. Introduction
The genus Cistus L., belonging to the family Cistaceae (eight genera and 180 species), is composed of perennial shrubs primarily distributed in the Mediterranean region (Arrington and Kubitzki, 2003). One of the most abundant and widely distributed species in the genus is Cistus ladanifer L., whose distribution area ranges from Portugal and Morocco to the French Riviera and Algeria (Guzmán and Vargas, 2009). This species is of great interest both for its ecological role in maintaining Mediterranean ecosystems and for its value as a natural resource. Regarding its ecological importance, it is considered a pioneer plant, capable of quickly colonizing disturbed areas, such as those affected by forest fires, thanks to its high resprouting and germination capacity. Its recovery after fires is faster compared to other shrub species due to its heat resistance and the breaking of seed dormancy by high temperatures (Luna et al., 2022). This makes it the dominant species after the first or second-year post-fire (Correia and Ascensão, 2017). As a natural resource, the species’ trunk, branches, and leaves are impregnated with labdanum, a viscous substance. This compound is highly valued in the perfume industry for its warm and sweet aroma, as well as its excellent fixative properties that help prolong the duration of other fragrances in perfumes (Papaefthimiou et al., 2014). In fact, it is an essential component in many modern perfumes. Additionally, this species is also known to have applications in human and animal food, medicine, veterinary, fuel, construction, crafts, and environmental uses (Frazão et al., 2024).
The exploitation of wild C. ladanifer populations to obtain labdanum is a common practice with a long tradition in Mediterranean countries (Pardo de Santayana et al., 2018). In fact, these exploitation practices are being studied to improve them through the promotion of mechanization and the development of cultivation (Mariotti et al., 1997; Mediavilla et al., 2021). The wide ecological tolerance of the species enables its successful cultivation across diverse environmental conditions (Raimundo et al., 2018). This is reflected in the species’ ability to survive in a wide range of altitudes and soil types and in dry areas with high insolation (Núñez-Oliveira et al., 1996). It also shows high stress tolerance allowing it to compete in diverse environmental conditions (Rossini-Oliva et al., 2016). These include soils poor in organic matter, low pH, and high concentrations of trace elements, water stress, high temperatures, and solar radiation (Alados et al., 1999; Kidd et al., 2004).
The present study aims to explore the reproductive ecology of C. ladanifer in its natural populations, specifically its ability to interact with other species of the genus. This knowledge is of interest in the current context of developing the potential of this species as a natural resource. Evidence exists of hybrid individuals appearing in natural areas where C. ladanifer coexists with other congeners (Demoly, 1996; Demoly and Montserrat, 1993). This could indicate an ability of C. ladanifer to pollinate and fertilize them. It is known that pollen transfer from one species to another can have genetic and demographic consequences for the recipient species (Allendorf et al., 2001; Rieseberg et al., 1989). One of them is genetic introgression, which can alter the genetic composition of native populations (Ellstrand, 1992; Rhymer and Simberloff, 1996). Another is the reduction of the reproductive capacity of the pollen recipient species, which can diminish its offspring and affect the maintenance of natural populations (Levin et al., 1996; Wolf et al., 2001). The demographic and genetic effects can cause ecosystem alterations by changing population dynamics (Todesco et al., 2016).
The objective of this study is to investigate the pollen dispersal pattern from C. ladanifer to C. laurifolius L. Specifically, it aims to answer the question of whether the pollen of the former species can be transferred to its congener, and if so, to understand the dispersal pattern. The main hypothesis of this work is that C. ladanifer and C. laurifolius share common pollinators, allowing these pollinators to bidirectionally transport pollen between C. ladanifer and C. laurifolius plants. However, this study focuses exclusively on pollen dispersal from Cistus ladanifer to Cistus laurifolius, given the increasing interest in the exploitation and cultivation of C. ladanifer, which makes it essential to assess the potential for genetic transfer between this species and its congeners. The natural occurrence of Cistus × cyprius Lam., a hybrid taxon between these two species described in 1786, suggests the existence of pollen flow and gene exchange in both directions—either from C. ladanifer to C. laurifolius or vice versa. Numerous herbarium specimens of the hybrid taxon have been collected from overlapping distribution areas of C. ladanifer and C. laurifolius in the western Mediterranean region (southeastern France, central, eastern, and southeastern Spain, and northern Morocco) (GBIF, 2025). Although there are no detailed studies on the frequency of this hybrid in populations where both parental species coexist, several works and field observations indicate that its presence is relatively common (Dansereau, 1941; Demoly, 1996; GBIF, 2025; Martín and Guinea, 1949). The morphological characteristics of C. × cyprius have been studied and compared with those of the parent species, revealing a combination of traits: some resembling one or the other parent, some intermediate, and some expressing features of both parents simultaneously (Carazo and Jiménez, 1989; Dansereau, 1951). Diagnostic traits used to identify this hybrid include leaf margin characteristics, trichome size and type on floral peduncles, inflorescence type, sepal shape, and the number and morphology of fruit valves. Specifically, in the present study, pollen flow from C. ladanifer to C. laurifolius was investigated in two natural mixed populations located in the Iberian Peninsula, with the aim of determining: (1) whether pollen dispersal from C. ladanifer to C. laurifolius occurs; and (2) if so, whether the pollen dispersal pattern is influenced by the distance from the pollen-emitting plant.
. Materials and methods
. Study species
Cistus ladanifer inhabits shrublands on siliceous or calcareous soils, within an altitudinal range of 0–1500 m a.s.l., and is distributed across the Iberian Peninsula, France, and North Africa (Guzmán and Vargas, 2009). Cistus laurifolius is typical of oak and holm oak forests on slopes and rocky, siliceous, and decalcified calcareous soils, ranging from 400–1900 m a.s.l. Its distribution includes Greece, Turkey, Italy, France, Spain, Portugal, and Morocco (Astuti et al., 2017).
Both species are woody shrubs up to 2 m tall with clear morphological differences. Cistus ladanifer has reddish-brown stems and sessile, lanceolate leaves that are sticky due to the presence of labdanum. Cistus laurifolius has grayish stems and larger, ovate-lanceolate leaves with wavy margins and petioles (Demoly and Montserrat, 1993). Cistus ladanifer presents solitary terminal flowers with short peduncles, while those of C. laurifolius are grouped in pubescent cymose inflorescences of 1–9 flowers. The flowers of C. ladanifer have large petals (30–55 mm), and there are two color varieties: albiflorus (white petals) and maculatus (white petals with a reddish spot). The flowers of C. laurifolius are smaller and always white. Both species have numerous stamens and produce abundant pollen (Figure 1). Cistus ladanifer has a tomentose ovary with a large sessile stigma, while C. laurifolius has a silky ovary with a pentagonal stigma. Cistus ladanifer blooms from March to June, and C. laurifolius from May to July, with partial overlap between the two species (Demoly and Montserrat, 1993). Both taxa are xenogamous and require pollinators to produce seeds, primarily insects from the orders Diptera, Coleoptera, and Hymenoptera (Astuti et al., 2017; Talavera et al., 1993; Teixido and Valladares, 2014). They produce lignified, globose capsule fruits with numerous seeds lacking long-distance dispersal structures (Demoly and Montserrat, 1993).

. Study site
Fieldwork was conducted between March and June 2023 in two locations situated in the Sierra de Guadarrama, northwest of the Community of Madrid (Spain): Miraflores (latitude: 40,752332°, longitude: –3,991522°; municipality: Miraflores de la Sierra, 1362–1389 m a.s.l.) and La Barranca (latitude: 40,856541°, longitude: –3,743657°; municipality: Navacerrada, 1193–1204 m a.s.l.) ( Figure 2). Both study sites contain mixed populations of Cistus ladanifer and C. laurifolius. Specimens of both species are often found in clusters of several plants, rarely isolated ( Figure 3). In preliminary observations before conducting the pollen analog dispersal experiment (fluorescent dye) (described in the next section), C. × cyprius (= C. ladanifer × C. laurifolius) individuals were detected at the two study sites. In La Barranca and Miraflores, plants of both species appear in clearings, surrounded by sclerophyllous vegetation and bordered by deciduous forests or meadows, in sloping areas.
Figure 2
Location of the study site. (A) Map of the area. (B) Study site at the Sierra de Guadarrama (Community of Madrid). (C) Location of plants studied at La Barranca. (D) Location of plants studied at Miraflores.
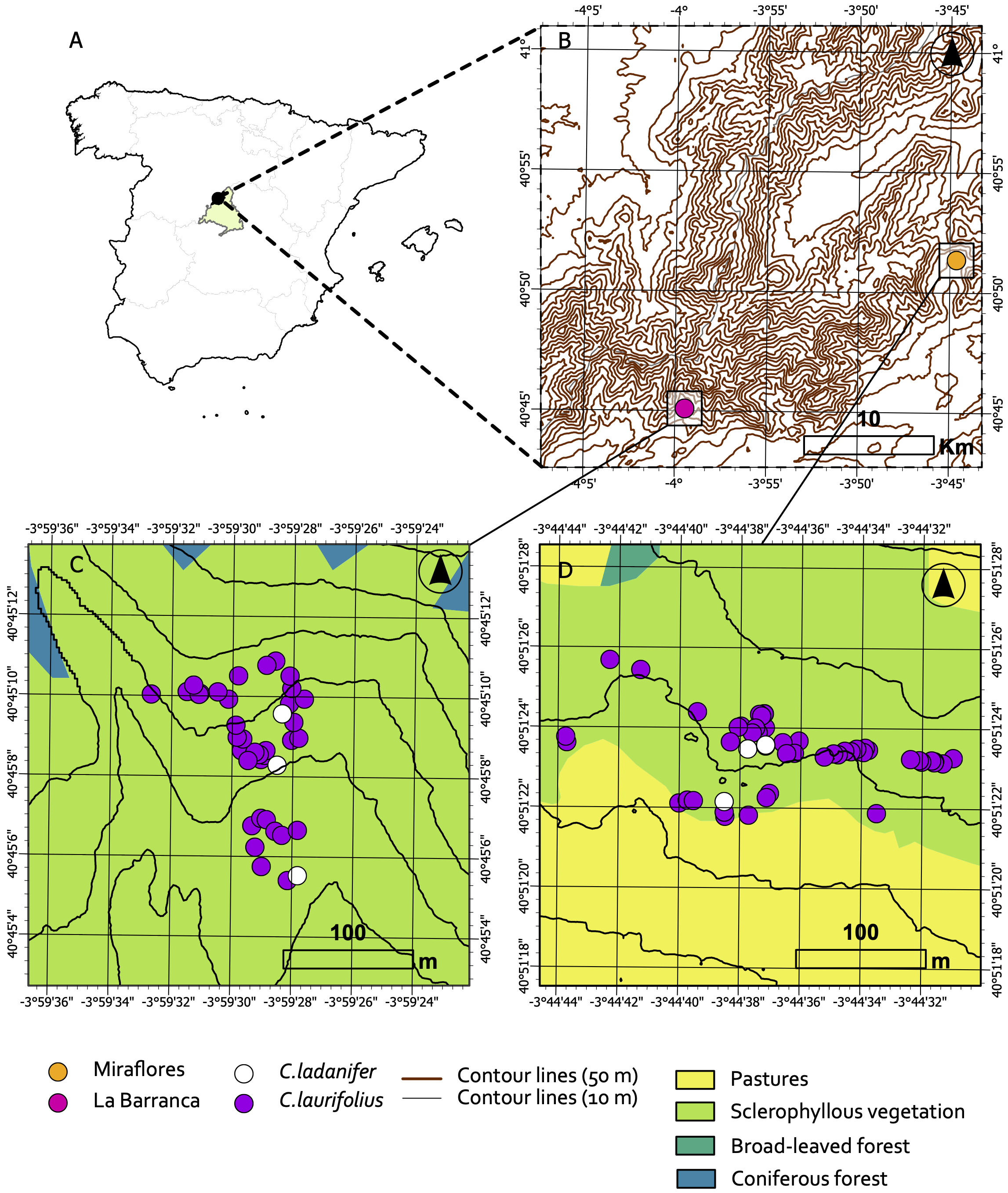
Figure 3
Photograph of La Barranca taken on 05/06/2023 showing Cistus ladanifer and C. laurifolius individuals in bloom growing together. White circles indicate C. ladanifer individuals and purple circles indicate C. laurifolius individuals. Not all but only a few observed specimens of both species are highlighted.

. Pollen dispersal experiment using fluorescent dye
To study pollen dispersal from Cistus ladanifer to C. laurifolius within the period of floral synchrony of both species, periodic visits were made to the two study sites in 2023. These visits took place between March and May (every 15–20 days in March and April and every 3–5 days in May). In Miraflores, the period of floral synchrony began the week of May 8, while in La Barranca it began the week of May 29. Between 5–8 days after, pollen flow from C. ladanifer plants to C. laurifolius plants was estimated by studying the dispersal of pollen analogs (fluorescent dye). This methodology has proven effective in estimating the distribution of dispersed pollen in natural populations and has been employed in numerous studies (Huais et al., 2022; Van Rossum and Triest, 2010, 2012). For this experiment, the following steps were followed here and repeated three times at each study site (Table 1): 1) application of pollen analogs (fluorescent dye) on C. ladanifer flowers; 2) collection of C. laurifolius flowers and observation of the stigmas under an epi-fluorescence microscope.
Table 1
Study site and start date of the pollen analog dispersal experiment (fluorescent powder) along with the geographic coordinates of the emission source and the number of treated flowers (Cistus ladanifer) and collected flowers (C. laurifolius) at the two study sites
Fluorescent dye was applied to all flowers of one or two randomly selected C. ladanifer individuals (hereafter referred to as the emission source). Yellow fluorescent dye (Radiant Color Corp.) was chosen for the experiment. Before applying it, it was tested whether flowers treated with the fluorescent dye attracted species from the main pollinator groups visiting the species. A previous study in the area recorded visits from five functional groups: bumblebees (Bombus spp.), solitary bees (Andrenidae, Colletidae, and Halictidae), honeybees (Apis mellifera), hover flies (Syrphidae), and beetles (Coleoptera) (Teixido and Valladares, 2014). After confirming visits to the flowers by the mentioned pollinator groups, the fluorescent dye was applied using fine-tipped brushes (size 0–2) or wooden sticks with blunt tips on the flower anthers. The application of the fluorescent dye was done on sunny days between 8:00 a.m. and 10:00 a.m. (Figure 4a). All selected C. ladanifer plants were georeferenced using a GPS (Garmin Etrex 32×).
Figure 4
Treated Cistus ladanifer flower and stigma sampled from C. laurifolius. (A) Flower with pollen analogs (fluorescent dye) applied to the anthers. (B) Stigma collected at La Barranca on 14/06/2023 and observed with an epi-fluorescence microscope under UV light. Photographic montage of 72 images at 200× magnification taken with Viewfinder Lite (version 1.0) and subsequently processed with Studio Lite (version 1.0.131). The image composition was done using Freeform (version 2.4).

After treating the C. ladanifer flowers with the fluorescent dye, C. laurifolius plants (8–14 plants) with open flowers at different distances from the emission source were selected and marked. Random walks from the emission source were conducted for this purpose. The selected plants were representative of the spatial distribution at the study site and covered a wide range of distances from the emission source (8–167 m). Although the initial aim was to sample C. laurifolius plants up to 200 m from the emission source, limitations due to fencing or the absence of flowering plants reduced this distance. The geographic position of the selected plants was recorded using a GPS, and between 5 and 20 open flowers from each plant were tagged. These flowers were not treated to allow free pollination by insects. After one day (the observed lifespan of the flower at the study site), the flowers were collected and stored for 24–48 hours at 4°C. Subsequently, the stigma from each flower was separated, mounted on a slide, and observed under an epi-fluorescence microscope under UV light to check for the presence of the fluorescent dye (200×) and to count the number of fluorescent dye particles (400×).
. Data analysis
To investigate the pollen flow from Cistus ladanifer to C. laurifolius within and between the study sites, data on particle deposition (fluorescent dye) observed on the stigmas of C. laurifolius were used. For this purpose, the deposition rate and deposition intensity were obtained. The deposition rate was the proportion of stigmas on each sampled plant that contained pollen analog particles (fluorescent dye). The deposition intensity was calculated as the average number of pollen analog particles (fluorescent dye) counted on the stigmas of flowers collected from each plant. Finally, the deposition rate and intensity for each plant were associated with the distance (in meters) of the plant from the emission source, which was calculated using ArcGIS Pro (v. 3.0). These data were used to generate a dataset for each study site by combining all data obtained during the three days that the experiment was repeated at each study site. The minimum and maximum dispersal distances of pollen analogs (fluorescent dye) were obtained. The minimum dispersal distance was considered the distance of the C. laurifolius plant with pollen analogs found closest to the emission source, while the maximum dispersal distance was the distance of the C. laurifolius plant with pollen analogs found farthest from the emission source.
To estimate the general pattern of pollen dispersal, descriptive statistics and frequency histograms were obtained for the deposition rate and deposition intensity at each study site. The Mann-Whitney U test was used to assess differences between populations in these parameters. To evaluate the relationship between the deposition rate and intensity, a Spearman correlation analysis was performed. Finally, the deposition rate and intensity per plant at each study site were analyzed in relation to the distance of the plants to the emission source using a Generalized Linear Model. For the model, a binomial distribution and logit link function were used to analyze the deposition rate, and a Poisson distribution and log link function were used to evaluate the deposition intensity (Zuur et al., 2009). These analyses were performed using the Past4 software (version 1.0.6).
Additionally, the proportion of pollen analog particles (fluorescent dye) deposited on the stigmas of C. laurifolius plants in relation to their distance from the emission source was calculated. Each dataset was ordered by the distance variable (distance of each sampled C. laurifolius plant to the emission source, from the lowest to the highest observed distance). Then, an accumulated particle number was calculated using the average particle count value for each increasing distance. The accumulated number of particles at each observed distance was divided by the total number of particles (obtained by summing the average particle count value for each distance). This analysis allowed the determination of specific particle percentages (Huais et al., 2022) and served to determine the distances at which 50% (D50), 80% (D80), and 90% (D90) of the particles deposited on C. ladanifer flowers were observed at each study site.
To assess whether there was a spatial pattern of pollen analog (fluorescent dye) dispersal, a Moran’s I spatial autocorrelation analysis was performed for the deposition rate and intensity using inverse Euclidean distance values (Bivand and Wong, 2018). The analysis was conducted for each study site using the ArcGIS Pro (v. 3.0).
. Results
Overall, 90.27% of the sampled Cistus laurifolius plants had at least one stigma with pollen analogs ( Figure 4b). The percentage was higher in Miraflores de la Sierra (97.27%) than in La Barranca (78.57%). Table 2 shows the statistics for the deposition rate and deposition intensity of pollen analog particles (fluorescent dye) at the two study sites. Both the deposition rate and intensity were notably higher in Miraflores compared to La Barranca. In Miraflores, the average value for the deposition rate was 0.783 and for deposition intensity was 79.588, while in La Barranca, these values were 0.355 and 6.493, respectively. Lower variability, as reflected in the standard errors of both parameters in Miraflores, indicated greater consistency in the deposition measurements of pollen analog particles at this site. The statistical analysis of these data revealed significant differences between the two sites for both measurements (deposition rate: Mann-Whitney U = 162.5, z = 5.270, P < 0.001; deposition intensity: Mann-Whitney U = 5.46, z = 5.460, P < 0.001).
Table 2
Mean (± standard error), median, range (minimum‑maximum), and the number of plants and flowers sampled for the analysis of the deposition rate and deposition intensity at the two study sites: Miraflores de la Sierra and La Barranca.
The comparison of frequency histograms for the deposition rate and deposition intensity highlighted differences between the populations (Figure 5). In Miraflores, there was a higher frequency of plants with high deposition rates. Most observations were concentrated in deposition rates ranging from 0.5 to 1.0, with the highest number of observations in the 0.8 to 1.0 range. In contrast, in La Barranca, the distribution of values was more uniform, and low deposition rates were more frequent. Regarding the deposition intensity, the frequency histogram in Miraflores showed that most deposition intensity values were concentrated in the 0 to 50 range, with few observations at higher intensities, while in La Barranca, most observations were also concentrated at low values, within the 0 to 10 range, with few observations at higher intensities.
Figure 5
Frequency histogram for the deposition rate and deposition intensity in Miraflores and La Barranca.
A significant positive correlation between the deposition rate and deposition intensity was observed at both study sites, although higher in La Barranca (Miraflores: rs = 0.701, P < 0.01; La Barranca: rs = 0.920, P < 0.01), indicating that C. laurifolius plants with a higher deposition rate also showed a higher deposition intensity of pollen analogs (fluorescent dye).
The maximum and minimum dispersal distances of pollen analogs (fluorescent dye) detected at both study sites are shown in Table 3. In Miraflores, pollen analogs (fluorescent dye) were found on C. laurifolius plants situated at distances between 9.56 m and 144.91 m from the emission source. This range coincided with the distances of the nearest and farthest sampled plants from the emission source. In La Barranca, the nearest sampled individual to the emission source was at 8.60 m, but the minimum dispersal distance obtained was greater (19.84 m). In contrast, the maximum dispersal distance was similar to that of the farthest sampled plant from the emission source.
The analysis of the accumulated pollen analog particles (fluorescent dye) in relation to the distance from the emission source is shown in Table 3. In Miraflores, the results showed that 50%, 80%, and 90% of the pollen analog particles (fluorescent dye) deposited on the stigmas of C. laurifolius were concentrated, respectively, at distances up to 16.70 m (D50), 51.00 m (D80), and 74.00 m (D90). In La Barranca, these results were similar, with 50%, 80%, and 90% of the particles concentrated at distances up to 18.20 m (D50), 56.00 m (D80), and 81.00 m (D90) from the emission source.
Table 3
Distance of sampled Cistus laurifolius plants from the C. ladanifer emission source, detected dispersal distance of pollen‑analog particles (fluorescent dye), and D50, D80, and D90 values for the two study sites: Miraflores de la Sierra and La Barranca.
The deposition intensity of pollen analog particles (fluorescent dye) on C. laurifolius plants was affected by the distance to the C. ladanifer emission source at both Miraflores and La Barranca (Miraflores: G = 1635.1, P < 0.00001; La Barranca: G = 117.74, P < 0.00001) (Figure 6). Conversely, the statistical analysis did not support an effect of the distance to the emission source on the deposition rate of pollen analog particles (fluorescent dye) at either study site (Miraflores: G = 1.5373, P = 0.2150; La Barranca: G = 0.6298, P = 0.4274).
Figure 6
Deposition rate and deposition intensity of pollen analogs (fluorescent dye) on C. laurifolius plants in relation to the distance from the C. ladanifer emission source in Miraflores and La Barranca. The curve indicates the expected values according to the statistical model (GLM).
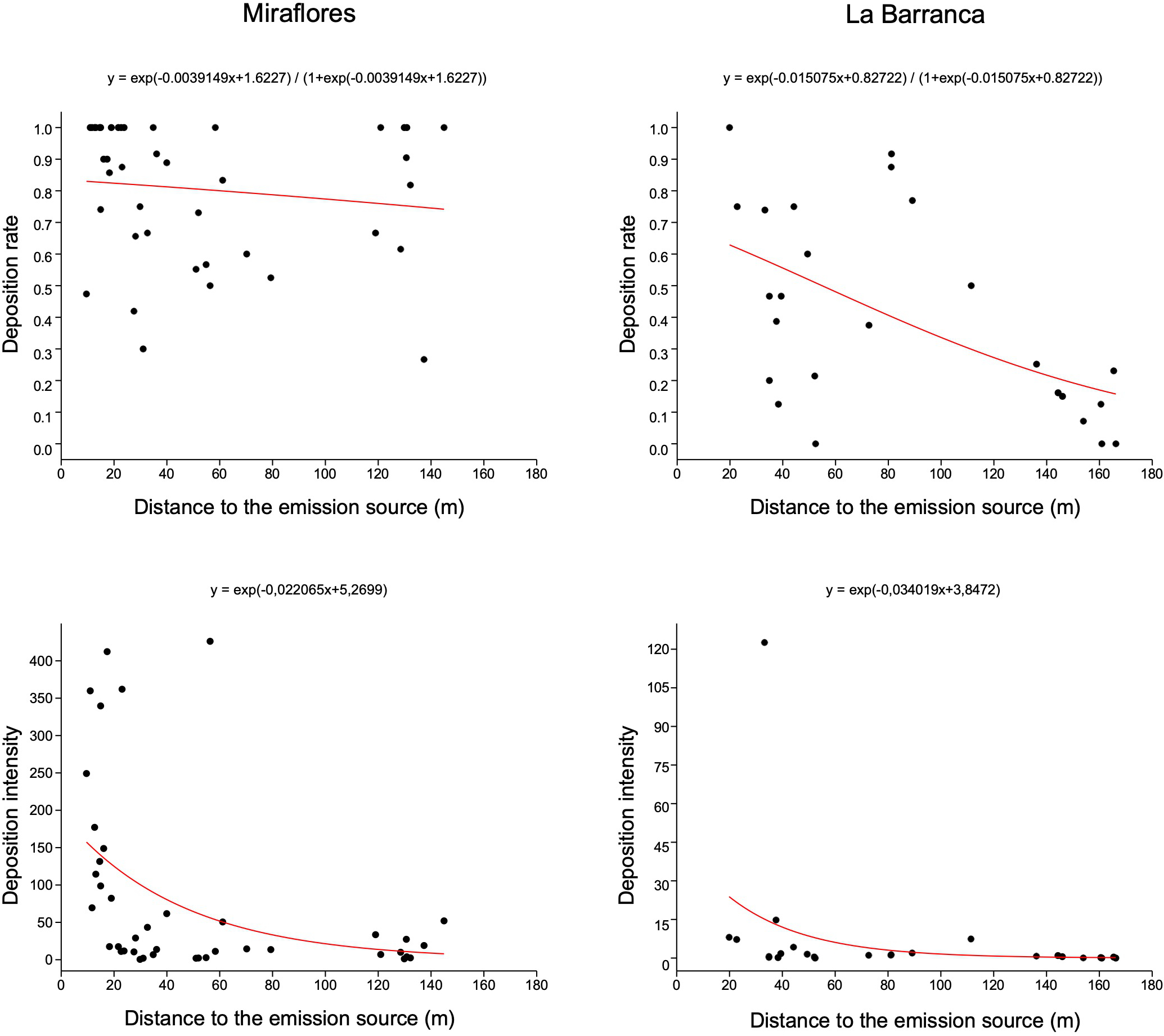
The results from the Moran’s I spatial autocorrelation analysis evidenced spatial patterns in both the deposition rate and intensity at both study sites (Table 4). In both Miraflores and La Barranca, the deposition rate and intensity followed a spatial clustering pattern. This pattern was statistically significant in all cases except for the deposition rate in Miraflores, although the probability that the obtained spatial clustering pattern could be due to chance was less than 10%.
Table 4
Summary of the results of the Moran’s I spatial autocorrelation analysis for the deposition rate and deposition intensity at the two study sites: Miraflores de la Sierra and La Barranca.
| Deposition rate | Deposition intensity | |||
|---|---|---|---|---|
| Miraflores | La Barranca | Miraflores | La Barranca | |
| I | 0.3060 | 0.3561 | 0.1625 | 0.1222 |
| z | 2.8458 | 2.7463 | 1.6595 | 4.5598 |
| z | 0.0044 | 0.0060 | 0.0970 | 0.0000 |
. Discussion
. Pollen analog dispersal pattern
In the present study, Cistus ladanifer pollen grain analogs were detected on the stigmas of C. laurifolius. Although the rate and intensity of pollen analog deposition varied between the two study sites – likely due to differences in ecological conditions between the populations – the results clearly demonstrate the ability of C. ladanifer to pollinate C. laurifolius at both locations. This capacity is likely attributable to the high morphological similarity between the flowers of C. ladanifer and C. laurifolius, which promotes the attraction of the same insect pollinator species. Both species produce large hermaphroditic flowers with white petals, abundant pollen, and are visited by generalist pollinators belonging to the orders Coleoptera, Diptera (mainly syrphid flies), and Hymenoptera (primarily Apis mellifera and solitary bees) (Astuti et al., 2015, 2017; Talavera et al., 1993).
The similarity of floral traits between species facilitates a greater degree of pollinator sharing (Bergamo et al., 2020; Fornoff et al., 2017), as they respond to similar sensory stimuli (Faegri and Van der Pijl, 1979; Suárez-Mariño et al., 2022). Thus, when species within the same community bloom simultaneously, exhibit morphologically similar flowers, and share pollinators, cross-pollination between them is common (Fang et al., 2019; Johnson et al., 2019). Consequently, the ability of C. ladanifer to pollinate C. laurifolius was predictable, considering the reproductive biology and shared pollination ecology between both species.
Cistus ladanifer plants were able to pollinate C. laurifolius plants located at least 165 meters away. This distance is consistent with the distances measured in previous studies on pollen dispersal in entomophilous pollination systems, which also used fluorescent dye (Van Rossum et al., 2013). Moreover, in the two sites where the present study was conducted, the same dynamics of pollen analog dispersal were evidenced. This was characterized by a downward trend, meaning that C. laurifolius plants closest to the focal C. ladanifer plants received a greater amount of pollen analogs on their stigmas than those further away. Specifically, 80% of the pollen analogs found on the flowers of C. laurifolius were transferred from donor plants within a distance of less than 60 meters.
The pattern of pollen analog dispersal observed in this study is consistent with most research on pollen dispersal in entomophilous plants, which indicates that pollen dispersal follows a leptokurtic distribution, with the majority of pollen deposited near the source of emission (Kameyama et al., 2000; Tonsor, 1985). This phenomenon is attributed to the propensity of pollinators to make short flights between adjacent plants (Klinger et al., 1992). The behavior and movements of pollinators result in the predominant deposition of their pollen load on the first individuals visited from the source (Cresswell et al., 1995; Morris et al., 1995).
. Ecological and conservation implications
The transfer of pollen by pollinators from a species to a congener can have significant negative repercussions on the latter. Firstly, it can affect various stages of reproduction, such as conspecific pollen germination, pollen tube growth, and ovule fertilization, consequently reducing seed production (Ashman and Arceo-Gómez, 2013; Ashman et al., 2020) and impacting population size. Secondly, pollen transfer can also result in spontaneous hybridization (Fan et al., 2014; Wang et al., 2019), which may lead to population and/or species extinction through genetic assimilation (Levin et al., 1996; Wolf et al., 2001) or the generation of highly invasive genotypes (Schierenbeck and Ellstrand, 2009). Due to the potential negative consequences mentioned and given that this study has demonstrated the ability of C. ladanifer to pollinate C. laurifolius plants, future cultivation experiences of C. ladanifer should avoid affecting wild populations of C. laurifolius.
These findings provide a preliminary basis for proposing a series of practical preventive recommendations in case of the establishment of C. ladanifer crops. First, the cultivation of C. ladanifer should be prioritized in areas outside the distribution range of C. laurifolius. This recommendation is particularly important in Italy, where C. laurifolius is considered a threatened species that could be seriously impacted by the proximity of nearby C. ladanifer crops. Second, if cultivation is to be established within the distribution area, a minimum preventive separation distance from wild populations of C. laurifolius should be maintained. This distance should be assessed through a detailed study to measure the true pollen dispersal distance of C. ladanifer when the plants are under cultivation conditions. In these circumstances, there may be a variation in local pollinator assemblages (Alarcón et al., 2008), which could generate spatial and temporal variations in the movement rates between species by the pollinators (Fang et al., 2019), and consequently could affect the pollen dispersal distance.
. Conclusion
This study on the pollen analog dispersal is one of the few attempts made so far (Van Rossum et al., 2013) to estimate pollen dispersal patterns between congeners in natural populations. Our findings demonstrate the potential of C. ladanifer plants to pollinate nearby C. laurifolius plants. Therefore, our results emphasize the importance of considering pollen dispersal experiments when establishing new cultivations of wild species within the distribution range of other congeners.
