. Introduction
Purple moor grass meadows (Molinion caeruleae) represent semi-natural plant communities that have developed in seasonally wet habitats. They occur on mineral, organic, and muck soils, often poor in nutrients with acidic, neutral, or alkaline pH (Mencel et al., 2024; Zelnik & Čarni, 2008). Molinion caeruleae meadows are characterized by great diversity, which results from geographical and edaphic variability, as well as from different methods and intensity of management. Their formation and maintenance is associated with extensive human management. Compared to other natural habitats, purple moor grass meadows are distinguished by high species richness and the presence of many endangered species (e.g. Dianthus superbus, Gentiana pneumonanthe, Gladiolus imbricatus, Iris sibirica, and Trollius europaeus), which makes them biodiversity hotspots (Havlová, 2006; Řezníčková, 2007; Ziaja & Wójcik, 2016). Most of these meadow patches were excluded from use for economic reasons, which led to the initiation of the secondary succession process and the transformation of these communities into herbaceous vegetation, thickets, or forests and into sedge reed beds in wet places. They are also threatened by changes in water conditions and intensification of management (Kulik, 2014; Michalska-Hejduk & Kopeć, 2012; Ziaja et al., 2017). Meadows from the Molinion caeruleae alliance (6410) are currently rare and endangered natural habitats (Perzanowska & Korzeniak, 2020), which is why they are protected throughout the European Union (Council Directive, 1992). According to the current state of knowledge, the proper way to protect purple moor grass meadows is late mowing in late August or early September combined with biomass collection. Mowing should take place once a year or every two years. Less frequent mowing may stop succession but is not able to maintain high biodiversity. On the other hand, more intensive use leads to the disappearance of characteristic species of the Molinion caeruleae alliance and a decrease in species diversity (Havlová, 2006; Swacha et al., 2018; Zelnik, 2011). It has been evidenced that grazing is not the best way to protect Molinion caeruleae meadows, because it selectively affects different plant species. Although grazing can be a way to maintain this habitat, after a few years it leads to changes in the species composition (Kulik, 2014; Sienkiewicz-Paderewska et al., 2020). Burning is also not an effective way to protect Molinion caeruleae meadows. Although it stops the accumulation of litter and the development of succession, it leads to a decrease in species richness and disappearance of some characteristic species (Bódis et al., 2021; Wójcik et al., 2022). So far, the effect of accidental or prescribed burning on regeneration of particular species occurring in Molinion caeruleae meadows has focused on seedling recruitment and frequency in plant cover. The weak positive effect of fire on seedling recruitment was evidenced inter alia in the case of Gentiana pneumonanthe (Křenová & Lepš, 1996) and Gladiolus imbricatus (Jõgar & Moora, 2008). Other authors reported that Achillea millefolium (Halpern et al., 2019), Antoxanthum odoratum (Milberg et al., 2018), Briza media (Milberg et al., 2018), Ranunculus acris (Milberg et al., 2018), and Serratula tinctoria (Jefferson & Walker, 2017) are much less abundant in recently burnt stands, in contrast to Cirsium arvense (Milberg et al., 2018), Deschampsia caespitosa (Nuckols et al., 2011), Galium verum (Milberg et al., 2018), Leucanthemum vulgare (Milberg et al., 2018), and Molinia caerulea (Brys et al., 2005; Jacquemyn et al., 2005; Taylor et al., 2001). Moreover, in Molinia caerulea populations, a fast spread of seedlings and a significant increase in the aboveground biomass of individuals growing in post-fire stands were evidenced (Brys et al., 2005; Jacquemyn et al., 2005). Also, Pyke et al. (2010) found that cryptophytes and hemicryptophytes are able to survive fires due to protection of renewal buds. However, as suggested by the aforementioned authors, seed production by resprouters may be limited initially after a fire, especially in species that require a year of growth to develop flowers. Similarly, Clark and Wilson (2001) found that burning significantly decreases inflorescence production in Deschampsia caespitosa. In contrast, greater post-fire production of inflorescences was evidenced in populations of Molinia caerulea (Brys et al., 2005) and Briza media (Lloyd, 1968).
Inula salicina L. is a species occurring in Molinia meadows, which is prone to progressive diminishing of population numbers. In recent decades, Inula salicina has been included in several national red lists and books with the status near threatened NT (Bornand et al., 2016; Colling, 2005; Grulich, 2017; Sparrius et al., 2014), endangered EN (Curtis & McGough, 1988), or critically endangered CR (Wyse Jackson et al., 2016). It is also included in the red lists and books of many regions of e.g. France (The National Inventory of Natural Heritage), Germany (Metzing et al., 2018), and Poland (Jackowiak et al., 2007; Pliszko, 2017; Podgórska, 2014). The greatest threat to populations of the aforementioned species is connected with the abandonment of traditional land use in semi-natural habitats leading to secondary succession. Considering this, numerous authors have conducted observations of the regeneration of its populations, particularly in meadows representing Deschampsion (=Cnidion) and Molinon alliances.
The aforementioned studies confirmed that spontaneous seedling recruitment of Inula salicina without seed addition in artificially made openings appeared as an effect of manual removal of litter, above-ground plant cover, and topsoil (e.g. Kostrakiewicz, 2011; Kostrakiewicz-Gierałt, 2013). Other authors’ observations focused on the influence of mowing (Bissels et al., 2006), topsoil removal (Hölzel & Otte, 2003), or topsoil rotovating (Donath et al., 2007; Harvolk-Schöning et al., 2020), followed by diaspore transfer, on the appearance of Inula salicina individuals. Nevertheless, despite the growing scientific interest in the effects of disturbances in abandoned meadows on the regeneration of populations of Inula salicina, the current state of knowledge is still insufficient. Particularly, considering the lack of investigations of the impact of fire, the presented studies aiming to assess the effect of accidental burning on the regeneration of populations of the aforementioned species in Molinia meadows were undertaken.
We hypothesized that: (I) the number of seedlings is greater while the number of vegetative and generative stems of Inula salicina would be lower in the burned patch than in the unburned patch; (II) the traits of Inula salicina generative stems (height, number and dimensions of leaves, number of inflorescences, diameter of the receptacle with disc flowers of the largest inflorescence) would achieve lower values in the burned patch than in the unburned patch; (III) the numbers of seedlings and stems, as well as the individual traits of the generative stems of Inula salicina would not show temporal variability within both study patches.
Considering the presented hypotheses, the following specific research aims were set: (I) to study the number of seedlings, as well as the number of generative and vegetative stems on the burned and unburned patches in particular study years; (II) to investigate the individual traits of generative stems on the burned and unburned patches in particular study years.
. Material and methods
. Study species
Inula salicina L. (Mirek et al., 2020) is a perennial clonal herb producing hypogeogenous stems (rhizomes) with long internodes, enabling fast vegetative spread (Klimešová et al., 2017). Its thin above-ground stems (20–80 cm in height) and narrow, elongate, alternate, sessile leaves are roughly haired. The inflorescences (flower heads) 2.5–4.0 cm in diameter are collected in loose umbel-shaped synflorescences or set singly at the top of the stem. Each head contains 100–250 yellow disc flowers located in a receptacle and 35–70 yellow ray flowers (WFO, 2025).
The Inula salicina species is neutral to continentalism and occurs in moderately warm localities with moderate light intensity in lowland areas and lower mountainous sites (Chytrý et al., 2018; Zarzycki et al., 2002). In edaphic terms, it has a wide range of tolerance, as it occurs in fresh habitats, as well as moist, periodically moist, and even dry sites. It grows on neutral or alkaline soils which are often rich in calcium carbonate. These habitats have varied contents of nutrients (from poor to moderately rich habitats) (Bissels et al., 2004; Chytrý et al., 2018; Donath et al., 2007; Stokłosa et al., 2016; Zarzycki et al., 2002). Inula salicina can occur both on mineral substrates and on muck soils. It most often grows on alluvial soils with a varied granulometric composition in waterlogged areas characterized by large fluctuations in water levels (Bissels et al., 2004; Donath et al., 2007; Goncharenko et al., 2020; Kącki, 2007; Šilc et al., 2014; Stokłosa et al., 2016; Załuski, 1995).
The species shows a wide range of phytocoenotic tolerance. It is most often recorded in wet Molinion caeruleae meadows (Kącki, 2007; Priede, 2011; Šilc et al., 2014; Swacha et al., 2016). It often grows in alluvial meadows from the Deschampsion caespitosae alliance (Bissels et al., 2004; Goncharenko et al., 2020; Załuski, 1995). Additionally, the species was recorded in moist and wet localities, i.e. transitional mires of the Scheuchzerio-Caricetea fuscae class (Zlinská, 1994). On the other hand, it has also been found in much drier habitats, e.g. fringe vegetation of the Trifolio-Geranietea class (Klinkovská & Roleček, 2024) and dry grasslands of the Festuco-Brometea class (Škodová et al., 2015). Interestingly, this species may also occur in forest communities with substantially thinned tree stands (Bjørndalen, 1985). According to the EUNIS habitat classification (2022), Inula salicina is considered a characteristic species of Atlantic and sub-Atlantic humid meadows (E3.41), Purple moorgrass meadows and related communities (E3.51), and Calcicline purple moorgrass meadows (E3.511).
. Study area
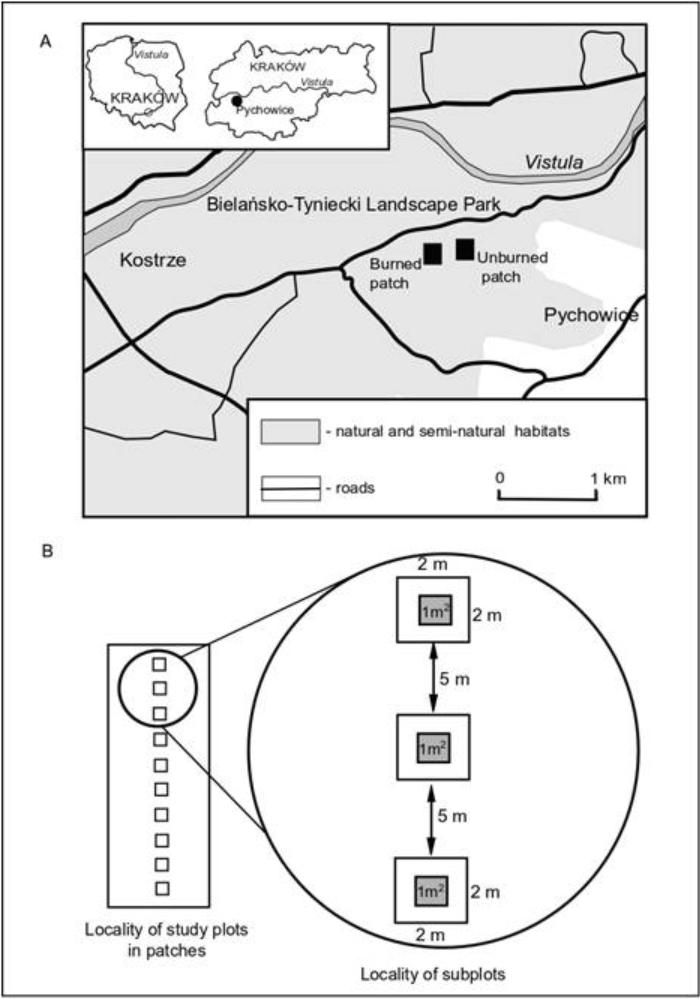
The field observations were carried out in the south-western part of Kraków (southern Poland) in Molinion caeruleae patches, i.e. remains of meadows that are widely scattered along the Vistula valley and date from the beginning of the 20th century (Dubiel, 1991, 1995; Zarzycki, 1958). Transformations of these meadows induced by intensive economic activities were observed as early as in the first half of the 20th century. As reported by Zarzycki (1958), some Molinion caeruleae patches were dissected by drainage ditches, which led to their desiccation. The aforementioned meadows have been abandoned since the late 1980s. The cessation of mowing has resulted in the progressive process of secondary succession by high-growing macrophores, expansive grasses, shrubs, and trees, which can be observed in areas that have not been used for a long time (Dubiel, 1996). The progressive transformation of vegetation was also reported by Wójcik and Janicka (2016), who found a significant increase in the coverage of herbal species from the Filipendulion ulmariae alliance (Lythrum salicaria, Filipendula ulmaria, Geranium palustre) resulting from long-term abandonment. This led to the fragmentation of initially large areas of Molinion caeruleae meadows and a reduction in their surface area. Reed communities dominated by Phragmites australis were found to develop in highly humid habitats located in the land depressions present in the area. In turn, drier sites were colonized by the expansive grass Calamagrostis epigejos and the invasive species Solidago gigantea and S. canadensis. Simultaneously, the researchers reported that, in the spring of 2015, some part of the meadow was accidentally burned (Figure 1), while the other part remained intact (Figure 2).


. Field studies
The research plots were designated in homogeneous patches based on previous studies (Wójcik & Janicka, 2016) and field observations conducted by the authors in the period preceding the accidental burning. The burned and unburned plots were characterized by a similar floristic composition before the fire in 2015. In the burned and unburned plots, a total of 57 species of vascular plants occurred in 2012–2014. In both plots, the most numerous species represented the Molinion caeruleae alliance, with the greatest cover achieved by Molinia caerulea, Galium boreale, and Betonica officinalis. Cirsium rivulare from the Calthion palustris alliance and Geranium palustre from the Filipendulion ulmariae alliance also had high cover, as well as species characteristic of the Molinietalia order (Sanguisorba officinalis, Serratula tinctoria, Selinum carvifolia). Moreover, Phragmites australis and Carex gracilis from the Phragmitetea class achieved significant coverage in both plots. The pilot observations were conducted in July 2016, while the investigations were started in July 2017 when the plant cover recovered after the disturbance. In the accidentally burned (50.030393°N, 19.867092°E) and unburned (50.030882°N, 19.868433°E) patches, two line transects, each with 10 permanent plots measuring 4 m2, were established at 5 m intervals (Figure 3). The detailed characteristics of plant cover and soil properties investigated in particular plots after burning were presented by Wójcik et al. (2022). In the central part of each plot, one permanent subplot was established. The terms “individual” and “stem” were adopted as the main demographic units. The term individual was adopted in the case of seedlings (characterized by the presence of cotyledons or their remnants), because only at this stage could it be established for sure that they had developed from the zygote. In the case of later stages, the term “stem” was applied due to difficulties in distinguishing particular individuals without digging up their underground organs. Within the aforementioned subplot, observations of the number of seedlings as well as the number of vegetative and generative stems of Inula salicina were conducted over a period of 3 years (2017–2019). Moreover, selected traits of all generative stems were studied in each subplot. In subplots where they occurred abundantly (at least 21 stems), the number of examined stems was limited to 20. The stems were randomly selected based on 20 throws with an iron rim, 20 cm in diameter. Each time, one stem occurring in the centre of the rim was measured and labeled. The following traits were investigated: (I) height of generative stems from the ground level to the base of the top inflorescence, (II) number of leaves, (III) length of the longest leaf, (IV) width of the longest leaf taken at ½ of its length, (V) number of inflorescences, (VI) diameter of the receptacle with disc flowers of the largest inflorescence.
. Data analysis
Vascular plants were listed according to Mirek et al. (2020), while the nomenclature of syntaxa was adopted according to the EuroVegChecklist (Mucina et al., 2016). Statistical analysis was performed using nonparametric tests. The U Mann-Whitney test was applied to check the statistical significance of differences in (I) number of generative stems, (II) number of vegetative stems, (III) height of generative stems, (IV) number of leaves in generative stems, (V) length of the longest leaf in generative stems, (VI) width of the longest leaf in generative stems, (VII) number of inflorescences, and (VIII) diameter of the receptacle of the greatest inflorescence between the burned and unburned study patches in particular years. The value of U statistics was reported for comparisons of groups not exceeding 20 data, but the standardized value Z was given when at least one group was more numerous. The statistical significance of differences of the aforementioned traits between particular study seasons was tested using the Friedman rank test.
. Spatial variability of the number and traits of Inula salicina stems
The presented studies showing a greater number of vegetative and generative stems in the burned patch, compared to the unburned patch, during the whole study period do not support the first working hypothesis. In 2017–2019, the number of vegetative and generative stems per subplot was significantly greater in the burned patch than in the unburned one (Table 1). Altogether, in the unburned patch 36, 25, and 9 generative stems were investigated in the subsequent study years, whereas the number of generative stems in the burned patch reached 166, 179, and 182 respectively. Moreover, during the whole study period, no seedlings were noted at either study site. Therefore, our first hypothesis must be fully rejected.
The individual traits were investigated in generative stems occurring in the unburned patch and the investigations performed did not support the second hypothesis that the height of generative stems, the number and dimensions of leaves, the number of inflorescences, and the diameter of the receptacle of the largest inflorescence of Inula salicina would achieve lower values in the burned patch than in the unburned site. The height of generative stems (Table 2), the length of the longest leaf (Table 3), and the number of inflorescences per flowering stem (Table 4) did not differ between the patches in 2017. They were greater in the burned patch in 2018 and in the unburned patch in 2019. The number of leaves per generative stem (Table 2) was significantly greater in the burned patch only in 2018, the diameter of the receptacle of the largest inflorescence (Table 4) was noticeably greater in the burned patch only in 2017, whereas the width of the longest leaf (Table 3) was similar at both study sites.
Table 1
Mean (±SD) number of vegetative and generative stems of Inula salicina recorded in 10 subplots in unburned and burned patches of Molinion caeruleae meadow in 2017–2019.
Table 2
Mean (±SD) height (cm) of generative stems of Inula salicina and number of leaves per subplot in unburned and burned patches of Molinion caeruleae meadow in 2017–2019.
Table 3
Mean (±SD) length (mm) and width (mm) of the longest leaf in generative stems of Inula salicina per subplot in unburned and burned patches of Molinion caeruleae meadow in 2017–2019.
Table 4
Mean (±SD) number of inflorescences in generative stems and diameter (mm) of the receptacle of the largest inflorescence of Inula salicina per subplot in unburned and burned patches of Molinion caeruleae meadow in 2017–2019.
. Temporal variability of the number and traits of Inula salicina stems
The part of the third hypothesis stating that the numbers of vegetative and generative Inula salicina stems do not present temporal variability might be partially accepted. The number of vegetative and generative stems in the unburned patch and the number of vegetative stems in the burned patch (Table 1) did not differ among the particular years of observation. However, the number of generative stems in the burned patch differed significantly among the particular years of observation and had the greatest value in 2019 (Table 1).
The part of the third hypothesis stating that the individual traits of Inula salicina do not present temporal variability must be partially rejected. In the unburned patch, the height of generative stems (Table 2), the number of leaves (Table 2), the length of the longest leaf (Table 3), and the number of inflorescences in the generative stem (Table 4) achieved significantly greater values in 2019 than in the earlier years. The diameter of the receptacle of the largest inflorescence (Table 4) and the width of the longest leaf (Table 3) did not vary in 2017–2019.
In the burned patch, the height of generative stems (Table 2) was similar in the whole study period. The number of leaves (Table 2) and the number of inflorescences in generative stems (Table 4) were greater in 2019 than in the other seasons. The diameter of the receptacle of the largest inflorescence (Table 4) achieved much lower values in 2017 than in the following seasons. The width of the longest leaf (Table 3) was lower in 2019 than in the previous seasons. The length of the longest leaf (Table 3) reached its greatest value in 2018.
. Discussion
The results showing a greater number of vegetative and generative stems in the burned patch compared to the unburned site may be surprising, considering the studies conducted by Valkó et al. (2018), who evidenced a much greater abundance of the close congener Inula ensifolia in untouched grasslands than in those subjected to prescribed burning. The aboveground stems of Inula salicina observed in the burned patch may have originated from rhizomes. Such a phenomenon is probable, considering previous findings demonstrating the response of individuals of the aforementioned species to injury. Several authors have noted that removal of total aboveground biomass of juvenile individuals at 1 cm above the topsoil level does not lead to the death of the individual (Martínková et al., 2023a) but substantially reduces above- and below-ground biomass (Martínková et al., 2023b) and does not contribute to the formation of adventitious root buds (Filartiga et al., 2022). Bucharova et al. (2024) added that clipping of aboveground parts of Inula salicina individuals boosts vegetative reproduction. On the other hand, the much greater number of vegetative and generative stems in the burned patch may be an effect of the germination of anemochorous achenes dispersed from adjacent localities directly after the accidental burning and the successful colonization of safe sites for seedling recruitment (sensu Harper et al., 1965). Such a phenomenon seems to be probable, especially considering the findings reported by Burmeier et al. (2010), who noticed that Inula salicina showed maximum seedling emergence from seeds sown directly on the soil surface, while the increasing burial depth led to decreased germination, emergence, and growth of individuals. Taking into account the findings reported by Florianová et al. (2022), we can speculate that Inula salicina may have colonized the disturbed site earlier than other plants and became the dominant species. Moreover, the domination of Inula salicina may have been strengthened by allelopatic activity inhibiting the germination of other species (Solymosi, 2012). Moreover, it is worth mentioning that Johanidesová et al. (2015) observed that the natural seed dispersal from populations resident in adjacent patches contributes to the appearance of Inula salicina in restored dry grasslands.
Although previous observations conducted in experimental conditions (Hölzel & Otte, 2004) and natural localities (Kostrakiewicz-Gierałt, 2015) demonstrated the potential for germination of Inula salicina directly after shedding in late summer and autumn, as well as in spring of the following year, the present observations showed a lack of seedlings at both study sites. This fact may be surprising, especially in light of the findings reported by Schmiede et al. (2013), who evidenced that Inula salicina can germinate on the surface and below the litter layer. On the other hand, considering several other findings (Dostálek et al., 2022; Eckstein & Donath, 2005; Ludewig et al., 2014), we may speculate that shade and intermittent drought occurring beneath the litter layer could inhibit seed germination and contribute to a lack of seedlings in both study patches. Moreover, according to the findings shown by Donath and Eckstein (2010), we may suppose that the seedling recruitment was hampered by the bryophyte cover. At the same time, the activity of herbivorous animals contributing to seedling damage or even absence should be mentioned. According to Hensgen et al. (2011), specifically young seedlings are prone to slug herbivory, while Těšitel et al. (20152021) evidenced considerable damage to Inula salicina leaves caused by insects. Additionally, it is worth adding that the lack of seedlings may be a consequence of diminished production of achenes caused by insect attack on receptacles, as observed by Rohfritsch and Arnoldrinehart (1991).
The lack of uniform spatial variability of individual traits of Inula salicina during the subsequent study seasons may indicate that observations of burning effects should be conducted over a longer period. The lack of temporal variability in the number of vegetative and generative stems in the unburned patch may be caused by the moderate rate of clonal multiplication reaching from 1 to 10 offspring stems per year (Klimešová et al., 2017). On the other hand, the considerable increase in the number of generative stems in the burned site is consistent with the findings reported by Pyke et al. (2010), who observed that cryptophytes resprouting from basal or root buds after aboveground buds are burned respond quickly to post-fire nutrient flushes by growth or coppicing.
The greatest values of the surveyed individual traits (i.e. height of generative stems, number of leaves and inflorescences) in 2019 may be a response to progressive secondary succession and the overgrowing of the study sites by competitors such as Phragmites australis in the case of the unburned patch and Salix repens var. rosmarinifolia in the case of the burned patch (Wójcik et al., 2022). This phenomenon is consistent with the findings shown by Bartušková et al. (2015) and Bucharova et al. (2024), who documented markedly higher investments in stems and reproductive organs of Inula salicina growing in unused patches compared to mown plots. Considering the findings reported by Klecka et al. (2018), we may speculate that the increasing height of generative stems, number of inflorescences, and diameter of receptacles of Inula salicina might influence plant-pollinator interactions and augment the visitation rate. A similar phenomenon of increased allocation in generative reproduction along the gradient of vegetation height was noticed in a population of Serratula tinctoria in an abandoned Molinion caeruleae meadow (Kostrakiewicz-Gierałt & Bąba, 2014). The aforementioned authors observed a significantly greater stem height, number of capitula, and mean number of seeds per capitulum in individuals growing in a patch overgrown by shrubs and trees than in a patch dominated by large-tussock grasses and in a patch dominated by species with a low height.
To sum up, we may conclude that controlled burning may contribute to regeneration of Inula salicina populations in abandoned meadows. The damage to plant cover and litter caused by fire enables successful vegetative or/and generative propagation of the analyzed species. However, the persistence of populations at occupied sites and the condition of individuals in a changing environment should be monitored. Moreover, further studies on the post-fire regeneration of other meadow species from the soil seed bank and underground organs are still required.
. Conclusions
The significantly greater number of vegetative and generative stems in the burned patch compared to the unburned patch during the whole study period might be a consequence of vegetative spread from underground organs or/and successful seedling recruitment and site colonization directly after the disturbance and earlier than other plants. The absence of seedlings of Inula salicina during the study period at both study sites might be caused by unfavorable environmental conditions, such as local water deficiency, shade beneath the litter layer, or herbivore attack. The lack of uniform spatial variability of the individual traits of generative stems, such as height, number, and dimensions of leaves, number of inflorescences, and diameter of the receptacle of the largest inflorescence, suggests that observations of the effects of burning should cover a longer period of time. The lack of temporal variability in the number of vegetative and generative stems may be caused by the moderate rate of clonal multiplication influencing the appearance of offspring stems produced per year. The temporal increase in selected traits of generative stems noticed in both study patches may be a response to progressive secondary succession and the overgrowing of both study sites.
To sum up, controlled burning may contribute to regeneration of populations of Inula salicina in abandoned meadows. However, the persistence of populations at occupied sites and the condition of individuals in a changing environment should be monitored. Moreover, considering the promising results of the present investigations, it may be concluded that further studies on the post-fire regeneration of other meadow species from the soil seed bank and underground organs are still required.
