. Introduction
Theobroma cacao L. is a widely valued species in the food, pharmaceutical, and cosmetic industries because of its multiple applications (Mendoza-Meneses et al., 2023). Its cultivation has historical and economic relevance in the regions that produce it, as it is a source of employment and well-being for its inhabitants. In Mexico, this crop has been cultivated for more than 500 years. Phytosanitary problems represent the main limitation for its production.
The most important diseases affecting T. cacao are moniliasis, caused by Moniliophthora roreri, which causes production losses of up to 75%, and cocoa black pod (CBP) disease, caused by Phytophthora spp., which results in losses of 6 to 25% (Torres-de la Cruz et al., 2023). To date, only two species of Phytophthora associated with CBP have been identified in Tabasco, Mexico: Phytophthora tropicalis (P. tropicalis) (Chávez-Ramírez et al., 2021) and P. capsici (Torres-de la Cruz et al., 2023). Phytosanitary problems, among other problems, have led many producers to abandon cocoa plantations or allocate soils to more profitable crops (Oporto-Peregrino et al., 2020), causing a shortage of products and putting a rich cultural heritage associated with this crop at risk.
In the context of sustainable agriculture, the need to develop biotechnologies free of toxic residues that are affordable and accessible to producers and that control cocoa pests and diseases is urgent. Various studies in the humid tropics of Mexico have explored sustainable routes for the control of moniliasis (Ortiz-García et al., 2015; Reyes-Figueroa et al., 2016; Torres-Rodríguez et al., 2024) and have recently explored the use of plant extracts. The Capsicum genus is rich in secondary metabolites, and extracts obtained from its fruits have potential for the control of various phytopathogens (Bacon et al., 20242017), including M. roreri (De la Cruz-Ricardez et al., 2020) and P. megakarya (Simo et al., 2019). An important characteristic of plant extracts is that they are made up of a mixture of bioactive compounds, such as phenols, terpenes, and alkaloids, which have different modes of action against pathogens (Chtioui et al., 2023). This not only helps to prevent the development of pathogen resistance to treatments, but also avoids the generation of toxic residues that can negatively impact the environment (Chtioui et al., 2023; Zhou et al., 2023).
Considering the importance of the control of CBP in cocoa plantations, the absence of sustainable alternatives for its control in the humid tropics of Mexico, and given that various species of Capsicum coexist with the cacao agrosystem, including C. annuum var. glabriusculum, whose fruit extracts contain phenolic acids (De la Cruz-Ricardez et al., 2024), which have demonstrated antifungal activity (De la Cruz-Ricardez et al., 2020), the objective of this research was to evaluate the in vitro and in vivo effects of methanolic extracts (MEs) of phenolic compounds extracted from the leaves and immature and mature fruits of C. annuum var. glabriusculum morphotypes Amashito and Garbanzo, C. chinense Jacq., and C. frutescens L. on P. tropicalis, the causative agent of CBP.
. Materials and methods
. Pathogen identification
The pathogen was provided by the Laboratory of Plant Pathology at the Colegio de Postgraduados, Campus Tabasco, Mexico. Molecular identification was carried out by the National Laboratory of Agricultural, Medical, and Environmental Biotechnology at the Potosino Institute for Scientific and Technological Research (IPICYT) in San Luis Potosí, Mexico. Sequencing of the ITS region was performed with primers ITS1 and ITS4 (White et al., 1990). Editing and assembly of the sequences were performed in the Chromas® 2.6.6 program. Subsequently, a homology search using the BLAST® tool of the National Center for Biotechnology (NCBI) was performed to compare similarities with other sequences previously registered in the GenBank® database. The strain isolated presented a sequence of 770 base pairs (bp), which was deposited in the GenBank® database with the access number PQ142877. The result of the homology search showed 100% identity with the P. tropicalis type isolate (access number NR_147864).
. Methanolic fruit and leaf extracts
The methodology proposed by De la Cruz-Ricardez et al. (2020) was used with modifications. Two hundred milliliters of 80% methanol (1:4) was added to 50 g of dry and crushed plant material from each Capsicum species (fruit or leaf). The mixture was gently stirred for 1 min and allowed to stand for 24 h at 4°C in the dark. Afterwards, the mixture was filtered, and the filtrate was placed inside an extraction hood in a water bath at 48 ± 2°C and 160 rpm until the methanol evaporated. The residue obtained was stored at 4°C until use, and prior to use, it was diluted with sterile distilled water in the same initial proportions.
. Total phenolic compounds
The concentration of MEs to be applied to each treatment was quantified based on the concentration of total phenolic compounds, following the methodology of Bautista-Rodríguez et al. (2017).
. Capsaicinoids
The capsaicinoids, capsaicin and dihydrocapsaicin, were detected and quantified in a Gas Chromatograph (GC) (Varian model 3900), coupled to a mass spectrophotometer (model Saturn 2100T) and ionization energy (EI) of 70 eV. The column was VF-5ms with a stationary phase of 5% diphenyl and 95% dimethylpolysiloxane, of 30 m × 0.25 mm, with particle size of 0.25 μm. The injection volume was 1 μl, the helium flow rate was 1.20 ml min-1, and the initial oven temperature was 228°C for 1 min, with a temperature rise in gradients of 0.20°C min-1 until reaching 230°C, maintaining constant 230°C for 7 minutes. Total content of capsaicin and of dihydrocapsaicin was calculated with the chromatograms obtained from the calibration curve of 250 to 1000 mg/l of each capsaicinoid tested (Sigma-Aldrich®). The spectra of the extracts were compared with the standards and with the library NIST MS search 2.0 (National Institute of Standards and Technology, Gaithersburg, MD, U.S.A.), and quantitative data were obtained from the integration of the areas of the peaks. The results were expressed in mg/g of dry weight.
. In vitro bioassay
The treatments included different concentrations of MEs (0.25, 0.50, 1.0, and 2.5 mg/ml) of the leaves and the immature and mature fruits of C. chinense Jacq., C. frutescens L., and C. annuum var. glabriusculum morphotypes Amashito and Garbanzo, which were added to clarified V8 medium, mixed, and placed in Petri dishes 8.5 cm in diameter, with five repetitions. A control (concentration of 0 mg/ml) was included. A 10-day-old P. tropicalis mycelium agar disk was transferred to the treated medium, which was subsequently sealed and incubated at 25°C in the dark. After 96 h, mycelial growth was measured on two orthogonal axes. The percentage of mycelial inhibition was calculated according to Lu et al. (2013) to express the result in percentage of inhibition of mycelial growth with respect to the control: (%) = ((area of mycelial growth in the control − area of mycelial growth in the treatment)/ area of mycelial growth in the control)*100. The sporangium count was performed following the methodology of Gisi et al. (1980). The results were expressed in percentage of inhibition.
. In vivo bioassay
Four-month-old Trinitario cacao pods (clone INIFAP 4) from the Los Pinos farm, Cunduacán, Tabasco, Mexico (18° 5'42.569''N and 93° 20'8.03''W), were used to evaluate the in vivo antioomycotic effectiveness of the MEs that showed greater inhibition in the in vitro tests (immature fruits of C. frutescens at 0.25 and 2.5 mg/ml, mature fruits of C. frutescens and C. chinense at 2.5 mg/ml). The pods were disinfected with 1% sodium hypochlorite and halved and placed in polyethylene bags with absorbent paper moistened with sterile distilled water. Next, 10 ml of each ME was sprayed onto the pods. P. tropicalis (agar disks with mycelium) was then inoculated at the ends of each half of the pod (Cedeño-Moreira et al., 2020). The humid bags were sealed and incubated at 25°C in the dark, with five repetitions and a control (concentration 0 mg/ml). CBP progress was recorded every 24 h, and the diameter along two orthogonal axes was measured. The measurements were stopped on day 6 because the advancement of CBP in the treatment with C. chinense ME prevented further measurements. However, for the other treatments, the progression of the disease in the later days was documented by photograph. The antioomycotic effect was calculated as a percentage of disease inhibition with respect to the control, according to Lu et al. (2013).
. Statistical analysis
Data were subjected to analysis of variance to assess the effect of the treatments on response variables, and a Tukey mean comparison test (p ≤ 0.05) was used for comparison between the treatments. Data normality was tested using a Shapiro-Wilk test (p > 0.05), and homoscedasticity of variance was tested using a Bartlett test (p > 0.05). Statistical analyses were performed using RStudio® 2024.04.2 software.
. Results
. Phenolic compounds and capsaicinoids
Phenolic compounds in the immature and mature fruits of Capsicum spp. showed significant differences between the species and maturity stages (Table 1). The mature fruits of C. chinense had the highest content (110.41 mg/g), and the immature fruits of C. annuum var. glabriusculum Amashito and Garbanzo morphotypes had the lowest contents, with 77.32 and 63.25 mg/g, respectively. In contrast, the immature fruits of C. frutescens showed a high content of phenolic compounds (104.96 mg/g). The contents of phenolic compounds in the leaves were higher than in the fruits. The highest content was observed in the leaves of C. annuum var. glabriusculum Garbanzo morphotype (215.33 mg/g), followed by C. frutescens (152.19 mg/g) and C. annuum var. glabriusculum Amashito morphotype (127.13 mg/g); there were no significant statistical differences between the latter two. The C. chinense leaves had the lowest content with 90.01 mg/g.
Table 1
Total phenolic compounds (TPC) in fruits and leaves of Capsicum spp.
The results obtained from Fisher’s LSD mean comparison test (p ≤ 0.05) for capsaicin content, according to the species and fruit maturity, are presented in Table 2. Different groupings of means are observed, indicating significant differences in the capsaicin content. The highest concentration of capsaicin was found in the mature fruits of C. chinense with 31.44 mg/g, while only 7.73 mg/g was recorded in its immature stage; the latter did not present differences from the other species. The dihydrocapsaicin levels in the fruits were lower than the capsaicin content and did not show significant differences between the species or maturity stages.
Table 2
Capsaicinoids content in fruits of Capsicum spp. studied.
. In vitro mycelial growth inhibition in P. tropicalis with methanolic extracts of Capsicum spp. fruits
The MEs of immature fruits of C. frutescens at the concentrations of 0.25 and 2.50 mg/ml and the MEs of mature fruits of C. chinense and C. frutescens at 2.50 mg/ml exhibited 52.5 to 63.9% inhibition of the in vitro mycelial growth of P. tropicalis, whereas the treatments with the remaining ME concentrations produced less than 50% inhibition (Table 3, Figure S1). Some treatments had the opposite effect, stimulating mycelial growth to levels higher than those of the control, as indicated by the negative values in Table 3.
Table 3
In‑vitro Phytophthora tropicalis mycelial growth inhibition (percentage) when methanolic extracts of Capsicum spp. fruits were used.
. In vitro mycelial growth inhibition in P. tropicalis with methanolic extracts of Capsicum spp. leaves
The MEs of the Capsicum leaves (Table 4) were less effective at inhibiting the in vitro mycelial growth of P. tropicalis than the MEs of the fruits, i.e., the C. chinense leaf MEs at a dose of 2.50 mg/ml inhibited the mycelial growth of this fungus by 27.4%.
Table 4
In‑vitro Phytophthora tropicalis mycelial growth inhibition (percentage) when methanolic extracts of Capsicum spp. leaves were used.
. In vitro inhibition of the production of P. tropicalis sporangia with Capsicum spp. methanolic extracts of fruits
A total of 90.63% of the treatments showed greater than 90% inhibition, with 2.50 mg/ml MEs of the mature fruits of the Capsicum species evaluated as the most effective (Table 5).
Table 5
In‑vitro inhibition (percentage) of the production of Phytophthora tropicalis sporangia when methanolic extracts of Capsicum spp. fruits were used.
. In vitro inhibition of the production of P. tropicalis sporangia with Capsicum spp. methanolic extracts of leaves
No treatment inhibited the production of sporangia, which was stimulated instead (Table 6), in contrast with the results of some leaf extracts that inhibited mycelial growth (Table 4).
Table 6
In‑vitro inhibition (percentage) of the production of Phytophthora tropicalis sporangia when methanolic extracts of Capsicum spp. leaves were used.
. In vivo inhibition of cocoa black pod disease with Capsicum spp. methanolic extracts
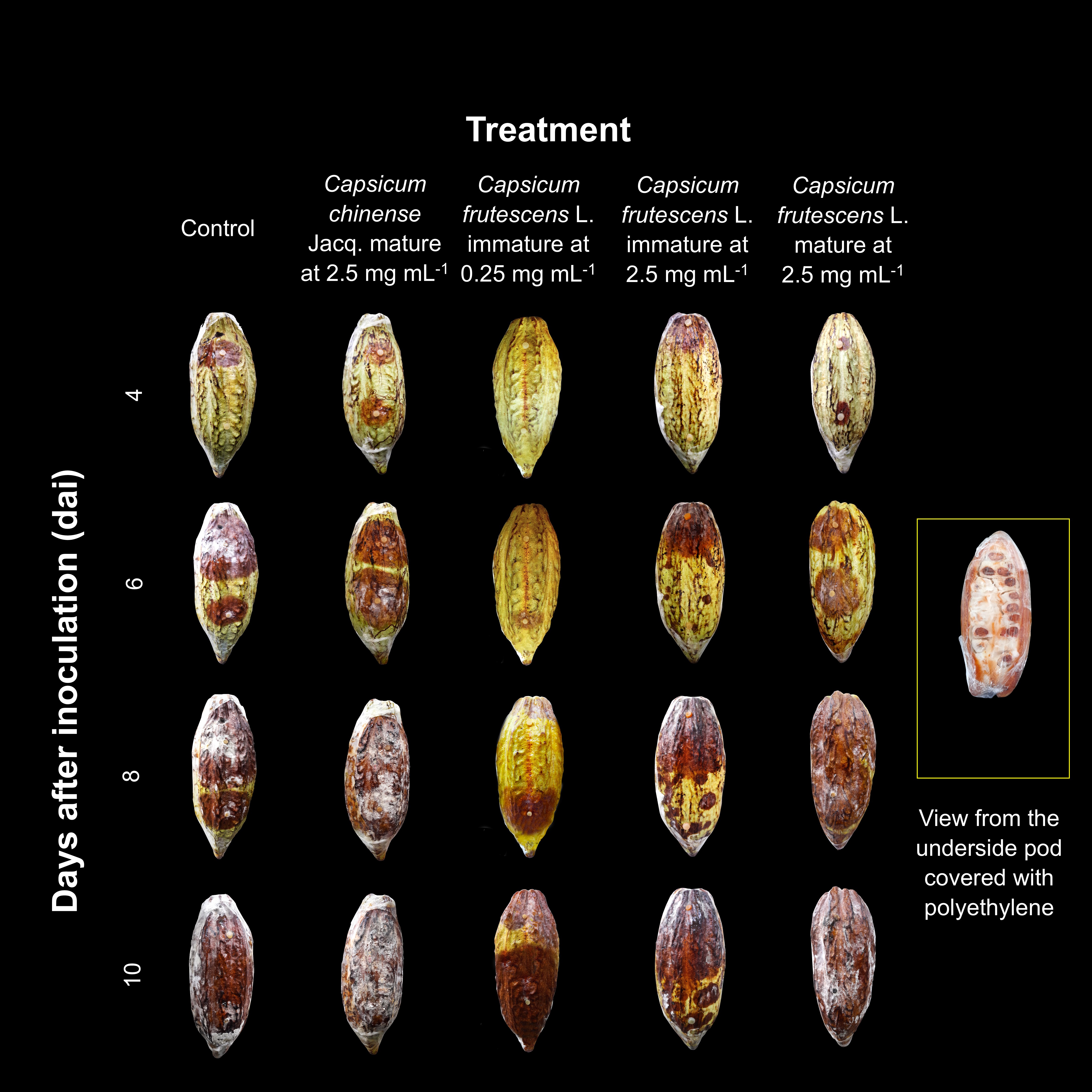
Table 7 shows the results of the inhibition of CBP by the MEs of the Capsicum fruits. The positive result shown by 0.25 mg/ml ME of the C. frutescens immature fruits was consistent with the greater than 70% inhibition of sporangium production and the 63.7% inhibition of mycelial growth (Figure 1), indicating a strong antioomycotic effect. In contrast, 2.50 mg/ml MEs of the mature C. chinense and C. frutescens fruits did not show the same trend observed in vitro (Table 3).
Table 7
In‑vivo cocoa black pod disease inhibition (percentage) at 6 days after inoculation when methanolic extracts of Capsicum spp. fruits were used.
| Species | State of maturity | Methanolic extract concentration (mg/ml) | |
|---|---|---|---|
| 0.25 | 2.50 | ||
| C. chinense Jacq. | Mature | — | -11.50c |
| C. frutescens L. | Immature | 88.60a | 36b |
| Mature | — | -0.90c | |
| Control | 0c | 0c | |
. Discussion
The variations in mycelial growth inhibition between extracts of the different species may be related to the variation in the types and proportions of the secondary metabolites present in the evaluated extracts (Aguirre-Hernández et al., 2017; Cervantes-Hernández et al., 2019; De la Cruz-Ricardez et al., 2024; Rodriguez-Maturino et al., 2015) and are probably associated with the way in which these extracts act on the cell membrane of the phytopathogen. Some authors have noted that, on the one hand, at the cell membrane level, plant extracts can cause membrane rupture, protein denaturation, lysis, and intracellular osmotic imbalance, among other effects (Lu et al., 2013; Madiha et al., 2018). On the other hand, secondary metabolites present hydroxyl free radicals, which are highly reactive and can cause peroxidation of cell membrane lipids. In addition, the carbon double bonds (C=C) are highly electronegative and interfere with the transfer of electrons during biological processes, giving these metabolites antimicrobial effects (Lu et al., 2013). The ME of the immature C. frutescens fruits showed the highest content of phenolic compounds among the species studied. Recent studies on mature fruits of this species of chili have shown that the pericarp contains compounds with bioactive properties, such as isopimaric acid, Pedalitin, Xi-8-acetonyldihydrosanguinarine, Pratenol B, and Uralenneoside (Cervantes-Hernández et al., 2019). In Capsicum chinense fruits, the complex mixture of capsaicinoids, phenolic acids, and other minor bioactive compounds seem to contribute to the antimicrobial activity of pepper extracts (Vuerich et al., 2023). The metabolomic profile should change in immature fruits; therefore, research in this regard should be considered in future studies.
Among the Capsicum species evaluated whose MEs inhibited the growth of P. tropicalis, C. chinense has previously shown antifungal activity against fungi of agricultural importance, such as Alternaria alternata (Rodriguez-Maturino et al., 2015), and C. annuum has shown activity against P. megakarya (Simo et al., 2019). Among the Capsicum species present in Tabasco, Mexico, MEs from the immature fruits of C. annuum var. glabriusculum and C. frutescens have been shown to inhibit the growth of M. roreri in vitro (De la Cruz-Ricardez et al., 2020). The results obtained in the present investigation revealed that 2.50 mg/ml ME from the C. frutescens fruits also inhibited the growth of P. tropicalis. The results of both investigations are promising since they may facilitate the development of effective and sustainable control strategies to combat moniliasis and cocoa black pod.
When comparing the inhibitions achieved with the MEs from the leaves and fruits of Capsicum spp., it was observed that the MEs from fruits were more effective in inhibiting P. tropicalis. The results of the analysis of phenolic compounds in the fruits and leaves (Table 1) showed that the leaves had a higher phenolic compound content than the fruits. This seemed to indicate that it was more likely to find more phenolic compounds with antioomycotic activity in the leaves than in the fruits. However, the lower inhibition induced by the leaf extracts suggest that leaves contain a lower concentration of compounds with antioomycotic activity, or it is possible that secondary metabolites in the leaves are in inadequate proportions, making their antioomycotic activity ineffective. A lower inhibition effect of leaf extracts has already been reported in previous research. Ameziane et al. (2007) studied the inhibition of mycelial growth of Geotrichum candidum using extracts from various plant species. They found that, when using leaf extracts of Lippia citriodora, mycelial growth was not inhibited; on the contrary, it was stimulated, showing a 2% increase with respect to the control. This contrasted with the other treatments in which they used extracts of seeds, bark, or a combination of stem and leaves, reaching 100% inhibitions. A similar effect was observed by Askarne et al. (2012) using different plant extracts on Penicillium italicus.
The MEs were more effective in inhibiting sporangium formation than mycelial growth. This result underscores the ability of Capsicum spp. to indirectly reduce the formation of infectious propagules (zoospores) of P. tropicalis. The effectiveness of these extracts is possibly attributable both to the action of aldehydes and to the complexes formed between phenolic compounds and soluble proteins of the MEs, which exert a degenerative action on the cell wall (Nielsen et al., 2006; Rodriguez-Maturino et al., 2015). These effects not only destabilize the cell wall of the phytopathogen but also interfere with critical pathways for the synthesis of RxLR effector proteins (He et al., 2024) and with the restructuring of cellular material, including the production of sporangia. A comparison of the results obtained in the present investigation with those reported by De la Cruz-Ricardez et al. (2020) revealed that with 2.50 mg/ml ME of the immature fruits of the studied species, especially the Amashito and Garbanzo morphotypes, significantly reduced the number of M. roreri and P. tropicalis propagules, indicating the potential for the development of effective biotechnologies that combat both diseases.
In contrast, the compounds present in the leaves may not be sufficient to inhibit mycelia and sporangia simultaneously. Another explanation is that P. tropicalis may have responded to the stress induced by the MEs, increasing the production of sporangia as a survival mechanism. The results highlight the complexity of the interactions between the secondary metabolites of Capsicum and P. tropicalis since a single treatment can have different effects on the different phases of the latter’s life cycle. This stimulation of sporangium production from Capsicum spp. by MEs has not been reported in the literature, opening new possibilities for the use of leaf extracts in the induction of sporangia.
The Capsicum spp. MEs did not prevent pod infection or impeded disease progression, but there was a slowing effect on the progression of necrosis and sporulation. The observations suggest that the appearance and development of white mycelium was delayed by the treatments with the immature and mature C. frutescens at 2.50 mg/ml and the immature C. frutescens at 0.25 mg/ml (Figure 1), indicating that MEs could interfere with the cocoa black pod cycle in field conditions. In particular, the treatment with the immature C. frutescens fruit MEs at 0.25 mg/ml contributed to weak surface mycelium development until day 10 after infection, suggesting its potential to reduce the vertical spread of black spot in cocoa plantations. The delay in the appearance of the mycelium has important implications since the infective potential of P. tropicalis is reduced. In the mycelium, hyphae form sporangia and chlamydospores, which contain infectious propagules (zoospores). Consequently, future outbreaks of the disease in plantations would have a progressively lower incidence. This behavior differed from that reported by Simo et al. (2019), who used aqueous and ethanolic Capsicum extracts, where all the concentrations evaluated delayed the appearance of P. megakarya symptoms in cocoa pods, and the aqueous extracts significantly reduced the incidence index. Similarly, Nana et al. (2015) reported a positive correlation between the inhibition of necrosis in cocoa pods infected with P. megakarya and the concentration of Zanthoxylum zanthoxyloides and Syzygium aromaticum essential oils.
However, in our study, the difficulty in stopping the disease from advancing could also be attributed to the humid conditions in the chamber where the pods were placed, which favored their development. As shown in Figure 1, the effectiveness of the treatments decreased over time, possibly due to several factors, including the half-life of the bioactive molecules that act against the phytopathogen, the volatility of the extracts, and the possible adaptation of P. tropicalis to the treatments.
. Conclusion
In conclusion, we found that the MEs from Capsicum fruits showed the best antioomycotic effect in vitro compared to the MEs from leaves. The greatest inhibition of mycelial growth was achieved with the MEs of immature C. frutescens fruits at 0.25 and 2.50 mg/ml, followed by those of mature C. chinense and C. frutescens fruits at 2.50 mg/ml. In addition, more than 90% of the treatments with the fruit MEs efficiently inhibited the production of sporangia. The in vivo test revealed that 0.25 mg/ml ME of immature C. frutescens fruits was the most effective at slowing the progression of cocoa black pod. This treatment delayed the appearance of aerial mycelia on the pods for 10 days, highlighting the potential of this treatment for the development of control strategies against P. tropicalis in cacao.
