. Introduction
The growing human population requires adequate food sources. With a projected global population of 9.2 billion by 2050, food demand is expected to rise significantly. Population growth, climate change, and many new, developing, and invasive plant diseases threaten global food security (Anderson et al., 2004). Numerous plant pests and pathogens threaten global food security, as they lead to 20% to 40% reduction in crop yields worldwide each year (Savary et al., 2019). Climate change worsens the situation and leads to alterations in the assembly of pathogens, further exacerbating the issue (Chaloner et al., 2021). Managing plant diseases effectively is essential for ensuring a sustainable food supply, meeting global demand while maintaining and improving crop quality and yield.
Plant disease management can be approached environmentally benignly and often complementarily by creating plant types with intrinsic disease resistance (Sánchez-Martín & Keller, 2019). Both conventional and molecular technologies are used in plant breeding today to produce crops that are safe, nutrient-dense, productive, and free of disease (Kaiser et al., 2020; Tyagi et al., 2021). In recent decades, conventional and mutation breeding as well as transgenic technology have all demonstrated potential. However, they have various drawbacks, including high labour costs, inherent genetic variation within plant populations, and transfer of unwanted genes or traits along with selected resistance genes (Ahmad et al., 2020; Gao, 2018).
In response to these limitations, innovative breeding technologies, including meganucleases (MNs), zinc finger nucleases (ZFNs), transcription activator-like (TAL) effector nucleases (TALENs), and clustered regularly interspaced palindromic repeats (CRISPR) and CRISPR-associated proteins (Cas), with the most commonly used Cas9 endonuclease, have been created (Nekrasov et al., 2013; Shan et al., 2013). These innovative breeding techniques enable specific genetic modifications of plants’ single or multiple gene targets (Borrelli et al., 2018). Target DNA is cleaved by sequence-specific nucleases, such as MNs, ZFNs, TALENs, and Cas proteins. DNA double-strand breaks are repaired by homology-directed repair (HDR) or non-homologous end-joining (NHEJ), typical host cell repair mechanisms; nevertheless, minor indels (insertions/deletions) might happen inside the target area (Langner et al., 2018).
This comprehensive review will provide a careful overview of the intricate mechanisms involved in plant defence during biotic stress action. Subsequently, the review explores the cutting-edge tools and methodologies that enable precise genome editing, facilitating the creation of targeted genetic variations by inducing double-strand breaks in DNA and facilitating large-scale genome engineering. Additionally, the review investigates the application of genome editing in pathogens to point their virulence genes, shedding light on the complex dynamics of the host-pathogen relationship and paving the way for developing more effective strategies to combat pathogens and curtail the spread of diseases.
Moreover, we will scrutinise how host plant functional genomics can harness the CRISPR technique to identify resistance (R) or susceptibility (S) genes and modify host genomes to enhance resistance breeding. Furthermore, our discussion will explore the “gain-of-function” of host genes. Lastly, we will meticulously examine documented genome editing-mediated host resistance augmentation instances against various diseases and pests.
. Mechanism of the Plant Defence System
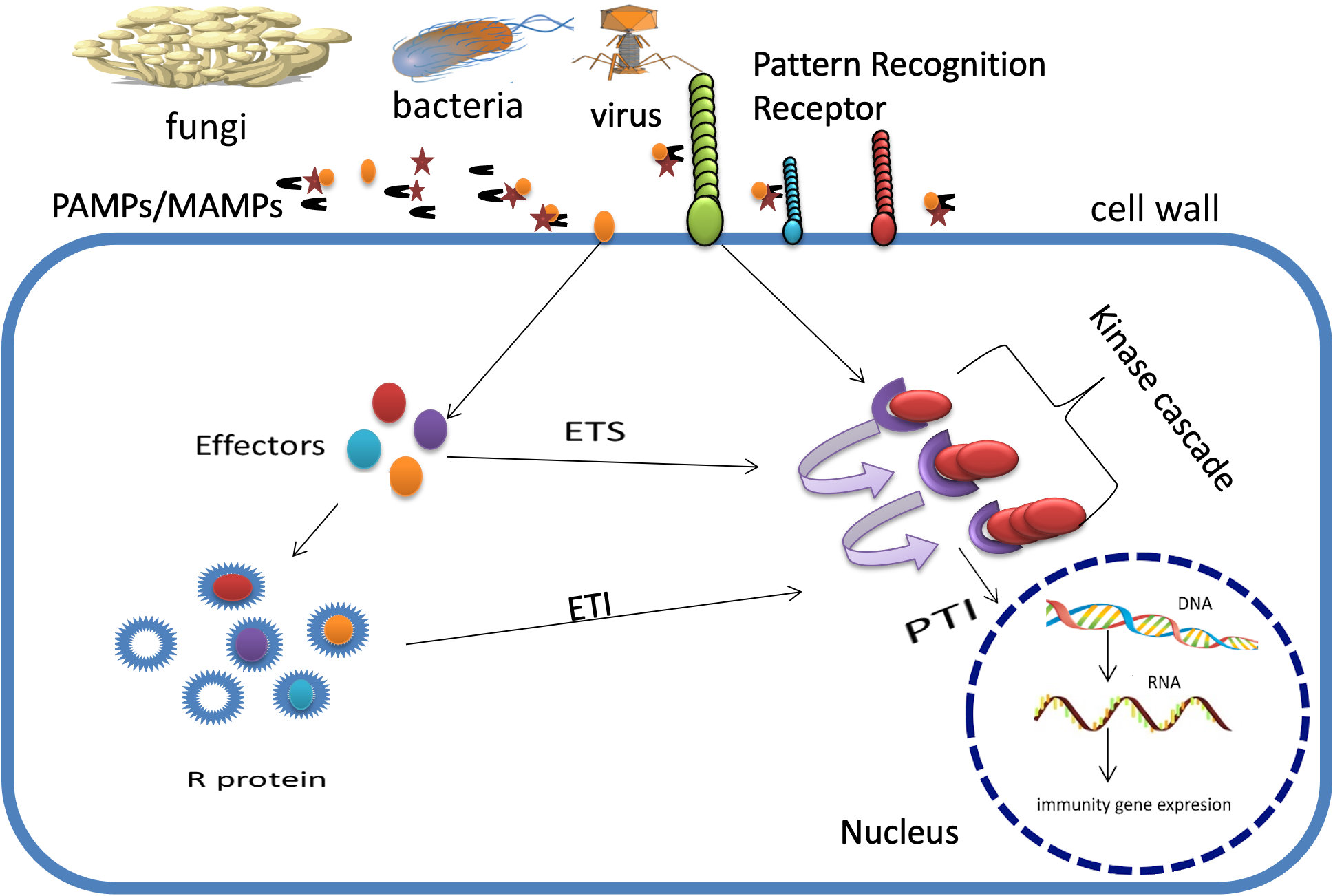
Developing an effective strategy for managing plant diseases requires dissecting the relationship between the pathogen and the host plant and identifying the components of plant defence response (Veillet et al., 2020). To repel invasive infections, plants rely on a complex defence system (Langner et al., 2018). The plant’s sophisticated two-layered resistance system can detect pathogens or pathogen-secreted chemicals. This immune system consists of PTI (pattern-triggered immunity) and ETI (effector-triggered immunity), and it can detect these threats either within the plant cell or on the cell’s periphery (Yuan et al., 2021) (Figure 1). Immune response pathways are buffered by signalling pathways of defence hormones, such as the ethylene pathway for chewing pests and necrotrophic pathogens and the salicylic acid (SA) pathway for biotrophic and hemibiotrophic pathogens (Robert-Seilaniantz et al., 2011). Plants can balance growth and defence through the intricate interplay of phytohormone signalling pathways (Berens et al., 2017). A comprehensive analysis was recently conducted on the components of plant resistance systems, their interactions and controls, and the latest developments in identifying pattern recognition receptors (PRRs) and nucleotide-binding leucine-rich repeats (NLRs) in the plant kingdom (Ngou et al., 2022). There is some protection against plant disease from using pesticides and cultural and horticultural methods. However, due to pathogen evolution, pesticides are rapidly losing their efficacy and causing catastrophic environmental devastation. As a result, they are generally subject to tight regulations aimed at minimising undesired side effects (Esse et al., 2020; Tyagi et al., 2021).
. Unlocking the potential of genome editing with tools for creating knock-outs, inducing genetic variation, and making precise modifications
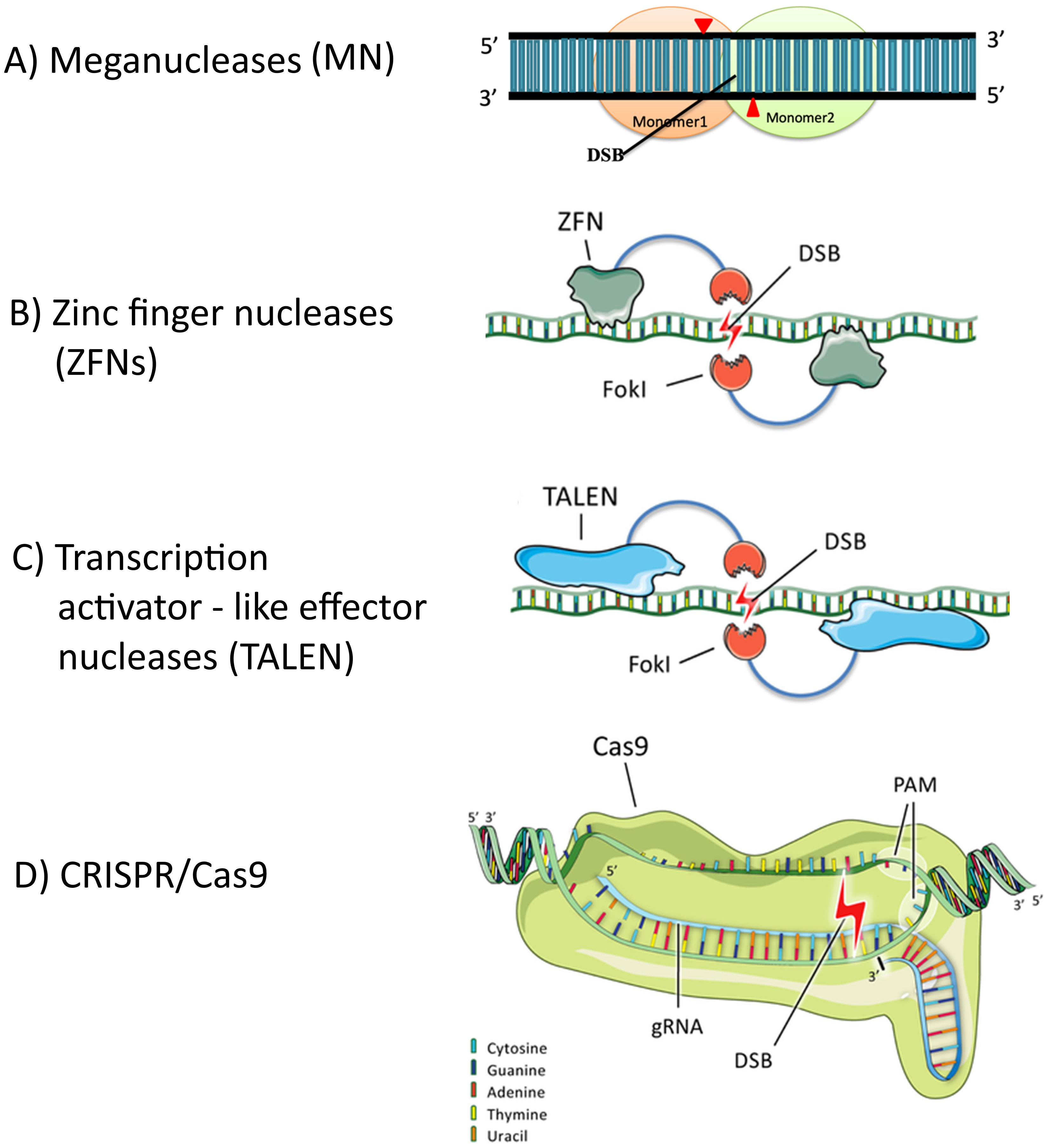
The novel breeding technology (NBT) known as genome editing (GE) has rapidly earned popularity. Unlike genetically modified crops, most genome editing-derived cultivars are not subject to extra regulations in many countries and do not contain any foreign DNA (Menz et al., 2020). According to Chandrasegaran and Carroll (2016), GE harnesses the power of programmable nucleases to target DNA sequences precisely, introducing specific and impactful modifications to the genome. These nucleases include MNs, ZFNs, TALENs, and clustered regularly interspaced short palindromic repeat (CRISPR)-associated nucleases (Cas) (Figure 2). Because of its versatility, amenability to multiplexing, and ease of use, the CRISPR/Cas technology has surpassed MNs, ZFNs, and TALENs (Gao, 2021). CRISPR/Cas, ZFNs, and TALENs produce targeted genetic variants through the uncontrolled and spontaneous production of indels.
The utilisation of various online tools for multiplexing, the creation of single-guide RNA (sgRNA), and the implementation of cutting-edge cloning techniques, such as GoldenBraid, BioBrick, and Golden Gate, represent significant progress in the CRISPR/Cas technology. Furthermore, the development of CRISPR/Cas constructions, advanced transformation methods including those mediated by Agrobacterium, and the adoption of DNA-free protoplasts for Cas9-guide RNA (gRNA) ribonucleoprotein (RNP) complexes collectively contribute to the enhancement of gene editing abilities. The incorporation of diverse Cas9 variants, such as Cas9-NG/XNG-Cas9 and PAM-free Cas12a (Cpf1), in conjunction with innovative approaches like base/prime editing utilising the Target-Activation-Induced Cytidine Deaminase (AID) technology and homologous recombination (HR)-based gene knock-in (HKI) mediated by geminivirus replicon, have significantly expanded the toolkit for exact genome modification (Naeem et al., 2024).
. Meganucleases
Meganucleases improved genome editing efficiency by recognising lengthy regions of 14–40 bp DNA, but they were limited by two significant issues (Langner et al., 2018). Firstly, it is unlikely to find a meganuclease that targets a specific location because each has a unique recognition sequence. Secondly, when double-strand breaks occur, most of them are repaired using the error-prone NHEJ DNA repair mechanism, which makes it challenging to incorporate the preferred DNA template at the break sites due to the possibility of random insertions or deletions of DNA segments (Adli, 2018).
. Zinc finger nucleases (ZFNs)
Identifying eukaryotic zinc finger proteins directed in a new phase of genome editing research. Zinc fingers are sequence-specific tiny protein motifs regulated by zinc ions, able to bind to DNA. ZFNs have a reduced specificity since each zinc finger module can target any 18–21 bp genomic region because it can recognise a 3bp DNA sequence (Swarthout et al., 2011). Specific changes were made to zinc finger proteins, enabling the creation of programmable nucleases with the DNA cleavage domain of the FokI endonuclease. FokI can form a homodimer using two distinct zinc finger modules targeting two nearby sites. This results in DNA strand breaks at the target sites (Adli, 2018; Chen et al., 2014).
. Transcription activator-like effector (TALE)
Xanthomonas produce transcription activator-like effector (TALE) proteins that can distinguish between one and three bases (Adli, 2018). Like zinc fingers, TALENs, which are chimeric fusions of the FokI DNA cleavage domain to a set of TALE modules, act as efficient programmable nucleases. While ZFNs and TALENs are valuable engineered nucleases that can improve genome editing efficacy, the challenges associated with their cloning and protein engineering have limited their widespread adoption by researchers (Adli, 2018; Chen et al., 2014).
. CRISPR systems: the CRISPR/Cas9 tool
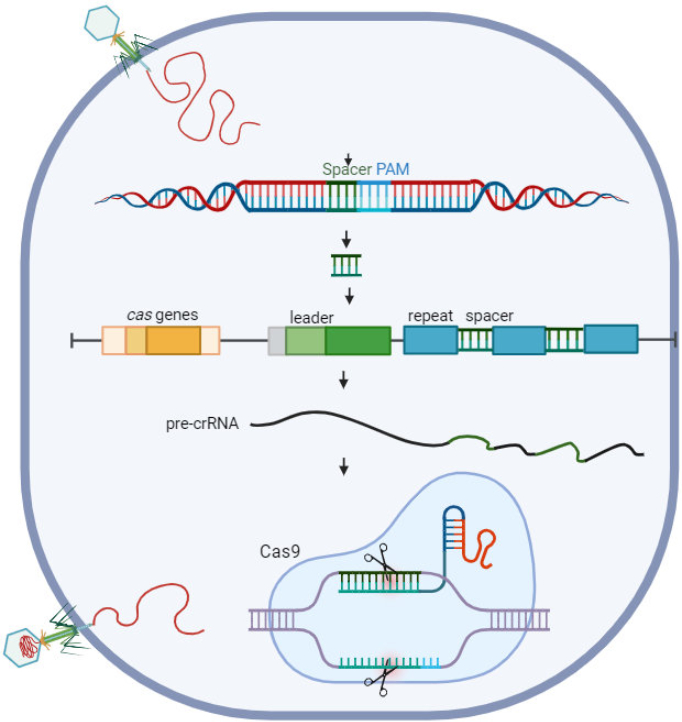
CRISPR, which stands for clustered regularly interspaced short palindromic repeats, was initially recognised as a unique prokaryotic DNA repeat element. It was later identified as the bacterial immune system, and subsequently utilised as a potent reprogrammable gene-targeting tool (Knott & Doudna, 2018). CRISPR/Cas9 is a powerful and exact genome-editing technique that facilitates gene function analysis and allows the production of targeted mutants with specific traits (Alamillo et al., 2023). The growing number of recent papers on plant genome editing utilising the CRISPR/Cas9 system suggests that this method is practicable due to its increased success rate and ease of usage.
The CRISPR gene editing method uses an RNA-guided endonuclease protein to create specific double-strand breaks DSBs (Knott & Doudna, 2018). Various genome editing tools, such as MNs, ZFNs, TALENs, and CRISPR/Cas9, can cause DSBs, allowing the disruption of desired genes via NHEJ or the insertion of desired genes via HDR. However, the CRISPR/Cas9 technology is the preferred method for researchers in various fields due to its comfort of use and high editing efficiency for genome targeting purposes (Adli, 2018).
RNA hybrid made up of a trans-activating crRNA (tracrRNA) and a CRISPR RNA (crRNA) directs the binding and cleavage activities of Cas9. Combining tracrRNA with crRNA creates a sgRNA (Jinek et al., 2012). A brief (20–23 bp) sgRNA segment is changed to target a new sequence once complementarity identifies the target nucleic acid. The Cas9 HNH and RuvC domains cleave the target DNA’s complementary and non-complementary strands, resulting primarily in blunt-ended DSBs. Subsequently, the host cellular component repairs the DSB (Makarova et al., 2011; Shao et al., 2016).
Despite discovering other Cas9 orthologs (Molla et al., 2020a), Streptococcus pyogenes (SpCas9) Cas9 is the most often employed nuclease for various genome editing research because of its high effectiveness and low requirements for the NGG PAM sequence. A protospacer adjacent motif, or PAM for short, is a conserved sequence of 2–5 base pairs that is needed by Cas9 to identify, bind, and cut target genomic locations (Jiang & Doudna, 2017). Different species’ Cas9 might require different PAMs (Molla et al., 2020a). For Cas9, PAM must be situated directly downstream of the target sequence. While most researchers believe that Cas9 cuts DNA three base pairs upstream of the PAM on both strands (Jiang & Doudna, 2017), a rising body of recent data suggests that Cas9 frequently mediates cleavage at 4-, 5-, and 6-bp upstream in the non-complementary strand (Molla & Yang, 2020b) (Figure 3). Cas12 cleaves at the PAM distal location, resulting in a staggered DNA double-stranded break (Zetsche et al., 2015). Cas13 cleaves single-stranded RNA (Abudayyeh et al., 2016).
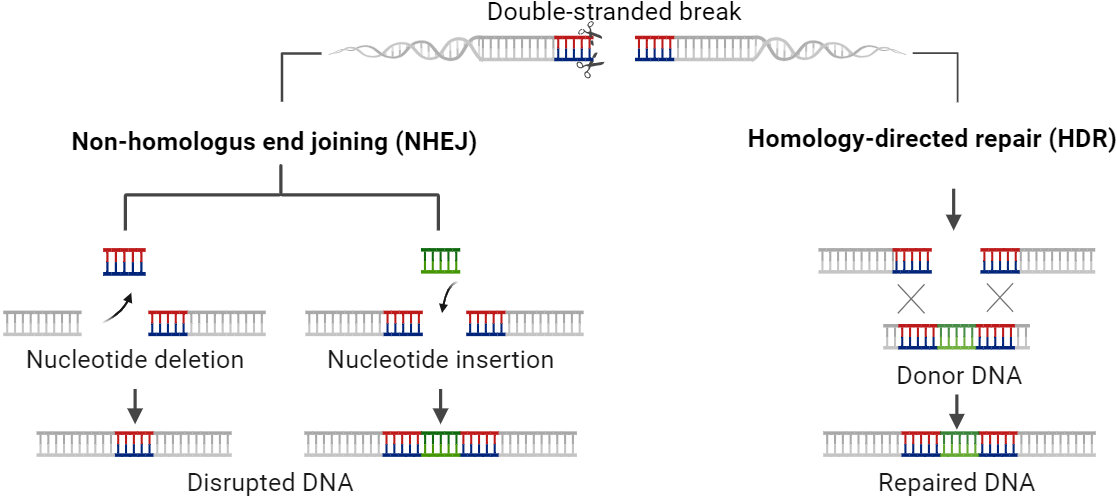
The two main mechanisms for repairing genomic double-strand breaks (DSBs) produced by site-specific nucleases are HDR and NHEJ (Figure 4). Higher plants use primarily NHEJ, which is error-prone and results in tiny insertions or deletions (indels). Indel-induced frameshift mutations offer efficient gene knockout. With the development of CRISPR/Cas9 tools, it is now easier to knock off a gene or produce targeted genetic variation (Jinek et al., Jinek et al., 2012). However, homology-directed repair (HDR) in higher plants is not as prevalent as in other organisms. It is utilised for precise adjustments, often requiring an extra source of donor templates containing the desired modification.
. Strategies for engineering disease resistance with CRISPR technologies
Recent advancements in the CRISPR technology have enabled researchers to create various CRISPR variants with multiple applications. This section will highlight the successful use of the CRISPR technology in engineering plants resistant to various diseases (Figure 5).
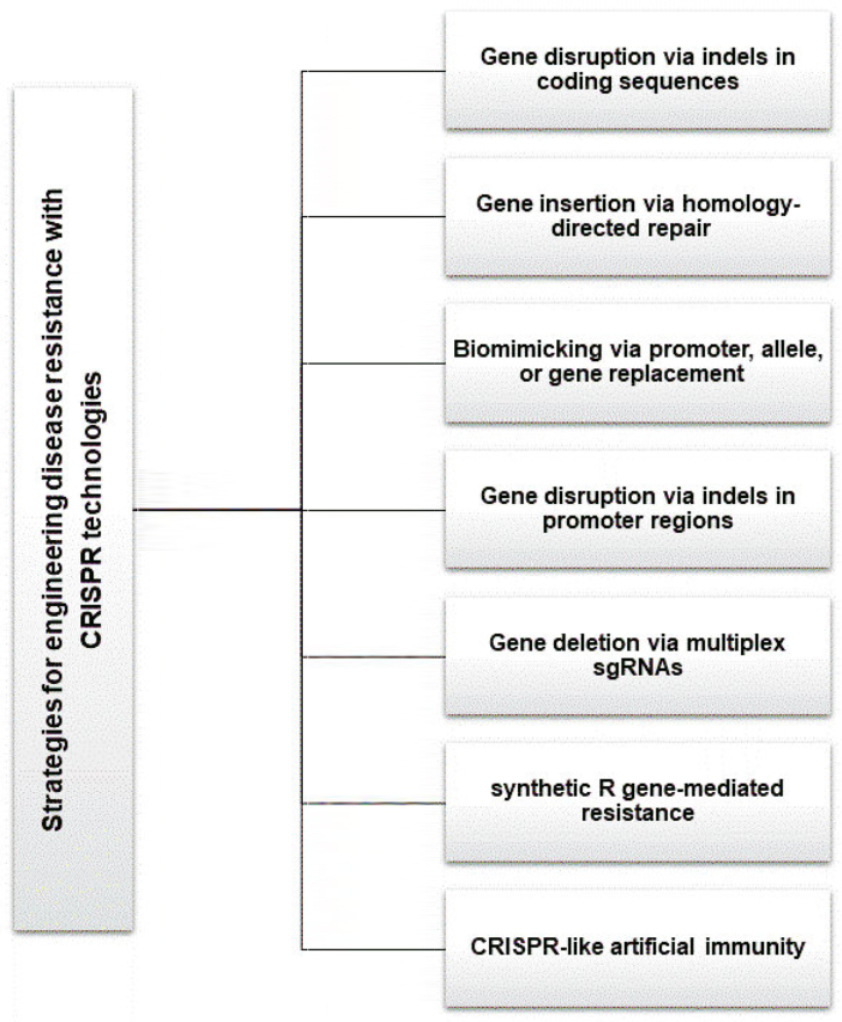
. Gene disruption via indels in coding sequences
Numerous prevalent applications of the CRISPR/Cas9 system involve leveraging the error-prone nature of the NHEJ DNA-repair machinery within cells. This process results in insertion or deletion of one or more nucleotides (indel) at the specific location guided by sgRNA. Consequently, a frameshift mutation occurs, disrupting gene transcription. This approach has effectively introduced favourable characteristics into a broad spectrum of crops, including key cereals like wheat and rice (Chen et al., 2014). By manipulating plant’s susceptibility (S) genes, which alters plant-pathogen interaction and reduces the pathogen’s ability to thrive on the host plant, this method has been employed to develop resistance within the disease management framework (Zaidi et al., 2018). The use of CRISPR/Cas9 to introduce mutations in the eukaryotic translation initiation factor 4E proteins (eIF4Es) has been particularly notable. This approach has been successfully employed in Arabidopsis and cucumber (Cucumis sativus) to confer resistance against multiple RNA viruses (Chandrasekaran et al., 2016; Pyott et al., 2016).
Susceptibility (S) genes in host plants promote pathogen invasion and colonisation (Li et al., 2014; Schie & Takken, 2014). Opposed to R genes, manipulating such S genes imposes long-term resistance in the field (Langner et al., 2018). The genes labelled as S genes can be divided into three groups: the first encompasses genes that assist in host recognition by pathogens and their penetration, such as IRG1 in Medicago and MLO in barley and other plants. The second includes genes that facilitate pathogen nutrition and promote the establishment of pathogens, such as SWEET genes in rice and eIF4 genes for viral infection in many plants. The third gene group operates as a negative regulator for host defence responses, with examples like S3H in Arabidopsis and PM4 in tomato (Schie & Takken, 2014). According to recent research, the CRISPR/Cas technique targeting a host gene is becoming increasingly popular for treating disease and pest resistance (Table 1).
Table 1
CRISPR/Cas approaches increasing pathogen and pest resistance by targeting a host gene reviewed by Schenke and Cai (2020) extended in this work.
| Plant species | Target gene | Resistance to pathogen/pest | Comment | Reference |
|---|---|---|---|---|
| Arabidopsis | AtEIF(iso)4E | Turnip mosaic virus (TMV) | Cas9 | Pyott et al., 2016 |
| AtIAN9 | Pseudomonas syringae (bacterial speck) | Cas9 | Wang et al., 2018 | |
| At2OGO (DMR6-like) | Fusarium graminearum (head blight) | Cas9 | Low et al., 2020 | |
| AtErf019 | Phytophthora parasitica | Cas9 | Lu et al., 2020a | |
| AtCYP71A13 | Botrytis cinerea (grey mold) | Cas9 | Ferrari et al., 2007 | |
| Apple | MpDIPM-1/2/4 | Erwinia amylovora (fire blight) | Cas9 RNP | Malnoy et al., 2016 |
| Barley | HvMorc1 | Blumeria graminis (Bgh) / Fusarium graminearum | Cas9 | Kumar et al., 2018 |
| Cacao | TcNPR3 | Phytophthora tropicalis | Cas9 | Fister et al., 2018 |
| Cassava | MenCBP-1/2 | Cassava brown streak virus (CBSV) | Cas9 | Gomez et al., 2019 |
| Cotton | Gh14-3-3d | Verticillium dahliae | Cas9 | Zhang et al., 2018b |
| Cucumber | CseIF4E | Several viral diseases: CVYV, ZYMV, PRSV-W | Cas9 | Chandrasekaran et al., 2016 |
| Duncan grapefruit | CsLob1 | Xanthomonas citri | Cas9 promoter | Jia et al., 2016 |
| Grape | VvMLO-7 | Erysiphe necator (powdery mildew) | Cas9 RNP | Malnoy et al., 2016 |
| VvWRKY52 | Botrytis cinerea (grey mold) | Cas9 | Wang et al., 2018 | |
| Maize | ZmNLB18 (WAK) | Setosphaeria turcica (Northern Leaf Blight) | Cas9 HDR | USDA, 2017 |
| Nicotiana benthamiana | CLC-Nb1a/b | Reduced PVY intracellular replication | Cas9 | Sun et al., 2018a |
| Oilseed rape | BnWRKY70 | Sclerotinia sclerotiorum (stem rot) | Cas9 | Sun et al., 2018b |
| BnCRT1a | Verticillium longisporum (stem striping) | Cas9 | Pröbsting et al., 2020 | |
| Rice | OsSWEET11/14 | Xanthomonas oryzae (bacterial blight) | Cas9 promoter | Jiang et al., 2013 |
| OsSWEET13 | Xanthomonas oryzae | Cas9 | Zhou et al., 2015 | |
| OsERF922 | Magnaporthe oryzae (rice blast) | Cas9 | Wang et al., 2018 | |
| OsBsr-d1 | Magnaporthe oryzae | Cas9 | Li et al., 2017 | |
| OsSec3A | Magnaporthe oryzae | Cas9 | Ma et al., 2018 | |
| OseIF4G | Rice tungro spherical virus (RTSV) | Cas9 | Macovei et al., 2018 | |
| OsCYP71A1 | Insects: brown planthopper and striped stem borer | Cas9 | Lu et al., 2018 | |
| OsPi21 | Magnaporthe oryzae | Cas9 multiplex | Li et al., 2019b | |
| OsXa13 | Xanthomonas oryzae | Cas9 promoter | Li et al., 2020a | |
| Os8N3 | Xanthomonas oryzae | Cas9 | Kim et al., 2019 | |
| OsCul3a | Xanthomonas oryzae / Magnaporthe oryzae | Cas9 | Gao et al., 2020; Liu et al., 2017 | |
| OsHPP04 | Meloidogyne graminicola | CRISPR/Cas9 | Huang et al., 2023 | |
| Soybean | GmF3H1/2, FNSII-1 | Soybean mosaic virus (SMV) | Cas9 | Zhang et al., 2020b |
| GmTCP19L | P. sojae | CRISPR/Cas9 | Fan et al., 2022 | |
| Tomato | SlDMR6 | P. syringae, Phytophthora capsici, Xanthomonas spp. | Cas9 | Thomazella et al., 2021 |
| SlMLO1 | Oidium neolycopersici (powdery mildew) | Cas9 | Nekrasov et al., 2017 | |
| SlPMR4 | Oidium neolycopersici (powdery mildew) | Cas9 | Santillán Martínez et al., 2020 | |
| Watermelon | Clpsk1 | Fusarium oxysporum | Cas9 | Zhang et al., 2020a |
| Wheat | TaMLO | Blumeria graminis (powdery mildew) | Cas9 | Wang et al., 2014 |
| TaEDR1 | Blumeria graminis | Cas9 | Zhang et al., 2017 | |
| TaNFXL1 | Fusarium graminearum (head blight) | Cas9 | Brauer et al., 2020 |
S gene editing can offer a broader and more resilient resistance range since changing S genes disrupts compatibility. The most researched S gene is mildew locus O (MLO), found in numerous plant species. It encodes a protein tethered to the membrane and facilitates the entry of powdery mildew fungus into epidermal cells (Büschges et al., 1997). Several plant species lost their vulnerability to MLO when it was edited using CRISPR/Cas (Nekrasov et al., 2017; Wang et al., 2014). Editing effector binding elements (EBEs) and sequences in the promoters of rice SWEET genes have been linked to the development of Xanthomonas oryzae pv. oryzae (Xoo) resistance, according to several studies (Duy et al., 2021; Kim et al., 2019; Zeng et al., 2020). The translation of viral RNA requires the presence of plant translation initiation factors (eIF) 4E and 4G (Wang & Krishnaswamy, 2012). Recent research has shown that CRISPR mutants of the eIF4E and eIF4G genes demonstrated resistance (Macovei et al., 2018; Mushtaq et al., 2020). S genes are a specific group of genes that produce proteins responsible for negatively regulating the plant’s defence mechanisms. These proteins help to modulate and balance the plant’s immune responses, preventing overreactions that could be harmful to the plant itself. For instance, the cotton gene Gh14-3-3d engages in BR signalling and interferes with Verticillium dahlie resistance (Gao et al., 2013).
According to Schie and Takken (2014), the majority of S genes that have been identified are monogenic; numerous genes can encode susceptibility factors, and CRISPR/Cas multiplexing offers an alluring way to target them all at once. Comparison of transcriptomes or proteomes after host-pathogen interaction may help identify more unique S gene targets in the plant system. Numerous S genes are crucial for the growth and development of crops. Crop productivity and growth would suffer if these S genes were functionally knocked out. For instance, while conferring disease resistance, the null mutants of OsSWEET14/Os11N3 (Antony et al., 2010) and mlo (Li et al., 2022) also showed growth penalties and yield losses.
. Gene disruption via indels in promoter regions
There are two strategies for utilising CRISPR-mediated promoter editing in genetic modification. The first involves disrupting the promoter sequence to halt gene production entirely. The second approach focuses on disrupting an effector-binding site to prevent a pathogen effector from binding to the promoter, thereby reducing the plant’s susceptibility to pathogens. By modifying the rice sugar transporter OsSWEET14 gene’s promoter, the latter method has been successfully employed to confer blight resistance by severing the gene’s interaction with an effector from a bacterial blight pathogen (Blanvillain-Baufumé et al., 2017; Xu et al., 2019). CRISPR, a revolutionary gene-editing technology, has demonstrated its capability to not only edit promoters but also modify gene regulation by targeting regions upstream of open reading frames (ORFs) and editing cis-regulatory elements (Si et al., 2020).
. Gene deletion via multiplex sgRNAs
Multiple sgRNAs can be utilised in CRISPR systems to introduce numerous DSBs at specific target genomic regions. For example, two sgRNAs binding to the gene of interest before its start codon and after its finish codon will result in DSBs at the corresponding places. Before the NHEJ mechanism can fix the breaks, the DSBs may lead to the loss of the DNA fragment that encodes the desired protein. This method can be employed to delete individual genes and substantial chromosomal segments (Cai et al., 2018; Zhou et al., 2014). Developing a logically constructed Cas9, known as SpCas9-NG, has enabled the identification of NG PAMs with a relaxed stringency compared to the usual NGG PAMs. This advancement further facilitates the process (Nishimasu et al., 2018). In S gene clusters, when many S genes are located on contiguous chromosomes (Ashfield, 2012), removing the chromosomal region will likely result in long-term resistance to the infectious agents.
. Gene insertion via homology-directed repair
Altering the significant and multifunctional proteins that are synthesised by S genes may lead to unforeseen consequences for both plant health and agricultural productivity. When gene disruption is not the preferred method, other options, like editing the cis-regulatory elements and promoters, can be utilised to modify gene expression. However, in cases where the plant-pathogen interaction is not comprehensively captured, and the S genes have not been thoroughly studied, resistance (R) genes against pathogens are often necessary. In such situations, R genes can be inserted using the CRISPR toolbox. The Cas9 protein and the sgRNAs are combined with a delivery fragment that includes an R gene, flanked by a sequence matching the ends of a DSB. The R gene is placed between the two DSB sites with the help of this cassette using HDR. This method has been used to insert one or more genes at precise positions in the genome (Van Vu et al., 2019).
Still, a significant obstacle to HDR’s frequent deployment is its low frequency in higher plants (Molla et al., 2020a). A variety of strategies have been investigated with varying degrees of success to improve HDR efficiency in plants, including viral replicons (Čermák et al., 2015), bacterial retrons (Sharon et al., 2018), transcript-templated HDR (Li et al., 2019a), tandem repeat HDR (Lu et al., 2020b), and chimeric guide RNA (Butt et al., 2017). Ali et al. (2020) developed a Cas9 and VirD2 relaxase fusion to create targeted DNA double-strand breaks. This allows the repair template to be positioned close to the DSBs, aiding in HDR.
. Biomimicking via promoter, allele, or gene replacement
When specific changes are made to the sequence of an intended gene using the CRISPR technology, resulting in a gene matching that of a disease-resistant type, this process is known as “biomimicking.” Suppose the nucleotide variances between the gene of interest in domesticated and wild varieties do not significantly impact the plant’s ability to survive and produce. In that case, the researcher will only introduce the particular mutations linked to the disease resistance trait instead of swapping out the entire gene. Fusing a nuclease-dead Cas9 or nickase with cytidine deaminase enables high-precision point mutagenesis. This innovative method has demonstrated practical application in gene modification across various species (Komor et al., 2016).
Wang et al. (2017) demonstrated the use of the CRISPR/Cas9 technology for biomimicking via promoter replacement to enhance disease resistance in rice. The researchers achieved targeted gene expression alterations by substituting the native promoter of particular genes associated with disease susceptibility, increasing resistance to bacterial and viral pathogens. This example shows how promoter replacement using CRISPR may mimic natural plant defence systems and confer disease resistance in crop plants.
Zhang et al. (2017) used the CRISPR/Cas9 technology to create disease-resistant wheat plants for allele replacement. The researchers edited three homoeologs of the TaEDR1 gene, which is linked to powdery mildew susceptibility, to create precise mutations that mimic natural genetic variability and improve disease resistance. The resultant wheat plants demonstrated greater tolerance to powdery mildew infection.
By targeting a susceptibility gene involved in geminivirus infection, Ali et al. (2015) replaced the susceptible allele with a resistant allele using CRISPR/Cas9. The plants that were produced showed improved resistance to geminivirus infections.
. Need for synthetic R gene-mediated resistance
The largest group of R genes in plants consists of genes that encode nucleotide-binding site (NBS)–leucine-rich repeat (LRR) proteins. If altered, the LRR domain may be able to identify elicitors from a wide range of pathogens in addition to pathogen-derived elicitors that confer resistance. Resistance to certain strains of potato virus X (PVX) is granted by the potato R protein (Rx) LRR domain. Wheat R protein, Pm3F, with two amino acid substitutions, increases its potency against other pathogens (Stirnweis et al., 2014). For identifying isolate-specific Avr effectors (AVRa7, AVRa9, AVRa10, and AVRa22) of Blumeria graminis f. sp. hordei, the barley Mla locus contains many allelic variants (MLA7, MLA9, MLA10, and MLA22) (Saur et al., 2019). Therefore, recognition specificity can be changed via knowledge-based modification of the molecular structure of R genes. Using CRISPR/Cas tools, particularly prime and base editors, can be beneficial in modifying R genes for broad-spectrum resistance in vivo.
To create resistance against TuMV, Kim et al. (2016) modified the AvrPphB cleavage site in PBS1 to incorporate the cleavage site of the NIa protease from turnip mosaic virus (TuMV). This strategy has recently been expanded to soybeans to establish resistance against the soybean mosaic virus (SMV) (Pottinger et al., 2020). Utilising advanced genome editing techniques such as prime editing and HDR, we can precisely modify the endogenous PBS1 protein to confer resistance to diseases for which robust hereditary resistance is currently lacking. Integrated domains (IDs), present in specific NBS-LRR proteins (for example, Rice Pik-1), are believed to be crucial in detecting pathogen effectors. As a result, targeting IDs for receptor engineering holds significant promise in enhancing disease resistance. Kourelis et al. (2023) conducted a groundbreaking study that showed how resistance against a virus can be gained by replacing the integrated HMA domain of Pik-1 with an effector-binding VHH-nanobody. To create “Pikobodies,” which provide novel resistance to any pest or pathogen that delivers effectors inside plant cells, nanobodies capable of binding the desired effectors can be introduced into Pik-1 to replace the HMA domain.
It should be remembered that altering an already described R gene to produce novel resistance could cause it to lose its original function and become susceptible to a different infection. Saturated mutagenesis via prime editors (Xu et al., 2021) and dual base editors (Li et al., 2020b) will enable knowledge-guided directed evolution of R genes to broaden their resistance range. The HDR-mediated targeted insertion of multiple R genes into a specific genomic locus provides an attractive method for stacking R genes in a single background. Furthermore, it is essential to address the insert size restrictions and low efficiency of this approach (Dong et al., 2020). Novel R gene stacks created with HDR would be considered genetically modified organisms (GMOs) and might be subject to strict restrictions. Gain-of-function mutagenesis techniques such as CRISPR base and prime editors are employed to create disease resistance.
. Developing CRISPR-like artificial immunity
To defend against the pathogen of interest, host plants that express the Cas9-sgRNA complex targeted to virus genomes may build a bacterial immune system akin to CRISPR. Past research has employed this strategy to build immunity against viruses (Ali & Mahfouz, 2021; Mushtaq et al., 2020). Assume a host cell possesses a homologous sgRNA protospacer and an active Cas effector, both essential viral genome components. The viral life cycle can then be stopped by the Cas protein, causing damage to the viral genome. While Cas13 is ideal for targeting viral RNA genomes, Cas9 and Cas12 are suitable for targeting viral DNA genomes.
One drawback of this strategy is that a transgene-free immunity system is impossible because the Cas9 and sgRNA expression cassettes must always be present in the plant cell. Furthermore, viruses can develop mutations resistant to Cas cleavage, shielding them from CRISPR-like plant immunity.
. Achievements in CRISPR/Cas9-edited crops for pathogen resistance
Plant diseases caused by viruses, fungi, and bacteria result in significant losses, accounting for 20% to 40% of worldwide agricultural production (Boubakri, 2017). Enhancing plant resistance to infections is essential for addressing the needs of the growing global population. However, resistant strains of plant diseases can emerge through mutation or recombination, rendering disease management measures ineffective. Genome editing tools, such as the CRISPR/Cas system, have shown promise in addressing agricultural challenges, including developing novel disease-resistant crop plants. This is particularly important due to concerns about the overall use of pesticides and its potential impact on human health (Langner et al., 2018) (Figure 6).
. CRISPR/Cas9-based resistance to viruses
Plant viruses are obligate parasites and rely on host machinery for survival. Managing viral diseases is challenging because viruses evolve rapidly with insect vectors. The conventional strategy based on pathogen-derived resistance (PDR) is time-consuming. However, the CRISPR/Cas9 tool has shown significant promise in controlling viral infection and spread (Baltes et al., 2015; Ji et al., 2015; Liu & Zhang, 2020) (Table 2).
Table 2
CRISPR/Cas approaches increasing resistance by directly targeting viral genomes reviewed by Schenke and Cai (2020), extended in this work.
| Plant species | Target region | Resistance to virus/disease | Comment | Reference |
|---|---|---|---|---|
| Arabidopsis / N. benthamiana | IR, CP, Rep | Beet severe curly top virus (BSCTV) | Cas9 | Ji et al., 2015 |
| 1A, CP, 3’UTR-A | Cucumber mosaic virus (CMV) / Tobacco mosaic virus (TMV) | FnCas9 > RNA | Zhang et al., 2018a | |
| Arabidopsis | AtEIF(iso)4E | Turnip mosaic virus (TMV) | Cas9 | Pyott et al., 20162016 |
| Barley | MP/CP, Rep/RepA, LIR | Wheat dwarf virus (WDV) | Cas9 | Kis et al., 2019 |
| Cassava | MenCBP-1/2 | Cassava brown streak virus (CBSV) | Cas9 | Gomez et al., 2019 |
| Cotton | Six overlapping genes of the three most prevalent strains of CLCuV | CLCuV | Engineering broad-spectrum resistance by co-infiltration with multiplex gRNA cassette | Mubarik et al., 2021 |
| Cucumber | CseIF4E | Several viral diseases: CVYV, ZYMV, PRSV-W | Cas9 | Chandrasekaran et al., 2016 |
| N. benthamiana | RBS, IR hairpin, 3× Rep | Bean yellow dwarf virus (BeYDV) | Cas9 | Baltes et al., 2015 |
| IR, CP, RCRII of Rep | Tomato yellow leaf curl virus (TYLCV) | Cas9 | Ali et al., 2015 | |
| IR, CP, RCRII of Rep | Geminiviruses (CLCuKoV, MeMV) | Cas9 | Ali et al., 2016 | |
| CP, Hc-Pro | Turnip mosaic virus (TuMV) | Cas13a > RNA | Aman et al., 2018a | |
| IR, C1 | Cotton leaf curl Multan virus (CLCuMuV) | Cas9 | Yin et al., 2019 | |
| Potato | P3, CI, Nib, CP | Potato virus Y (PVY) | Cas13a > RNA | Zhan et al., 2019 |
| PI, HC-Pro, P3, Cl1, Cl2, and VPg genes | Broad-spectrum resistance against multiple PVY-strains | CRISPR/Cas13 | Noureen et al., 2022 | |
| Rice | OseIF4G | Rice tungro spherical virus (RTSV) | Cas9 | Macovei et al., 2018 |
| Soybean | GmF3H1/2, FNSII-1 | Soybean mosaic virus (SMV) | Cas9 | Zhang et al., 2020b |
| SMV HC-Pro gene | Soybean mosaic virus (SMV) | CRISPR/CasRx | Gao et al., 2024 | |
| Tomato | CP, Rep | Tomato yellow leaf curl virus (TYLCV) | Cas9 | Tashkandi et al., 2018 |
| SlPelo | Tomato yellow leaf curl virus (TYLCV) | SpCas9 | Pramanik et al., 2021 | |
| SleIF4E1 | Resistance to potato virus Y (PVY) | CRISPR/Cas9-sgRNA | Kumar et al., 2022 | |
| SleIF4E2 |
. Resistance against DNA virus
Plant DNA viruses, geminiviruses, and pararetroviruses infect various plants, causing food instability in many nations. Geminiviridae is made up of single-stranded DNA (ssDNA) and can affect a variety of commercially significant crops globally, including those from the families Solanaceae, Fabaceae, Cucurbitaceae, Euphorbiaceae, and Malvaceae. Cas9 and guide RNAs that target a viral genome can be expressed in the host plant to develop artificial immunity to viruses. The bean yellow dwarf virus (BeYDV) was effectively controlled using CRISPR/Cas9-mediated viral genome collapse (Baltes et al., 2015). Tomato yellow leaf curl virus (TYLCV) accumulated less in transgenic tomatoes and Nicotiana benthamiana when Cas9 was used to target the viral coat protein or replication machinery (Tashkandi et al., 2018). Ji et al. (2015) reported that transgenic Arabidopsis and N. benthamiana, which overexpressed Cas9-sgRNA, demonstrated a strong resistance to infection by the beet-severe curly top virus (BSCTV). By focusing on overlapping genes or changing viral movement protein or coat protein (CP) genes, CRISPR-multiplexing has been thriving in producing resistance against the cotton leaf curl virus (Mubarik et al., 2021), wheat dwarf virus (Kis et al., 2019), and banana streak virus (Tripathi et al., 2019). Similarly, Cas9 and many sgRNAs have been used to knock out the CP region of the cauliflower mosaic virus in transgenic Arabidopsis to establish resistance (Liu et al., 2018). The rice black-streaked dwarf virus (RBSDV) causes maize rough dwarf disease (MRDD), which causes dwarfism in maize plants. It has recently been shown that ZmGDIα targeted mutation using CRISPR enhances resistance to MRDD without negatively impacting agronomic performance (Liu et al., 2022).
According to previous research, it has been demonstrated that focusing on the origin of replication located in the intergenic region of the DNA is more potent in achieving desired outcomes than directing interventions towards coding sequences, such as the coat protein and replicases (Ali et al., 2015). Geminivirus genomes have rapidly evolved to produce viral variants that can evade CRISPR/Cas9 protection in Cassava and N. benthamiana (Ali et al., 2016; Mehta et al., 2019). The intergenic region is less prone to mutation, allowing prolonged resistance. Viruses utilise concealed open reading frames, and characterising them could aid in developing effective antiviral strategies using CRISPR/Cas systems (Chiu et al., 2022; Molla et al., 2020a). The recent discovery of Cas14, which cleaves ssDNA, opens the door to targeting geminiviral ssDNA (Harrington et al., 2018). Cas14 could be employed in conjunction with Cas9 to target both entering ssDNA and replicative dsDNA, significantly strengthening resistance (Khan et al., 2019). In addition, various plant species have developed resistance to begomoviruses using the CRISPR/Cas9 system. This system targets specific viral regions cleaved during replication (Ali et al., 2015). Additionally, crop plants such as cucumber have gained resistance against zucchini yellow mosaic virus (ZYMV), papaya ringspot mosaic virus (PRSMV), and cucumber vein yellowing virus (CVYV), which belong to the potyvirus and ipomovirus families, respectively.
In 2019, Gomez et al. (2019) used the CRISPR/Cas9 system to target the eIF4E isoforms, which are novel cap-binding protein-1 and 2, linked to viral genome proteins. They discovered that a mutation in these isoforms decreased the signs of the cassava brown streak virus (CBSV). Additionally, a mutation in the translation initiation factor 4 gamma gene (eIF4G) in rice enhanced the crop’s resistance to both the rice tunic bacilliform virus (RTBV) and the rice tunic spherical virus (RTSV). According to Macovei et al. (2018), these two viruses cause rice tungro disease (RTD) and result in substantial economic losses. More recently, A. thaliana resisted the clover yellow vein virus (ClYVV) after targeting the eIF4E1 gene using CRISPR/Cas9 (Bastet et al., 2018). One essential protein needed for the infection cycle of the single-stranded positive-sense RNA viruses belonging to the Potyviridae family is called Eukaryotic Translation Initiation Factor 4E (eIF4E). It has been demonstrated that protection against potyviruses is conferred in many plant species by interfering with the interaction between eIF4E and the viral protein genome-linked (VPg) element at the 50-terminal of potyviruses (Bastet et al., 2017). In separate investigations, researchers have effectively disrupted eIF4E genes in Arabidopsis and Cucumis sativus (cucumber) to produce resistance against potyviruses and ipomoviruses using CRISPR/Cas9 (Chandrasekaran et al., 2016; Pyott et al., 2016).
. Resistance against RNA virus
Recently, new CRISPR-associated proteins targeting and editing RNA have been identified. Among these proteins, we can find Cas9 from Francisella novicida (referred to as FnCas9) and Cas13a, previously known as C2c2, which originates from Leptotrichia shahii (known as LshCas13a) (Shmakov et al., 2015). LshCas13a, a groundbreaking Cas13 ortholog, represents a significant milestone as it enables programmable RNA-targeting activities. This development expands the applications of CRISPR/Cas systems from solely targeting DNA to also targeting RNA (Abudayyeh et al., 2016).
A genetically modified variant of the FnCas9 enzyme, optimised for plant codons, along with RNA-guided RNA targeting, was employed to impart resistance against cucumber mosaic virus (CMV) and tobacco mosaic virus (TMV) in transgenic N. benthamiana and A. thaliana plants, respectively (Zhang et al., 2018b). A newly discovered RNA-targeting CRISPR/Cas system, LshCas13a, has been found to utilise the class 2 type VI-A CRISPR/Cas effector. This system has been specifically engineered to target and cleave single-stranded RNA (ssRNA) viruses present in plants. When paired with a sgRNA that targets viral RNA sequences, LshCas13a demonstrated swift and potent RNA interference against the turnip mosaic virus (TuMV). Notably, the sgRNA was under the control of a pea early browning virus (PEBV) promoter within a tobacco rattle virus (TRV)-based vector, and this system was tested in LshCas13a transgenic N. benthamiana or A. thaliana plants (Aman et al., 2018a; Aman et al., 2018b). Similarly, utilising this method, it was confirmed that transgenic tobacco and rice carrying LshCas13a selectively targeting viral RNA were resistant to TMV, rice stripe mosaic virus (RSMV), and southern rice black-streaked dwarf virus (SRBSDV) (Zhang et al., 2019). This CRISPR/Cas13a has also shown its efficacy in disrupting and impeding PVY infection by targeting the P3-, NIb-, or CP-coding regions in the potato virus Y (PVY) genomic area (Zhang et al., 2019). Moreover, a novel Cas13 protein from Ruminococcus flavefaciens was identified and categorised as a type VI-D effector named Cas13d (CasRx). When Cas13d was utilised to target the GFP, CP, or HC-Pro area in the TuMV virus, researchers discovered that it worked more effectively than Cas13a, Cas13b, or other Cas13 variants.
. CRISPR/Cas9-based resistance to fungi
The “mildew resistance locus” (Mlo) is a well-known S-gene mutation that provides resistance to powdery mildew in barley. By utilising loss-of-function Mlo alleles, barley variants with powdery mildew resistance have been developed. These resistant plants have demonstrated long-term resilience to highly pathogenic strains of powdery mildew in field cultivation, showcasing the effectiveness of S-gene-based resistance (Jørgensen, 1992). The discovery of the barley Mlo gene through cloning showed that it is present in various plant species and is found as a multi-copy gene family in higher plants. This suggests that the Mlo gene is widely conserved across the plant kingdom and plays a significant role in the genetic makeup of higher plants (Büschges et al., 1997; Devoto et al., 2003). Furthermore, many plant species, including Arabidopsis, tomato, pea, pepper, tobacco, bread wheat, and possibly grapevine and peach, have been linked to powdery mildew susceptibility due to Mlo like genes (Appiano et al., 2015; Bai et al., 2008; Consonni et al., 2006; Feechan et al., 2008; Humphry et al., 2011; Wang et al., 2014; Zheng et al., 2013). Grapevine powdery mildew resistance was increased by the CRISPR/Cas9 deletion of VvMLO7 and VvMLO3 (Malnoy et al., 2016; Wan et al., 2020). The RNP-based technique has been effectively applied to alter three (DIPM-1, DIPM-2, and DIPM-4) genes in apple (Malus domestica) protoplasts to produce resistance against Erwinia amylovora, the fire blight (Malnoy et al., 2016). Four guide RNAs were employed in the CRISPR/Cas9 strategy to target the VvWRKY52 transcription factor. In contrast to monoallelic mutant plants, the results revealed 21% of B. cinerea (Wang et al., 2018). The CRISPR/Cas9 system temporarily targeted the immune system suppressor gene, Pathogenesis-Related 3 (NPR3), in cocoa leaves, increasing resistance to Phytophthora tropicalis. The application of the CRISPR/Cas9 technology has led to the identification of a novel selection marker for Phytophthora species, a group of oomycetes known for causing devastating plant diseases. This significant breakthrough emerged from genetically modifying a fungicide-resistant gene known as PcMuORP1 (Wang et al., 2019). Resistance against rice blast disease is conferred by OsSEC3A and OsERF922 gene disruption mediated by CRISPR/Cas9 (Ma et al., 2018; Wang et al., 2016). Furthermore, rice with the pi21 gene exhibited long-lasting resistance against rice blast (Zhou et al., 2022). Additionally, the production of OsSEC3A mutants improved rice resistance to disease caused by Magnaporthe oryzae and resulted in a pleiotropic phenotype with a rise in salicylic acid (SA) concentration. Additionally, many genes linked to pathogenesis and SA were upregulated (Ma et al., 2018). By leveraging the CRISPR/Cas9 technology, researchers have successfully enhanced rice’s resistance to the devastating rice blast disease caused by the fungus M. oryzae. This advancement was achieved by targeting the negative regulators associated with the plant’s defence mechanism. Moreover, a groundbreaking CRISPR/Cas9 chromosomal rearrangement technique for soybeans has also been revealed (Nagy et al., 2021). Through the artificial creation of chimeric paralogs, this work generated two NLR gene clusters, named Rpp1-like and Rps1, which are associated with resistance to Phakopsora pachyrhizi and P. sojae, respectively. It also demonstrated a potential cutting-edge method for managing plant disease. In summary, the advantages, effectiveness, and potential of the CRISPR/Cas-based editing technology to improve crop plant resistance were ascertained by all of these successful fungal disease resistance outcomes (Table 3).
According to Frye et al. (2001), edr1 mutant plants in Arabidopsis demonstrated tolerance against bacteria and oomycetes. On the other hand, Taedr1 mutant plants in wheat produced immunity to Bgt after all homologs of EDR1 were simultaneously altered using CRISPR/Cas9 (Zhang et al., 2017). According to Nekrasov et al. (2017), tomato resistance to Oidium neolycopersici was enhanced by SlMlo1 deletion using CRISPR/Cas9. Nevertheless, powdery mildew resistance 4 (PMR4), a gene encoding callose synthase, has also been altered by GET, which has resulted in the establishment of resistance to O. neolycopersici in tomato and Arabidopsis as well. Similar to this, rice plants’ resistance to M. oryzae has increased due to a mutation in the Pi21 gene’s coding area generated by CRISPR/Cas9 (Li et al., 2019b). In addition, Xie and Yang (2013) reported that CRISPR/Cas9 was also applied to rice, specifically targeting the coding area of Oryza sativa mitogen-activated protein kinase 5 (OsMAPK5) to enhance resistance to Magnaporthe grisea. In a study by Wang et al. (2018), it was discovered that grapevines with homozygous mutants of VvWRKY52 showed improved resistance to Botrytis cinerea. The researchers developed four gRNAs targeting the VvWRKY52 transcription factor using the CRISPR/Cas9 system. Additionally, the application of CRISPR/Cas9 has led to heightened resistance against Phytophthora tropicalis and Phytophthora palmivora in papaya (Gumtow et al., 2018) and cacao (Fister et al., 2018) plants, respectively. These instances of CRISPR/Cas9 applications highlight its vital role in genetic engineering technologies, especially in enhancing plant resistance to fungal pathogens.
Table 3
Applications of CRISPR/Cas9 for plant resistance to fungal and oomycete pathogens reviewed by Paul et al. (2021) extended in this work.
| Fungal or oomycete pathogen | Plant species | Target gene | Encoding protein/function of gene | Disease resistance/phenotype | Method | Reference |
|---|---|---|---|---|---|---|
| Venturia inaequalis (apple scab) | Apple | MdERF3 | Trihydroxynaphthalene reductase (THN) gene | Reduced melanisation | Cas9 | (Rocafort et al., 2022) |
| Venturia inaequalis (apple scab) | Apple | MdERF3 | Trihydroxynaphthalene reductase (THN) gene | Reduced melanisation | Cas9 | (Rocafort et al., 2022) |
| Blumeria graminis f. sp. hordei (apple scab) | Barley | mlo | Intracellular calcium signaling | Enhanced resistance | CRISPR/Cas9 | (Koide et al., 2023) |
| Leptosphaeria maculans (apple scab) | Apple | MdERF3 | Trihydroxynaphthalene reductase (THN) gene | Reduced melanisation | Cas9 | (Rocafort et al., 2022) |
| Leptosphaeria maculans | Brassica napus (rapeseed) | AvrLm7 | Effector protein/Recognized by host R gene | Enhanced virulence | CRISPR/Cas9 | (Zou et al., 2020) |
| Phytophthora palmivora | Carica papaya (Papaya) | PpalEPICs | Cysteine protease inhibitor/Inhibition of Cysteine protease in host | Induced disease resistance in papaya | CRISPR/Cas9 and Agrobacterium-mediated transformation | (Gumtow et al., 2018) |
| Phytophthora sojae | Glycine max (Soybean) | PsSu(z)12 | Core subunit of H3K27me3 methyltransferase/Participate in gene silencing | Induces immune recognition by soybeans | CRISPR/Cas9 | (Wang et al., 2020) |
| PsAvr3c | RxLR effector/Promotes host susceptibility | Decreased aggressiveness | CRISPR/Cas9 | (Huang et al., 2017) | ||
| Avr4/6 | RxLR effector/Promotes host susceptibility | Evasion of R resistance in Rps soybean line | CRISPR/Cas9 | (Fang & Tyler, 2016) | ||
| Peronophythora litchii | Litchi chinensis (Litchi) | PlPAE5 | Pectin acetylesterase/Consume host pectin as a carbon source | Less invasion | CRISPR/Cas9 | (Kong et al., 2019) |
| Magnaporthe oryzae | Rice | OsERF922 | Negative regulator of blast resistance | Enhanced blast resistance | CRISPR/Cas9 | (Wang et al., 2016) |
| OsSEC3A | Subunit of the exocyst complex 3a | Enhanced resistance | CRISPR/Cas9 | (Rocafort et al., 2018) | ||
| Pi21 and | Broad-spectrum resistance that encodes a protein that includes a putative heavy metal–binding domain and protein-protein interaction domain | Enhanced resistance | CRISPR/Cas9 | (Nawaz et al., 2020) | ||
| Bsr-d1 | C2H2‐type transcription factor | Enhanced resistance | CRISPR/Cas9 | (Zhou et al., 2022) | ||
| Claviceps purpurea | Secale cereale (Rye) | CpTrpE, Cppyr4 | α-subunit of anthranilate synthase/Key enzyme of tryptophan biosynthesis | Decreased virulence | CRISPR/Cas9 and Homologous recombination (HR) knock-out | (Králová et al., 2021) |
| Oidium neolycopersici (apple scab) | Tomato | Mildew resistant locus o (SlMLO) | Membrane-associated protein | Enhanced resistance | CRISPR/Cas9 | (Nekrasov et al., 2017) |
| Powdery mildew resistance 4(SlPMR4) | Callose synthase | Enhanced resistance | CRISPR/Cas9 | (Santillán Martínez et al., 2020) | ||
| Phytophthora infestans | miR482b and miR482c | Negative regulator by regulating NBS–LRRs | Enhanced resistance | CRISPR/Cas9 | (Hong et al., 2021) | |
| Botrytis cinerea, Fusarium acuminatum, Rhizopus stolonifer | Pectate lyase (SlPL) | Pectin-degrading enzyme | Enhanced resistance | CRISPR/Cas9 | (Silva et al., 2021) |
. CRISPR/Cas9-based resistance to bacteria
Significant risks to agriculture are posed by phytopathogenic bacteria, which can produce rots, mosaics, spots, and other difficult-to-control illnesses that severely reduce productivity. Bacteria become resistant to antibacterial agents and proliferate infections due to their constantly changing environment and horizontal gene transfer. Bacteria typically gain entry into cells through mechanical wounds, stomata, stigma, and other plant openings. Additionally, they can penetrate cells by releasing bioactive chemicals that facilitate their entry (Zeng et al., 2010). Upon identifying these signals, the plant defence mechanism takes the necessary action to eradicate the infection (Figure 1). Nonetheless, numerous host plant genes, including a few “S” genes, aid in the complicated signal cascade that successful pathogens employ to overcome plant protection and start an infection (Das et al., 2018; Dong & Ronald, 2019). CRISPR/Cas9 techniques were utilised to modify tomato plants that were under attack by bacterial plant infections, such as Pseudomonas syringae and Xanthomonas spp. This led to a decrease in tomato yield and significant economic damage (Schwartz et al., 2015). According to Zeilmaker et al. (2015) and Langner et al. (2018), P. syringae pv. tomato infection was found to increase the expression of DMR6 (downy mildew resistance 6), function as a negative regulator of plant immunity, and aid in the spread of infection. By utilising CRISPR/Cas9, scientists were able to remove the tomato counterpart SlDMR6-1, resulting in an improved resistance of tomato plants to multiple diseases, such as P. syringae pv. tomato and Phytophthora capsici. Remarkably, this enhancement in disease resistance was achieved without causing any damage to the overall health of the plants (Thomazella et al., 2021). In another case, OsSWEET13, a sensitive host gene implicated in sucrose transfer during disease, was edited using CRISPR/Cas9 to confer resistance against the bacterial blight pathogen Xanthomonas oryzae pv. oryzae (Zhou et al., 2014). When Zhou et al. (2015) deleted the OsSWEET13 gene from rice, a null mutant with increased resistance to X. oryzae was produced. Similar to this, DspE is the name of the pathogen effector protein produced by Erwinia amylovora. The dspE protein interacts with DspE-interacting proteins of Malus (DIPM 1, 2, 3, 4) genes (Gimenez-Ibanez et al., 2017; Das et al., 2018). This interaction has been found to enhance plant susceptibility to fire blight disease. To build resistance against this disease, researchers utilised CRISPR/Cas9 to truncate these DIPM genes in apple protoplasts (Malnoy et al., 2016). A viable strategy for sustainable agriculture could be transgenic crops with unique alleles and broad disease resistance (Table 4).
Table 4
CRISPR/Cas9 application for plant bacterial disease resistance reviewed by Schenke and Cai (2020) extended in this work.
| Plant species | Target region | Resistance to pathogen/pest | Comment | Reference |
|---|---|---|---|---|
| Apple | MdDIPM4 | Erwinia amylovora | CRISPR/Cas9-FLP/FRT-based gene editing system | Pompili et al., 2020 |
| Arabidopsis | AtIAN9 | Pseudomonas syringae (bacterial speck) | Cas9 | Wang et al., 2018 |
| Duncan grapefruit | CsLob1 | Xanthomonas citri | Cas9 promoter | Jia et al., 2016 |
| Maize | ZmNLB18 (WAK) | Setosphaeria turcica (Northern Leaf Blight) | Cas9 HDR | USDA, 2017 |
| Rice | OsSWEET11/14 | Xanthomonas oryzae (bacterial blight) | Cas9 promoter | Jiang et al., 2013 |
| OsSWEET13 | Xanthomonas oryzae | Cas9 | Zhou et al., 2015 | |
| OsBsr-d1 | Magnaporthe oryzae | Cas9 | Li et al., 2017 | |
| OsSec3A | Magnaporthe oryzae | Cas9 | Ma et al., 2018 | |
| OsXa13 | Xanthomonas oryzae | Cas9 promoter | Li et al., 2020a | |
| OsPi21, OsXa13 | Magnaporthe oryzae / Xanthomonas oryzae | Cas9 multiplex | Li et al., 2019b | |
| Os8N3 | Xanthomonas oryzae | Cas9 | Kim et al., 2019 | |
| OsCul3a | Xanthomonas oryzae / Magnaporthe oryzae | Cas9 | Gao et al., 2020, Liu et al., 2017 | |
| OsSULRT3; 6 | Xanthomonas oryzae pv. oryzicola (Xoc) | CRISPR/Cas9 | Xu et al., 2021 | |
| Xa13 | Xanthomonas oryzae pv. oryzae (Xoo) | CRISPR/Cas9 | Li et al., 2022 | |
| Tomato | SlDMR6 | P. syringae, P. capsici, Xanthomonas spp. | Cas9 | Thomazella et al., 2021 |
| SlJAZ2 | Pseudomonas syringae pv. tomato DC3000 | Cas9 | Ortigosa et al., 2019 |
. Limitations of the CRISPR/Cas9 Technology
With the advent of CRISPR/Cas9, numerous avenues for precisely and accurately altering genomes to create superior crop types have become accessible. A new era of breeding systems is ushered in by CRISPR tools, which enhance plant defence by disrupting the beneficial relationship between pathogens and hosts. The rapid acquisition of resistance traits is attributed to the direct introduction of specific mutations into susceptibility alleles, leading to the development of resistance characteristics. While the CRISPR/Cas technologies provide unparalleled prospects for resistance breeding, there are significant constraints that could impede their widespread application. One of the main obstacles is that many plant species lack appropriate methods for plant tissue culture and regeneration. Certain tissue culture media are designed for specific genotypes and may not be suitable for other genotypes of the same species due to genetic differences. Growth regulators expressed transiently may enhance the frequency of genome editing and regeneration (Debernardi et al., 2020). The success of editing experiments is hampered by the lack of understanding regarding species-specific promoters that drive Cas protein expression and guide RNAs. We could get beyond this obstacle with the development of basic molecular biology toward the identification of natural promoters and the creation of synthetic promoters (Jores et al., 2021). We lack a thorough grasp of the interactions between the pathogen and host in many diseases. More targets for genome editing to increase disease resistance will become clear through research on understanding the molecular details of disease response. The accuracy and frequency of mutant production are major issues since agricultural plants have a longer life cycle than model plants. Consequently, more investigation is required into the quick and easy methods for verifying the resistant phenotype through the transient expression of CRISPR/Cas9 components.
The CRISPR/Cas9 technique has several limitations, including the larger protein size, restricted PAM site, increased off-target mutations, low HDR efficiency, low effectiveness against viruses, expensive and time-consuming Agrobacterium-mediated transformation system, and challenges in commercialising newly edited crops due to ethical concerns, national laws, and regulations.
. Conclusion and future perspectives
The impact of climate change is anticipated to intensify the challenge of protecting crops as it paves the way for the emergence of new infectious agents and the escalation of disease outbreaks (Chaloner et al., 2021). While CRISPR/Cas9 shows promise for improving plant resistance to diseases, the potential side effects of editing or modifying plant susceptibility genes pose a major obstacle. These susceptibility genes are linked to both plant development and fitness, and their alteration can lead to a range of effects across multiple traits (pleiotropic effects) (Tyagi et al., 2021). This is also the most commonly reported effect after artificially enhancing plant immunity by knocking off a negative regulator (Ding et al., 2018). The CRISPR/Cas9 system also introduces off-target alterations, which is an essential problem to consider. An off-target mutation refers to an undesired result that causes imprecise alterations in genomic sequences, potentially impacting the structure and function of proteins. To address this concern, scientists are actively employing advanced computational techniques, such as Guide-seq, Diagenome-seq, and DISCOVER, while also refining CRISPR components like Cas proteins and gRNA to mitigate the risk of off-target effects (Ahmad et al., 2020). In addition, to reduce off-target mutation and boost the effectiveness of the CRISPR system, researchers are currently employing machine learning methods like Bowtie or Elevation, linear regression model, and Random Forrest Model. While these models have shown promise in several scenarios, it has been shown that their effectiveness is limited to the particular goal for which they were designed. Therefore, for the CRISPR/Cas9 system to be achieved, an improved model that can operate effectively with a border target range must be developed (Hahn & Nekrasov, 2019). Recently, EvolvR, a modified CRISPR/Cas9 tool, has been developed to enable the alteration or introduction of nucleotide sequences at specific regions of the genome using gRNAs. EvolvR incorporates an error-prone DNA polymerase system, allowing more precise and controlled genetic modifications (Halperin et al., 2018; Sadanand, 2018). In contrast, scientists redesigned the Cas9 proteins to reduce the number of off-target mutations. Various Cas9 variations, including eSpCas9, HF-Cas9, HyperCas, and SniperCas9, were developed to enhance the effectiveness and precision of these proteins. Additional Cas proteins, such as NmCas9 (Neisseria meningitidis) (Hou et al., 2013), SaCas9 (Staphylococcus aureus) (Ran et al., 2015), StCas9 (Streptococcus thermophilus) (Müller et al., 2016), FnCas9 (Francisella novicida) (Hirano et al., 2016), and CjCas9 (Campylobacter jejuni) (Kim et al., 2017), have been discovered and are characterised by their smaller size compared to SpCas9. According to Wada et al. (2020), these proteins have advantages in terms of gene transport and more precise gene targeting. RNPs are tiny ribonuclear proteins that can break down after being expressed in the intended system and have shorter lifespans. Consequently, this lowers the system’s probability of off-target mutation (Liang et al., 2018; Liang et al., 2017). The CRISPR/Cas9/RNP complex has been effectively utilised in various studies, as indicated by multiple investigations. Jaiswal et al. (2024) discovered that Anti-CRISPR (Acr) proteins in phages are highly significant. Acrs are small proteins found in phages, and they serve as protective agents against the CRISPR/Cas bacterial immune system. These proteins can be utilised to create controlled and reversible CRISPR/Cas-mediated systems, which in turn facilitates more accurate genome editing. Molla et al. (2024) further describe how Acrs can reduce the off-target consequences of the CRISPR/Cas technology, increasing its potential as a genome editing tool.
Genome editing with CRISPR/Cas9 might rapidly generate the desired disease resistance compared to conventional or molecular breeding methods. However, because fungal and oomycete populations evolve quickly and have a diverse genetic makeup, infections may eventually overcome single gene-editing-based disease resistance. Targeting numerous genes and/or altering the genomes of pathogens and plants utilising these quickly evolving CRISPR/Cas9 technologies could be effective strategies for long-term disease management. Another intriguing strategy is to use the CRISPR/Cas9 system to beneficial fungal species, such as Trichoderma sp., to boost plant immunity and act as a biocontrol agent against oomycete and fungal infections. CRISPR/Cas9-based genome editing of plants and pathogens is expected to be widely used to increase plant disease resistance. In the future, disease-resistant transgene-free crops will be required to meet the world’s food needs.
By accurately inserting genomic changes, advanced CRISPR techniques like base editor and prime editor hold enormous promise for creating disease-resistant types. Variations in the DNA methylation state caused by epigenome editors are inherited; persistent epialleles can break compatible host-pathogen interactions to confer resistance to disease. To integrate disease-resistant features with other agronomically important traits and achieve broad-spectrum durable resistance, multiplex editing creates new opportunities to target numerous genetic loci at once simultaneously. Moreover, the utilisation of CRISPR-based instruments in conjunction with genome-wide guide RNA libraries and consequent phenotypic screening has the potential to propel noteworthy progress in functional genomics and unveil an array of innovative strategies for combating infections and pests.
To acquire a thorough overview of biological activities in cellular processes, researchers have more recently coupled metabolomics with other “omics” methodologies (Bao et al., 2020). For instance, metabolomics analysis of Ciboria shiraiana and Phtyophthora sojae infections were carried out in response to mulberry fruit and soybean hypocotyls, respectively (Castro-Moretti et al., 2020; Zhu et al., 2018). This research paves the way for future investigations of crucial metabolic determinants in plant–pathogen studies. Future developments in the field of genetic engineering and genomics (GE/omics) are anticipated to enable the development of CRISPR systems in conjunction with other methodologies to produce plant types resistant to disease and biotic stressors, while also meeting society’s food needs. Despite the potential benefits, the use of continuously expressing nuclease crops is considered to involve genetically modified organisms (GMOs), which are subject to regulatory controls and banned in several countries due to concerns about their environmental and health impacts. As a result, there is a growing preference for developing transgene-free processes that can simultaneously utilise multiple pathogen effectors and resistant genes, allowing the production of non-GMO crops. It is important to recognise that policies regarding GMOs are influenced by a variety of factors, including cultural differences, environmental conditions, political pressures, and the interests of stakeholders, such as farmers, agro-industrialists, environmental activists, and government agencies. Across different nations, governments respond differently to public opinion and the views of the scientific community, leading to a wide range of regulatory approaches to GMOs. Recently, the USA and Canada have permitted the cultivation and sale of CRISPR without a genetically modified organism label, exempting it from the definition of a GMO under regulatory frameworks. According to USDA regulations, crops modified with CRISPR/Cas9 may be grown and sold without being subject to oversight by regulatory agencies (Waltz, 2018). The government of India has also permitted the SDN-1 category, creating hope for gene-edited crop approval and paving the way for its commercialisation. The CRISPR/Cas9 technology is a powerful and versatile tool for gene editing and regulation. It is driving advancements in various fields of study and is becoming the cornerstone of sustainable agriculture. In the future, its diverse applications in plant and microbial biology research are expected to significantly enhance our knowledge of fundamental biological processes and disease resistance mechanisms.
In conclusion, the CRISPR/Cas9 technology has much potential for use in agriculture. It offers opportunities to increase crop resistance, raise yield and quality, and solve issues related to global food security. Using the precision and efficiency of this technology, scientists can develop crops that are more resilient to biotic and abiotic stresses while enhancing their nutritional value and yield. However, to fully harness the potential of the CRISPR/Cas9 technology in agricultural innovation, further extensive research, development, and strategic implementation are essential.
