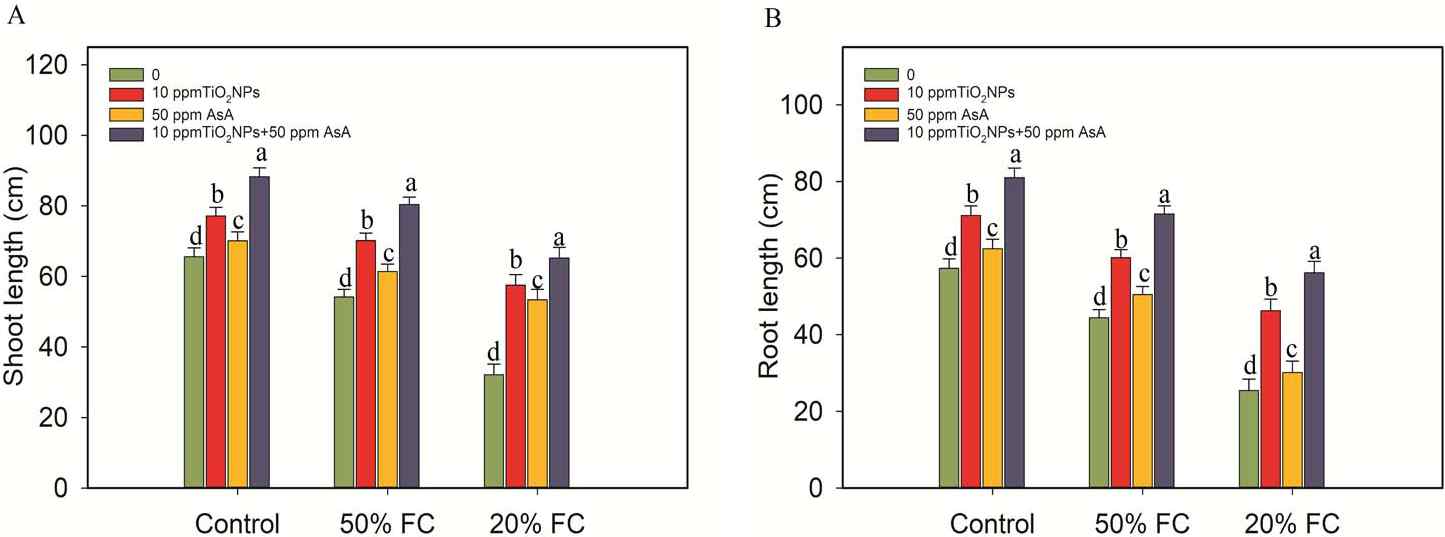
Figure 3
Impacts of TiO2NPs and AsA on the fresh weight of shoot and root and dry weight of shoot and root of coffee plants during drought stress conditions (50% FC and 20% FC). Statistically significant differences at the p ≤ 0.05 level are indicated by bars with different letter annotations.
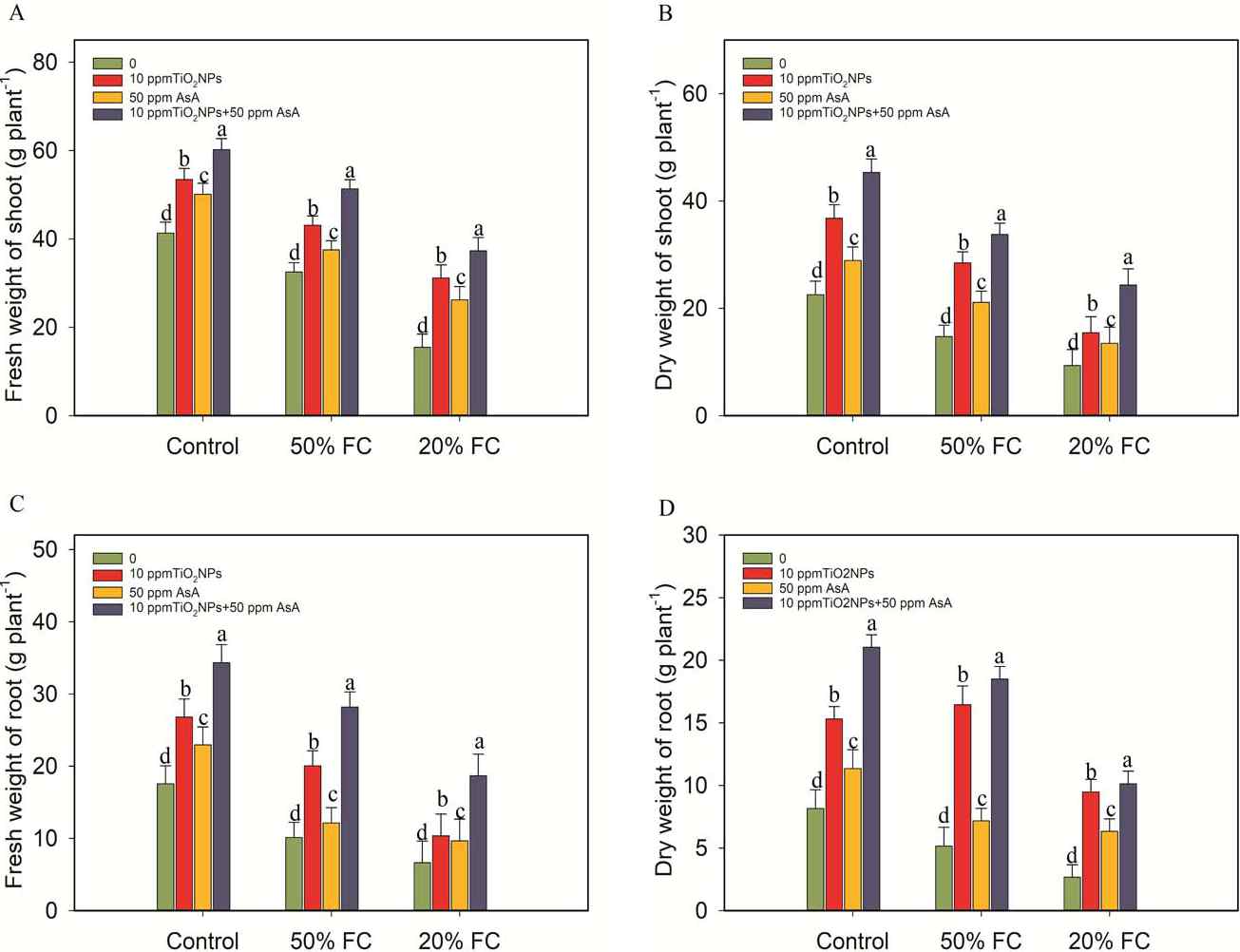
However, the diverse concentrations of TiO2NPs and AsA treatments (10 ppm TiO2NPs, 50 ppm AsA, and 10 ppm TiO2NPs + 50 ppm AsA) led to varying degrees of improvement in these growth parameters (Figure 2, 3).
The application of TiO2NPs and AsA treatments (10 ppm TiO2NPs, 50 ppm AsA, and 10 ppm TiO2NPs + 50 ppm AsA) resulted in increased RWC by 8%, 6%, and 13%, respectively, when compared to the control.
In contrast, the drought treatments at 50% and 20% FC resulted in a substantial reduction in the RWC in the coffee plants, with decreases of 25% and 40%, respectively, when compared to the control group. However, the application of the TiO2NPs and AsA treatments led to notable improvements in RWC. Specifically, TiO2NPs enhanced RWC by 13%, 8%, and 18%, while the AsA treatments increased it by 15%, 10%, and 19% in coffee plants subjected to drought stress at 50% and 20% FC, respectively. Furthermore, drought stress significantly (p ≤ 0.05) elevated the concentrations of organic compounds such as proline (Figure 4) and TSC (Figure 5) in the coffee plants.
Figure 4
Impacts of TiO2NPs and AsA on the relative water and proline content in coffee plants during drought stress conditions (50% FC and 20% FC). Statistically significant differences at the p ≤ 0.05 level are indicated by bars with different letter annotations.
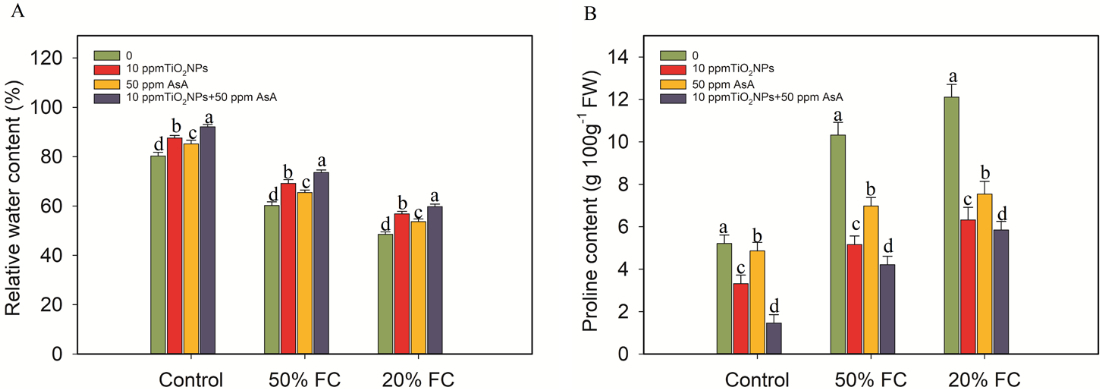
Figure 5
Impacts of TiO2NPs and AsA on the total soluble carbohydrates and MDA content in coffee plants during drought stress conditions (50% FC and 20% FC). Statistically significant differences at the p ≤ 0.05 level are indicated by bars with different letter annotations.
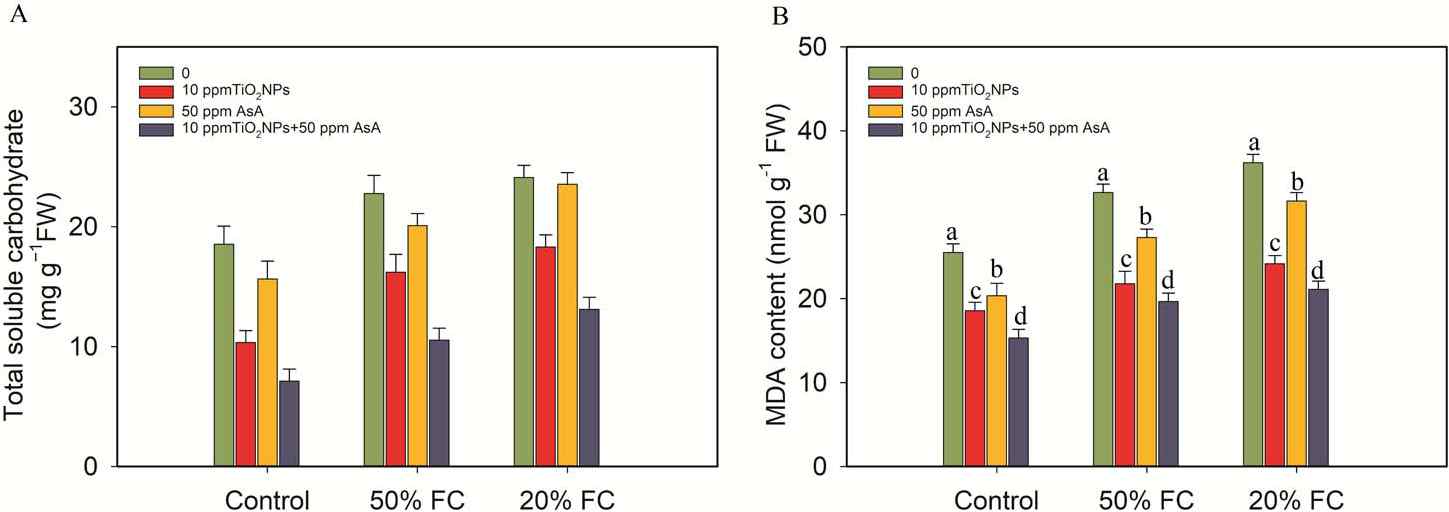
In contrast, when the coffee plants were treated with TiO2NPs and AsA, a notable reduction in proline content was observed. Specifically, the TiO2NP treatments led to a decrease in proline levels by 50%, 32%, and 59%, while the AsA treatments resulted in reductions of 48%, 38%, and 52%, compared to the proline levels in the drought-stressed coffee plants at 50% and 20% FC. Similarly, the levels of TSC also dropped significantly with the application of TiO2NPs and AsA. TiO2NPs reduced TSC by 29%, 12%, and 54%, whereas the AsA treatments led to reductions of 24%, 2%, and 45%, respectively, when compared to the TSC levels in the drought-stressed plants at 50% and 20% FC (Figure 5).
In the drought stress conditions at 50% and 20% FC, there was a substantial increase of 22% and 29%, respectively, in the malondialdehyde (MDA) levels, compared to the control. However, the application of the TiO2NPs and AsA treatments (10 ppm TiO2NPs, 50 ppm AsA, 10 ppm TiO2NPs + 50 ppm AsA) resulted in reductions of 33%, 16%, 40%, and 33%, 13%, 42%, respectively, in the MDA levels, compared to the coffee plants under drought stress (Figure 5).
The drought treatments at 50% and 20% FC caused a marked rise in the activities of SOD and CAT in the coffee plants. SOD activity increased by 77% and 89%, while CAT activity showed increases of 55% and 67%, respectively, when compared to the control plants. These results suggest that drought stress triggers a significant enhancement of antioxidant enzyme activity. However, the application of the TiO2NPs and AsA treatments, including 10 ppm TiO2NPs, 50 ppm AsA, and 10 ppm TiO2NPs combined with 50 ppm AsA, further increased SOD activity by 44%, 51%, 47%, and 27%, 48%, 18%, respectively (Figure 6).
Figure 6
Impacts of TiO2NPs and AsA on the SOD and CAT activity of coffee plants during drought stress conditions (50% FC and 20% FC). Statistically significant differences at the p ≤ 0.05 level are indicated by bars with different letter annotations.
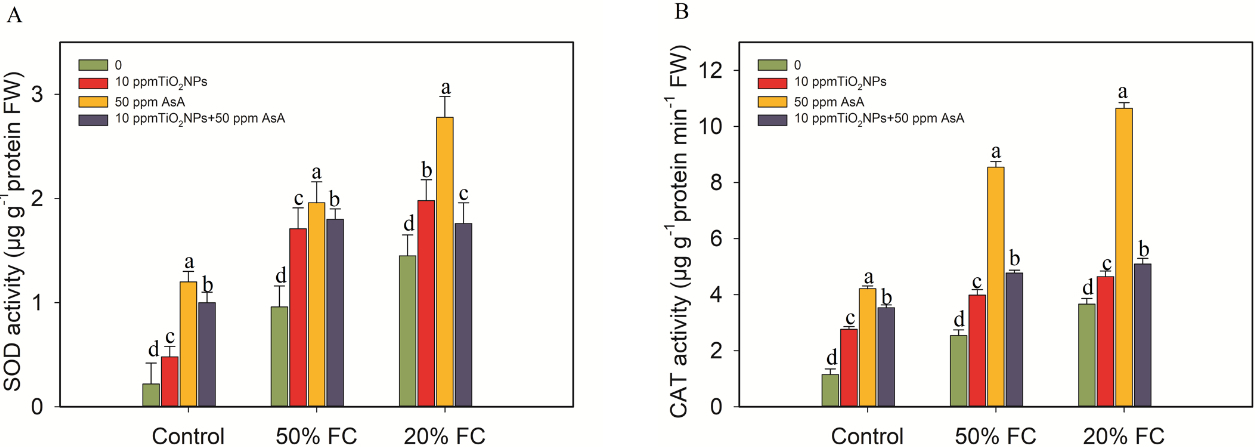
CAT activity showed a significant increase when the coffee plants were treated with the various concentrations of TiO2NPs and AsA, in comparison to those subjected to drought stress at 50% and 20% FC. Specifically, CAT activity rose by 36%, 70%, and 47% following the application of TiO2NPs, while the AsA treatments led to increases of 21%, 66%, and 28%, respectively (Figure 6).
. Discussion
Drought stress significantly restricts plant growth, resulting in a decrease in leaf water potential (ψ) (Yao et al., 2021; Waseem et al., 2021; Hasan et al., 2022; Liu et al., 2023). Conversely, when plants are treated with AsA and TiO2NPs, their ability to withstand water scarcity is enhanced, mitigating the harmful impacts of this stress and preserving adequate turgor pressure in the leaves (Celi et al., 2023). The application of AsA and TiO2NPs to coffee plants promotes increased drought tolerance in young coffee plants (Figure 7), as evidenced by the significant improvements in various growth parameters.
Figure 7
Schematic presentation of enhancement of the drought tolerance of coffee plants by titanium dioxide nanoparticles (TiO2NPs) and ascorbic acid (AsA).
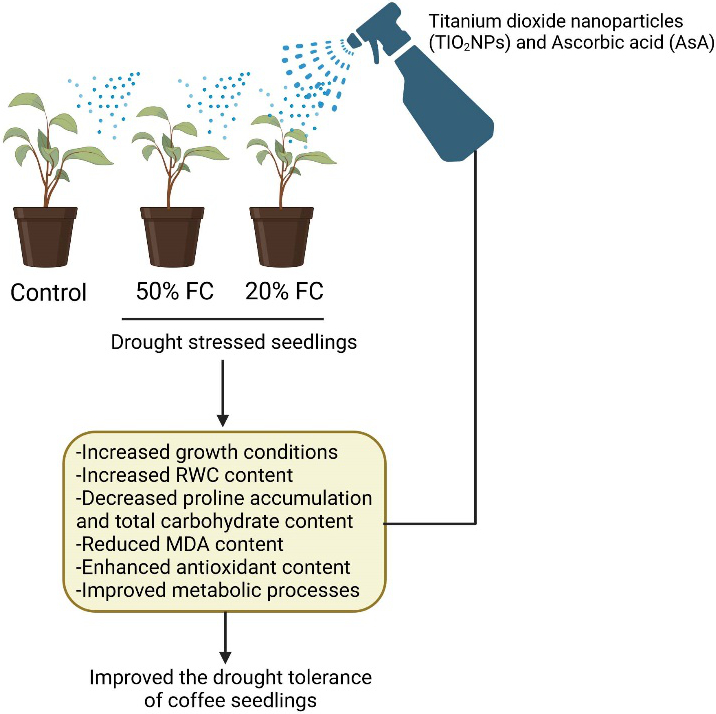
These results align with the findings reported by Ostadi et al. (2022) and Khazaei and Estaji (2020), both of which emphasize the important role that TiO2NPs and AsA play in mitigating stress responses in plants (Figure 7). The RWC in leaves serves as an indicator of leaf hydration (Patanè et al., 2022). In our recent studies, as expected, the relative water content (RWC) reached its highest levels in control conditions and its lowest levels in conditions of drought. Nevertheless, TiO2NPs and AsA resulted in an increase in RWC. Our analysis of the impact of TiO2NPs and AsA on coffee plants in drought stress conditions revealed that the beneficial effects of these substances on the studied characteristics can be attributed to their enhanced efficiency in water absorption. This, in turn, contributes to an improvement in turgor pressure and helps maintain water balance. The enhancement of RWC through the application of TiO2NPs and AsA aligns with findings in studies involving sweet pepper (Khazaei et al., 2020) and sweet corn (Karvar et al., 2022), which share similarities with the current research.
Small organic molecules, like sugars, amino acids, organic and aromatic compounds, have the ability to control the cell’s osmotic potential, which helps enhance water absorption during periods of drought stress (Jahan et al., 2023). Furthermore, these compounds serve to safeguard enzymes, biological membranes, and the photosynthetic system from oxidative harm (Khazaei et al., 2020). The current study demonstrates that coffee plants, when exposed to drought conditions, exhibit an increase in proline and soluble carbohydrates. In the context of drought-induced oxidative stress, there was an observed rise in proline levels in both pepper (Khazaei et al., 2020) and sweet corn (Karvar et al., 2022) plants and carbohydrate levels in pansy plants (Oraee et al., 2020), which is consistent with the findings in our present research. A related study demonstrated that multiple drought occurrences significantly affected the metabolic profile, including amino acids and carbohydrates, in coffee plants, indirectly aligning with our findings (Khazaei et al., 2020). Besides the amino acids and carbohydrates, other primary and secondary metabolites such as aromatic compounds and organic acids altered significantly in water-stress conditions, some previous studies were found parallel to our current results (Rodrigues et al., 2021; Guedes et al., 2018). In our study on coffee plants, the application of TiO2NPs and AsA in drought conditions resulted in decreased levels of proline and carbohydrates. This reduction suggests that an alternative protective mechanism may be in operation, potentially linked to the enhanced activation of antioxidative pathways by these treatments, which could mitigate oxidative stress more directly than osmotic adjustment.
In coffee plants, our study suggests that TiO2NPs and AsA may influence the plant’s oxidative balance by potentially modulating the enzymatic antioxidative defense system, rather than directly regulating already formed free radicals (Karamian et al., 2020). This could contribute to enhanced stress tolerance by mitigating the production of ROS, although direct measurements of ROS levels were not conducted in this study. The stability of the cell membrane is a crucial indicator of cellular integrity, especially in response to drought stress. The MDA content is a key marker for assessing cellular damage in plants, with elevated levels indicating increased membrane damage and compromised membrane integrity and function (Ahmed et al., 2023; Li et al., 2023; Sousa et al., 2023). In the present study, drought stress resulted in a notable rise in the MDA content, which indicates substantial disruption of the cell membrane. Nonetheless, the application of TiO2NPs and AsA effectively mitigated these detrimental effects. The current findings indicate that TiO2NPs and AsA play a critical role in safeguarding membrane stability in adverse environmental conditions. These observations are in line with previous studies on Verbascum nudicuale (Sousa et al., 2023) and pepper plants (Rodrigues et al., 2021), where TiO2NPs and AsA were shown to enhance plant tolerance to drought-induced membrane damage.
A widely employed strategy for counteracting the damaging impacts of ROS in plants involves the utilization of antioxidant mechanisms. These mechanisms are designed to safeguard plants from oxidative stress and include enzymes like superoxide dismutase (SOD), which converts superoxide into hydrogen peroxide (H2O2), and other enzymes such as catalase and peroxidase that break down H2O2 into water (H2O) and oxygen (O2) (Abdelaal et al., 2022). Thus, the processes mitigating oxidative stress in plants are intricately linked to the action of SOD, which plays a pivotal role in activating other antioxidant enzymes (Hasan et al., 2021b). Our findings show a continuous increase in SOD enzyme activity in coffee plants, which corresponds with prior studies reporting similar increases under drought stress in coffee plants (Sousa et al., 2023) and soybeans (Hasan et al., 2020). CAT plays a crucial role in the glutathione-ascorbate cycle, which is vital for detoxifying H2O2 produced by the SOD enzyme in various cellular compartments. Our findings reveal that the activity of CAT in coffee plants exhibited similar patterns of change as those observed with SOD. When the coffee plants were treated with TiO2NPs and AsA under drought stress, both SOD and CAT enzyme activities significantly increased. These results are consistent with earlier studies that showed that TiO2NPs and AsA treatments enhanced antioxidant enzyme activity in corn (Karvar et al., 2022), cucumber (Zahra et al., 2021), and pepper (Khazaei et al., 2020).
Based on the experimental results, it is plausible that the application of TiO2NPs and AsA led to an upregulation of both SOD and CAT activities, potentially linked to a reduction in the MDA content in the coffee leaves. This suggests that the combined use of TiO2 nanoparticles (TiO2NPs) and ascorbic acid (AsA) likely produces a synergistic effect by activating antioxidant defenses simultaneously. The enhanced activity of these key enzymes reflects their role in managing ROS and maintaining cellular stability, contributing to improved plant resilience in drought conditions. In addition, TiO2NPs enhance water uptake by boosting root system efficiency, helping to maintain turgor pressure during drought. Meanwhile, AsA, as a potent antioxidant, plays a key role in scavenging reactive oxygen species (ROS), particularly by detoxifying hydrogen peroxide (H2O2) generated under drought-induced oxidative stress. This complementary interaction improving water regulation via TiO2NPs and mitigating oxidative stress through AsA ultimately strengthens the drought tolerance of coffee plants.
. Conclusion
In summary, our study has revealed that TiO2NPs applied along with AsA leads to a substantial enhancement of physiological defense mechanisms in coffee plants. This combined application of TiO2NPs and AsA effectively mitigates the negative impacts of water scarcity by increasing parameters like RWC, SOD, and CAT activities, thus preventing oxidative damage. Given the limited existing literature on the joint use of TiO2NPs and AsA via foliar application in coffee plants under drought stress and their protective role against oxidative damage, we propose that employing TiO2NPs and AsA should be considered as a sustainable alternative strategy to enhance coffee production in drought conditions. Our findings could shed light on the beneficial impact of TiO2NPs and AsA not only in coffee plants but also in other significant field crops. Deeper investigations into the molecular mechanisms by which TiO2NPs and AsA improve drought tolerance in coffee plants are needed. They could involve gene expression analysis to comprehend how these treatments affect stress-related signaling pathways. Upcoming investigations should concentrate on the long-term effects of TiO2NPs and AsA application on coffee as well as other agronomically important plants.
