. Introduction
For annual crop species, the loss of seed dormancy is among the first signs of domestication. This trait is actively selected against by cultivation itself: lineages that tend to lay dormant will not contribute to the following harvest and thus will not be re-sown the following year (Fuller & Allaby, 2018; Hammer, 1984; Spengler, 2020). The breaking of seed dormancy is still one of the major tools to facilitate breeding programs for releasing new germplasms (Abbasi, 2020; Yalamalle et al., 2024), utilizing or domesticating wild species (Andriamparany & Buerkert, 2019; Khan et al., 2023; Singh et al., 2020; Zhao et al., 2024), to extinguish heterogeneous germination in medicinal plants (Ekren et al., 2024), and even for the preservation of uncommon species (Cuena Lombraña et al., 2024; Kırmızı, 2023; Kołodziejek, 2018; Krasylenko et al., 2023; Santiago et al., 2023).
Stachys annua (L.) L., a species from the Lamiaceae family, is native to the Anatolian-Armenian region, from where it was introduced in Europe as an archaeophyte plant (Csiky et al., 2023), and turned out to be a characteristic weed species of stubble fields (Skrajna et al., 2018; Pinke et al., 2021). In Central Europe, the development cycle of S. annua begins in late spring, in the lowest vegetation layer; however, it only flowers due to the larger light input after cereal stands are harvested (Pinke & Pál, 2009). Its most popular Hungarian name, ‘tarlóvirág’, which literally means ‘stubble-flower’, also hints to its most typical flowering site (Berkó & Kardos, 20231937; Juhász, 20231983; Pinke et al., 2021).
About one and a half centuries ago, S. annua was the most important honey plant in the Carpathian Basin. In favourable conditions at the peak of flowering, rich stands of S. annua were documented to yield up to 8 kg of honey per bee colony per day, and even seasonal average yields could reach 2–3 kg/day/colony (Berkó & Kardos, 1937; Juhász, 1983; Pinke et al., 2021). The nectar production by S. annua can however highly depend on weather conditions (Pinke et al., 2021). As a good quality product, its ample and characteristic monofloral honey used to be a “world brand” of Hungarian apiculture (Pinke et al., 2021). Facing an initial decline in its spontaneous populations, in the early 20th century, there were several well-documented efforts for “saving” this highly esteemed plant by taking it into cultivation (Pinke et al., 2021). Then, in the 20th century, the continued agricultural intensification resulted in a drastic population collapse, turning S. annua into a floristic rarity (Pinke, 2020; Pinke et al., 2021). The early attempts for cultivation were greatly hindered by the low germination rates of harvested seeds, which tended to lay dormant in the soil for several years (Pinke et al., 2021). For efficient cultivation, this dormancy must be eliminated through the invention of applicable seed treatments that can break it.
Although similar seed germination behaviours have already been reported for several, mainly biennial or perennial species of the genus Stachys, including S. alpina (Pinfield et al., 1972, 1975), S. byzantina (Rezvani et al., 2021), S. germanica (Dunn, 1997; Güleryüz et al., 2011), S. mouretii (Ismaili et al., 2023), S. palustris (Brändel, 2006), and S. sylvatica (Graae et al., 2009; Slade & Causton, 1979), the seed dormancy release and germination requirements of S. annua have not been studied yet.
Pinfield et al. (1972) achieved 90–95% germination proportion of S. alpina L. when seeds were treated with 104 ppm GA and kept in constant darkness at 20°C for 12 days. It was observed that higher concentrations of GA increased the germination rate but did not affect the final germination. However, reduced concentrations of GA resulted in lower germination percentage. No germination was observed in distilled water treatments or after stratification for less than 4 weeks. Their studies showed that storage in wet sand for 12–20 weeks was necessary to produce low germination (20–30%). Thus, in S. alpina, almost complete germination could be achieved with GA treatment, whereas stratification was only partially effective (Pinfield et al., 1972). This is due to the relatively slow increase in endogenous gibberellic acid levels in freshly ripened, dormant seeds of S. alpina during post-ripening (wet chilling), which can be accelerated by the application of external GA (Pinfield et al., 1975). Slade & Causton (1979) investigated the effects of five dormancy-breaking treatments – potassium nitrate, scarification, ethylene, light, and stratification – on S. sylvatica L. seeds in a full factorial experiment. It was found that germination occurred in about two-thirds of the different factor combinations, but germination rates were generally low. The best germination (70%) was achieved by stratification at 5°C for 6 weeks, with scarification being inhibited. Thompson & Grime (1979) recorded that seeds of S. sylvatica stored dry at 5°C required a large diurnal temperature fluctuation (9°C) to achieve the maximum germination %. Graae et al. (2009) found that fresh seeds of S. sylvatica, collected from a range of sites across central and northwestern Europe, had a very low germination percentage (0–19%). After 18 weeks of cold stratification followed by 6 weeks of warm incubation (14 h light/20°C and 10 h dark/10°C), germination ranged from 13% to 54%. When seeds were exposed to 6 weeks of warm incubation followed by 18 weeks of cold incubation and finally 6 weeks of warm incubation, germination was increased to 50–91%. Brändel (2006) observed that germination of S. palustris L. seeds took place between 12–36°C. Optimal germination was recorded at constant temperatures of 24–33°C if the seeds had been stratified (10 weeks at 5°C); however, germination increased under increasing diurnal temperature fluctuation (4°C) up to a maximum of 80% at a 14°C amplitude, whether the seeds were stratified or not. A period of cold stratification reduced primary dormancy, and germination was always lower (<25%) in the dark (constant), irrespective of temperature fluctuation regimes (Brändel, 2006). After-ripening effects on germination rates of seeds of S. palustris were not detected, and germination rates of seeds collected in autumn and stored at room temperature for 7 months did not increase (Brändel, 2006). Investigations on S. germanica L. seeds (Güleryüz et al., 2011) showed that treatments with GA or a combination of GA and KIN were successful in terminating seed dormancy. Seeds treated with a combination of 250 ppm gibberellic acid (GA) + 50 ppm kinetin (KIN) after 30 days of stratification (wet chilling + 4°C) achieved the highest germination rate (95%) under continuous darkness and photoperiodic (12 h dark/20°C and 12 h light/10°C) conditions. However, without stratification in cold wet sand, only slightly more than a third of seeds treated with GA germinated. In another experiment without pre-treatment for 26 days, S. germanica seeds germinated neither in light nor in darkness at temperatures between 5 and 26°C. Even at higher temperatures (29–35°C), a low rate of germination was observed (Dunn, 1997). However, seed germination of S. byzantina K. Koch was not markedly stimulated by GA concentrations in both light/dark and complete darkness conditions (Rezvani et al., 2021). In laboratory experiments, Ismaili et al. (2023) showed that alternating temperatures strongly affected the seed germination capacity and speed in the case of S. mouretii Batt. & Pit. The temperature regimes of 25/10°C and 35/20°C significantly increased the germination percentage to 66.5% and 65.5%, respectively. Furthermore, the most effective pre-treatment improving germination was GA at an optimal concentration of 500 mg/l by improving the germination percentage to 91.5% and reducing the median time of germination (T50) and mean germination time (MGT) to 11.7 days and 9.9 days, respectively.
The timing of germination is a critical circumstance in a plant’s life cycle (Donohue, 2005), which is determined by the seed dormancy mechanisms/behaviour (Postma & Ågren, 2015). From an ecological perspective, seed dormancy can be defined as the prevention of germination even when suitable conditions are prevalent (Benech-Arnold et al., 2000). The dormancy mechanism allows a species to synchronise its germination with favourable environmental conditions, which increases its probability of survival and establishment (Baskin & Baskin, 2014). In nature, based on its timing, two principal categories of seed dormancy are distinguished. Primary dormancy is an innate seed characteristic that establishes during seed maturation on the mother plant, while secondary dormancy occurs after shedding from the parent plant in non-dormant seeds that were exposed to abiotic stresses associated with prolonged lack of external environmental conditions required for germination (e.g. temperature, light, soil moisture, oxygen, nitrate) (Buijs, 2020). The causes of primary seed dormancy are complex; it may be developed in the embryo or imposed by the tissues that surround the embryo. In several species, both the embryo and the tissues enclosing it impose dormancy (Hilhorst et al., 2010). In temperate regions, the seeds of most plant families exhibit physiological dormancy, which is caused by insufficient embryo growth potential (Baskin & Baskin, 2014). Depending on its depth, several techniques can be used to overcome seed dormancy. These techniques include, for example, treatments with GA, potassium nitrate (KNO3), cold and warm stratification, physical and chemical scarification, and leaching (Baskin & Baskin, 2014). The stratification treatments mimic the slow seasonal changes that modify seed dormancy levels gradually over time (mean temperature, seed moisture), while GA addition as well as light and temperature fluctuations are signals that may indicate a “spatial window” of opportunity for germination, with a sudden termination of seed dormancy (Benech-Arnold et al., 2000; Brändel, 2006; Ismaili et al., 2023).
We hypothesized that the combination of stratification and gibberellic acid would release the dormancy of S. annua seeds. To explore this hypothesis, we addressed the following main aims: (I) exploration of the effect of different moist stratification regimes for 4 weeks (in cold/warm sand) on breaking dormancy and promoting germination, and (II) determination of the effect of gibberellic acid on seed germination under two light-temperature regimes (constant/fluctuating) for S. annua. Controlled laboratory experiments were conducted to achieve these objectives, preceded by stratification pre-treatments simulating natural environmental conditions. Identifying effective methods for breaking the dormancy of S. annua seeds in laboratory conditions may help to achieve higher and more uniform germination rates. The availability of such techniques could be an important first step towards introducing S. annua into cultivation.
. Materials and methods
. Collection and storage of seeds
The seeds of S. annua were manually harvested from natural matured plants in a population growing in a stubble of an extensively managed phacelia (Phacelia tanacetifolia Benth.) field in the Lesser Plain of north-western Hungary (47°84'97"N; 17°09'47"E). This specific phacelia field was cultivated for seed production, which is quite common in the region (Dunai & Pinke, 2023), and it was left uncultivated after harvest to preserve its spontaneously emerging S. annua population that was already known to local beekeepers (Pinke et al., 2020). The gathering of S. annua seeds took place in the middle of September 2020. The seeds were cleaned and stored dry in paper bags in ambient conditions (20±5°C) in the laboratory until the start of the experiments, which began on 10 November 2020.
. Germination experiments
. Dormancy breaking methods
We analyzed the impact of three possible treatments on the germination of S. annua in a full factorial design. The first one was a stratification (STR) treatment implemented at different levels: no stratification (NS: using fresh seeds as the control), cold stratification (CS: 5°C for 4 weeks), and warm stratification (WS: 21°C for 4 weeks). The second treatment involved the application of gibberellic acid (GA) with two possible levels: no GA (DW: distilled water as the control) and gibberellic acid (GA: 250 mg/l solution of GA). The third treatment was an environmental light-temperature regime (LTR) treatment, with two possible levels: a “constant regime” (CR: continuous darkness at a stable 20°C temperature as the control) and a “fluctuating regime” (FR: 14 h at 20°C & 300 μmol photons m-2 s-1 fluorescent light, followed by 10 h at 10°C in darkness to simulate a germination-friendly diurnal light-temperature pattern).
. Germination test protocol
We performed five replications for treatment combinations with no stratification (NS) and four replications for combinations involving stratified seeds (CS, WS). The stratification treatment was implemented first: the seeds selected for CS or WS were wrapped in fine nylon mesh, placed in moist sand-filled containers, and exposed to the selected temperature for four weeks. The germination trials were performed after the (optional) stratification in incubation chambers. At the beginning of each trial, seeds were tested for viability using a 1% solution of 2,3,5-triphenyl-tetrazolium chloride (TTC) with 50 seeds in four replications (Peters, 2000). Selected S. annua seeds were washed under running tap water, disinfected with a 50% ethanol solution for 4 minutes, and then rinsed 5–6 times with sterile deionized water. After rinsing, the seeds were not dried but immediately placed in incubation chambers. In each trial, 50 seeds were placed on two layers of filter paper (MN 640 W) in a Petri dish (with 9 cm diameter) and were moistened with 6 ml of the selected solution (DW or GA). The Petri dishes were sealed with two layers of para-film to prevent moisture loss and, in the constant dark regime (CR), they were also covered with two layers of aluminium foil to avoid light penetration. The number of germinated seeds was recorded 2, 7, and 15 days after starting the germination test, and the accumulated germination percentage (AGP) after the 15 days was used as the response variable. Seeds were considered to be germinated when they produced a visible radicle longer than 1 mm (ISTA, 1985).
. Statistical analysis
To account for the binary nature of germination data, we applied a logistic regression model with the logit link function for the data analysis. To evaluate the effects and interactions of the experimental factors, a series of nested models were fitted and compared using Likelihood Ratio Tests (LRT) to assess the significance of main effects (STR, GA, LTR) and their interactions (STR × GA, STR × LTR, GA × LTR, and finally STR × GA × LTR). Results were considered statistically significant at a 95% confidence interval (p < 0.05 and p < 0.001). The entire analysis was performed in R (version 4.3.2.) (R Core Team, 2024).
. Results
. General traits of seed dormancy release and germination
According to the tetrazolium test, 90% of the seeds of S. annua were viable. The accumulated germination percentage (AGP) in the control trials (i.e. NS, DW, CR) was 2.4%, indicating that the vast majority of freshly matured (NS) seeds were in the state of primary dormancy (Figure 1).
Figure 1
Effects of stratification, gibberellic acid, and light-temperature regimes on dormancy release and germination of Stachys annua seeds. Note: DW: distilled water; GA: 250 mg/l solution of GA; CR: constant regime (20°C in continuous darkness); FR: fluctuating regime (simulated diurnal light and temperature changes).
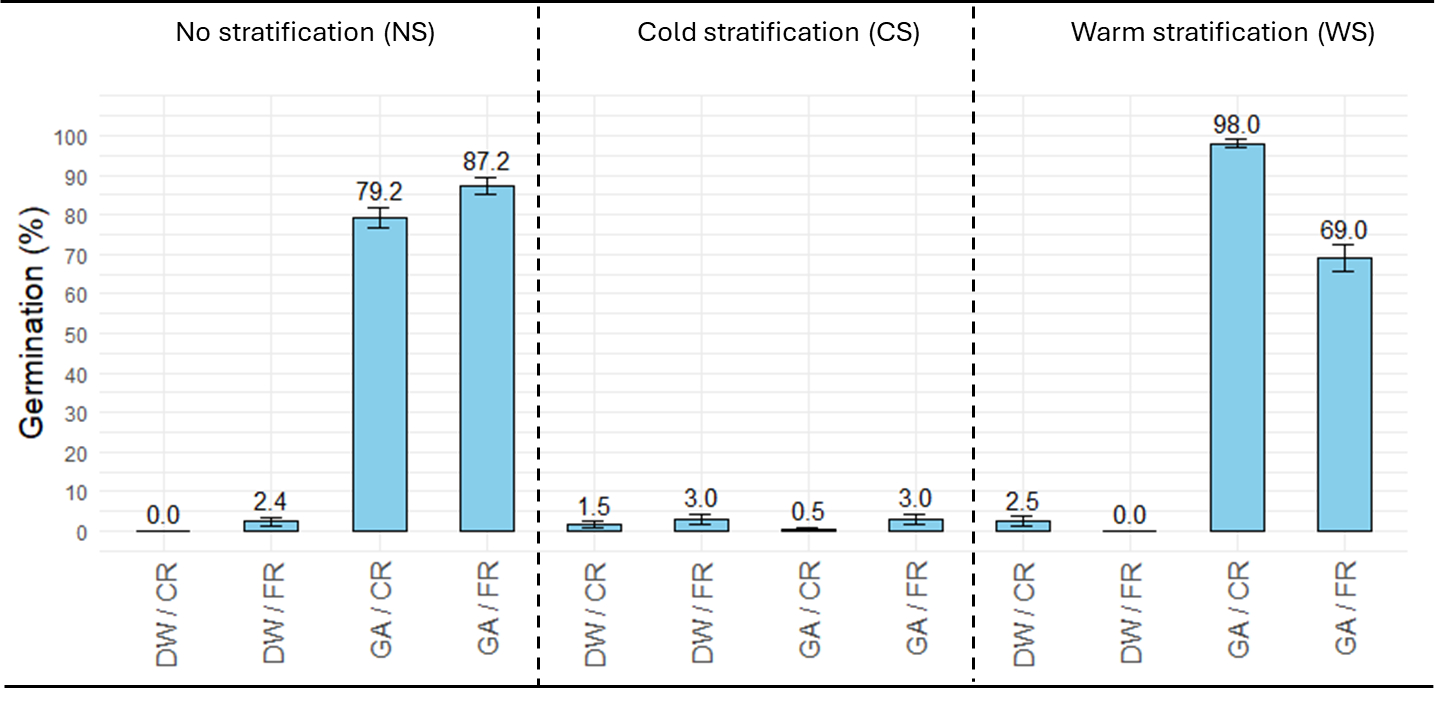
. Effect of stratification on seed dormancy release and germination
The results indicate that stratification (STR) had a strongly significant effect on the germination of S. annua seeds (Table 1). Nevertheless, despite this significant overall impact, the effect of stratification (STR) was clearly different in the various treatment combinations. With the control levels of the two other treatments, neither cold (CS) nor warm stratification (WS) had a clear effect (AGP = 0 – 3%) (Figure 1). However, warm stratification (WS) combined with gibberellic acid (GA) significantly increased the germination percentage, whereas cold stratification (CS) with GA showed no similar effect (Figure 1).
Table 1
Results of the LRT test
. Effect of gibberellic acid on seed dormancy release and germination
Gibberellic acid (GA) also had a highly significant overall influence on the seed germination (Table 1). The presence of GA significantly increased the germination of non-stratified (NS) and warm stratified (WS) seeds under both light-temperature regimes (LTR). In fact, these trials showed an enormous impact of gibberellic acid (GA), changing germination rates from close to zero to very high (70–100%), with an almost unbeatable 98% rate in one of the treatment combinations (WS/GA/CR, Figure 1). In contrast, cold stratified seeds (CS) that had been stored in wet sand at low temperature for 4 weeks (cold stratification) failed to germinate even with GA treatment (AGP = 0.5 and 3%). These results suggest that cold stratification (CS) has increased the seed dormancy level compared to freshly matured seed lots (NS) treated with the GA solution (Figure 1).
. Effect of light-temperature regimes on seed dormancy release and germination
From the three factors studied, light-temperature regimes (LTR) seemed to exert the weakest impact on the germination process. Nevertheless, the interaction of stratification (STR) with the environmental regime (STR × LTR) was found clearly significant (Table 1). This is probably the consequence of LTR having the opposite effect for warm stratified seeds (WS) than for the other two types of stratification. While for no stratification (NS) and cold stratification (CS) the fluctuating regimes (FR) resulted in slightly higher germination, after warm stratification (WS) the constant regime (CR) seemed to provide a better germination support, including the close to unbeatable 98% results (Figure 1).
. Discussion
It is well established from the relevant literature that freshly harvested, mature seeds of members of the Stachys genus are generally in primary dormancy (Dunn, 1997; Godefroid et al., 2010; Graae et al., 2009; Güleryüz et al., 2011; Pinfield et al., 1972, 1975). The results of this study demonstrate that most freshly harvested mature seeds of S. annua were dormant at maturity. Seeds failed to germinate in response to short-time (4 weeks) cold or warm stratification treatments with distilled water in both constant and fluctuating light-temperature conditions. A similar phenomenon was also observed in the study on seeds of S. germanica (Güleryüz et al., 2011). According to Pinfield et al. (1972), S. alpina seeds did not germinate after less than 4 weeks of cold chilling, and only a limited increase in germination (20–30%) occurred even after periods as long as 12–20 weeks (Pinfield et al., 1975). The study of Graae et al. (2009) indicated that fresh seeds of S. sylvatica, gathered in various habitats across central and north-western Europe, had very poor germination (0–19%). After 18 weeks of cold stratification followed by 6 weeks of warm incubation (14 h light at 20°C and 10 h dark at 10°C), germination fluctuated between 13% and 54%. When seeds were subjected first to 6 weeks of warm incubation followed by 18 weeks of cold incubation and finally 6 weeks of warm incubation, germination was raised to 50–91%. This means that in both natural habitats and agroecosystems, the dormancy of S. annua seeds presumably takes extended periods of time to disappear. This is confirmed by its prolonged field emergence, which usually takes place only in the second spring after the maturation (Dunai et al., 2020).
Based on our results, it appears that the seeds of S. annua are in non-deep physiological dormancy, with gibberellic acid playing a substantial role in terminating it. The application of GA alone considerably improves the germination of freshly harvested mature S. annua seeds. Similar results were found by Pinfield et al. (1972) for S. alpina, where maximum germination (> 90%) was achieved at 104 mg/l of GA. Ismaili et al. (Ismaili et al., 2023) reported that the most efficient pre-treatment to boost seed germination in S. mouretii was GA at an optimum concentration of 500 mg/l, increasing the germination rate up to 91.5%. Nevertheless, the environmental factors required for gradually overcoming seed dormancy act by increasing the active level of endogenous GA, so externally applied GA often substitutes for certain environmental stimuli such as specific temperature pre-treatment (stratification) or light (Karam & Al-Salem, 2001; Karssen & Laçka, 1986). This was confirmed by the results of our experiment with freshly collected mature S. annua seeds. At the same time, in addition to the substitution effect, the simultaneous combination of GA with other seed dormancy release methods, such as cold stratification (Balouchi & Sanavy, 2006) or other chemicals, such as potassium nitrate (KNO3) (Mousavi et al., 2019), and plant hormones, such as cytokinin (Ganjali et al., 2022), can have a synergistic effect in terminating the dormancy of the treated seed. Our results suggest that the highest germination rate (98%) can be reached in the seeds handled with a combination of 250 mg/l GA and 4 weeks of warm stratification, when germination takes place in the dark for 24 hours at 20°C. Comparing these results with laboratory studies with other species of the genus Stachys, it seems that the germination biology of S. annua exhibits strong similarities with the germination biology of S. alpina. To obtain this among natural circumstances, longer periods of seeds remaining on the mother plant and lying in the soil are needed.
At the same time, our findings do not exclude the probability that, beside the high ABA/GA ratio in tissues during seed maturation, the dormancy of S. annua seeds may also be influenced by inhibitors in the embryo and/or by the immaturity of the embryo itself. However, to clarify this, further investigations are required. In addition, it should also be noted that the type of pollination may also influence the different germination responses of various populations of Stachys species. Wilcock and Jones (1974) observed that mature seeds from plants collected from cross-pollinated stands of S. palustris showed nearly twice the germination rate (90%) of their peers collected from self-pollinating plants (52%). We have not performed such analyses on S. annua seeds, but considering that the site of the seed collection was frequently visited both by honey bees and wild pollinators (Pinke et al., 2020), the high germination rates of our seed samples suggest that they originated mainly from cross-pollinated plants. Hegi (1975) also documented that S. annua is primarily an insect-pollinated species, though self-pollination can occasionally also occur.
In our experimental design, we analyzed the effect of light and temperature regimes jointly as a combined treatment. This combined analysis was partly inspired by nature (where light and temperature fluctuations also occur somewhat jointly: deeply buried seeds receive neither, while seeds on the surface experience both), and partly necessitated by the availability of technical equipment in our laboratory (a limited number of incubators). Of course, this way we cannot know where the effects of “LTR” (our combined light-temperature regime treatment) come from: are these effects primarily caused by the changing temperature, or should they rather be attributed to the presence of (or changes in) light? Answering this question would demand a follow-up experiment. Since LTR had only minor impacts on the germination rate and (in particular) the most promising treatment combination involved a stable environmental regime, this research direction might not be urgent from a practical perspective.
Nevertheless, several further challenges still need to be solved before S. annua can become a successful crop. This list includes several complex technological and even social challenges ranging from motivating farmers for leaving their stubbles unploughed to developing efficient techniques for the laborious steps of seed harvesting (Karssen & Laçka, 1986; Pinke et al., 2021). In any case, our results are very encouraging for the development of a dormancy-breaking technology and its large-scale application on the seeds of S. annua to achieve a uniform germination which will enable its future growing as a valuable bee pasture.
. Conclusions
Our results confirmed our hypothesis that the combination of stratification and gibberellic acid would release the dormancy of S. annua seeds. Our findings proved that gibberellic acid plays an important role in breaking the seed dormancy of S. annua and can provide a replacement for the stratification requirement of seeds of this species for germination, which can be used to produce uniformly germinating seeds for commercial purposes. The seed dormancy-breaking treatments should be performed in strictly controlled conditions by seed companies. The cost of GA should be incorporated into the price of the seeds, ensuring that the final cost remains manageable for beekeepers and farmers.
