. Introduction
Mount Gede Pangrango National Park – ‘Taman Nasional Gunung Gede Pangrango’ (TNGGP), located in West Java, Indonesia, includes the volcanic landscapes of Mount Gede (2,958 m asl) and Mount Pangrango (3,019 m asl). UNESCO designated TNGGP as a biosphere reserve in 1977, making it an important site for biodiversity protection, education, and scientific research (Yani et al., 2022). The park, which spans the administrative regions of Bogor, Cianjur, and Sukabumi, is home to a diverse range of flora and fauna, including approximately 150–200 orchid species that thrive in the forest understory (Prapitasari et al., 2020). However, the orchid population in this region has declined significantly due to habitat alteration and environmental stress, resulting in numerous species being listed on the IUCN Red List and CITES protection lists (CITES, 2021).
Terrestrial or ground orchids are known to host a wide variety of endophytic fungi in their root systems. These endophytes frequently engage in mutualistic interactions with their host plants and may greatly improve host fitness by encouraging growth, increasing stress tolerance, and providing pathogen protection (Bayman & Otero, 2006; Chitnis et al., 2020). Endophytic fungi’s ability to generate bioactive secondary metabolites comparable to those found in the host plant facilitates such interactions (Mishra & Sharma, 2022).
In the context of red chili (Capsicum annuum), a high-value crop in Indonesia, endophytic fungi play a critical role in combating major biotic stressors, particularly Phytophthora capsici, a destructive oomycete pathogen responsible for Phytophthora blight. This disease is a major hazard to the chili plant’s lifecycle, infecting roots, stems, and fruits (Wan & Liew, 2020). Early identification challenges and rapid pathogen multiplication contribute to crop losses that surpass 40% in severe cases (Wang et al., 2024).
Endophytic fungi have gained attention for their antagonistic activity against P. capsici. These fungi live within plant tissues without causing harm and can support host resistance by producing bioactive compounds such as alkaloids polyketides belonging to several classes (Mondol et al., 2017). Their compatibility with host plants and persistence in the plant environment make them ideal biocontrol candidates. Recent studies highlight the promise of several endophytes in inhibiting P. capsici. For instance, Penicillium strains MAG1 and PAB2 demonstrated antibiosis through clear inhibition zones of 4.5–4.8 mm in an in vitro assay (Ramdan et al., 2018). While endophytic fungi possess antagonistic potential, their standalone efficacy may be limited; a more promising strategy, however, involves using biological substances in synergistic combinations. Bae et al., (2011) reported that endophytic fungi Trichoderma spp. greatly reduced plant death and increased vegetative growth in pepper plants infected with P. capsici. These dual uses are most likely achieved through a variety of processes, including direct mycoparasitism, competition for nutrients and space, and the creation of antifungal metabolites. Mmbaga and Gurung (2018) conducted another crucial study in which they investigated Nigrospora sphaerica strain A22F1 and its efficiency as a biocontrol agent. Although not the most powerful in dual culture assays, A22F1 dramatically reduced disease severity in greenhouse trials when administered before pathogen exposure. Shah et al. (2019) conducted an in-depth investigation into the diversity, identification, and functional characterization of endophytic fungi associated with the roots of the medicinal and ornamental orchid Dendrobium moniliforme. They evaluate the plant growth-promoting (PGP) attributes of these endophytes and assess their potential for use in orchid propagation and sustainable plant management.
Despite their ecological significance and biotechnological promise, endophytic fungi from terrestrial orchids, particularly those found in Indonesian conservation forests such as TNGGP, are understudied. Previous research has shown that certain endophytes, such as Fusarium spp., Penicillium spp., and Trichoderma asperellum, can act as biocontrol agents against soil-borne pathogens like P. capsici, a notorious pathogen that causes Phytophthora blight in chili plants (Capsicum annuum) (Ramdan et al., 2018; Irawati et al., 2020). Moreover, some endophytic fungi have demonstrated plant growth-promoting capabilities, particularly those colonizing root tissues like Dark Septate Endophytes (DSEs), which enhance nutrient uptake and increase biomass in orchid species and other plants (Hou & Guo, 2009).
Despite their established ecological and biotechnological roles, endophytic fungi from terrestrial orchids in conserved environments such as TNGGP are generally unknown and underutilized. There is a particular lack of integrative research on their dual roles in plant growth promotion and biocontrol efficacy, adaptability across plant species, and the functional processes that underpin these interactions. Furthermore, there is a lack of sufficient baseline data from Indonesian forests to support conservation-oriented bioprospecting and fungal biodiversity recording.
Given TNGGP’s ecological richness and the known relationships between terrestrial orchids and endophytic fungi, the objectives of this study were to investigate and characterize fungal endophytes found in terrestrial orchid roots. Specifically, the objectives included a) isolation and identification of endophytic fungi from terrestrial orchid roots in TNGGP and b) assessing the ability of these isolates to stimulate plant development with Chinese cabbage (Brassica rapa var. pekinensis) as an indicator species. Furthermore, the biocontrol efficiency of selected fungal isolates against Phytophthora capsici in chili (Capsicum annuum) were evaluated in vivo.
This research answers two key questions: whether endophytic fungi from terrestrial orchid roots in TNGGP can promote plant growth while suppressing P. capsici in chili plants, as well as whether endophytic fungi have the greatest potential for acting as both growth stimulants and biocontrol agents. We hypothesize that some endophytic fungi associated with terrestrial orchids in TNGGP have bioactive properties that can promote plant growth while suppressing P. capsici infection. The findings of this study are expected to contribute valuable baseline data for the development of endophytic fungi-based biofertilizers and biocontrol agents that can be sustainably applied to horticultural crops, particularly in managing diseases caused by P. capsici.
. Materials and methods
. 2.1. Experimental procedures
This study was conducted at the Indonesian Culture Collection (InaCC) and the Genomics Laboratory of the National Research and Innovation Agency (BRIN), both in Cibinong (6°29'30.9"S 106°50'43.3"E), as well as at the Cikeumeuh Greenhouse in Bogor, West Java. This work was conducted using two approaches: (1) identifying and isolating endophytic fungi from terrestrial orchid roots, and (2) testing selected isolates for their ability to stimulate plant development in Chinese cabbage and to reduce P. capsici infection in chili.
. 2.2. Sampling
The exploration and expedition research team of the National Research and Innovation Agency (BRIN) sampled terrestrial orchid species for this study in 2022, working in collaboration with resort staff from TNGGP for daily field assistants assigned to expedition sites. The roots of terrestrial orchid specimen samples were obtained from two resort locations within TNGGP, namely Cibodas Resort (GG) and Situ Gunung Resort (SIG), both located in Cipanas, Cianjur, West Java, Indonesia.
Sampling was carried out taking into account the natural population size of each orchid species. For species with large populations, many specimens were collected per population to ensure that the total number of individuals gathered did not exceed 20% of the population. The primary reference for species identification.was conducted using taxonomic references by Comber (1990) and O’Byrne (2011). The orchids were identified to the genus level based on vegetative morphology, while acknowledging the limitation imposed by the absence of reproductive structures. Root samples were collected and placed in plastic bags before being identified and documented. Identified plants were tagged and photographed. Sampling sites were recorded using GPS coordinates and marked with flagging tape. The collected samples were then taken to the laboratory for endophytic fungus isolation (Table 1; Figure 1). Orchid specimens were then returned to the Botanical Gardens for ex-situ conservation.
Table 1
Terrestrial orchid species collected in Mount Gede Pangrango National Park (TNGGP), West Java, Indonesia.

. 2.3. Isolation and purification of endophytic fungi
Prior to isolating endophytic fungi, root samples were obtained from healthy terrestrial orchid plants and surface sterilized according to the Narisawa et al. (2002) methodology. In brief, the roots were first thoroughly cleaned under running tap water to eliminate surface dirt. This was followed by three washes of 0.005% Tween 20 for three min each. The roots were then submerged in a 1% sodium hypochlorite (NaOCl) solution for 3 min before being rinsed three times with sterile distilled water to remove any leftover disinfectants. The sterilized roots were air-dried on sterile filter paper in Petri dishes, then sliced into 1–2 cm pieces with a sterile knife.
For fungal isolation, the sterile root segments were placed on corn meal agar (CMA) medium, which was made by combining 8.5 g of corn meal agar and 0.5 g of chloramphenicol in 1,000 mL of distilled water, supplemented with 25 g of agar. Each Petri dish was separated into four quadrants with sterile forceps. Two root segments were inserted in each quadrant, for a total of eight segments per dish. Each sample code had three replicates created. The inoculated Petri plates were sealed and incubated at room temperature (25 °C) for 7 days. Emerging fungal colonies were then subcultured to produce pure isolates. The hyphal tip approach was used to differentiate colonies with diverse morphologies. Fungal hyphae developing at the margin of the root segments were removed using sterile toothpicks and transferred to new CMA plates. The plates were sealed and incubated again for 7 days at room temperature. Following incubation, a macroscopic examination was performed. If colonies with differing morphologies persisted, further sub-culturing was performed until pure isolates were obtained. Pure endophytic fungal isolates were subsequently grouped based on macroscopic characteristics, including colony color (upper and lower surfaces), texture, and colony margin features.
. 2.4. Screening of endophytic fungi for plant growth-promoting potential and non-pathogenicity in vitro
Endophytic fungi were screened to identify isolates with plant growth-promoting potential and to ensure their non-pathogenicity to the model plant Chinese cabbage (Brassica rapa var. pekinensis) cv. ‘Siola‘ Hybrid F1. A total of 34 purified fungal isolates were cultured on oatmeal agar (OMA) medium, which was prepared by mixing 72.5 g of oatmeal agar powder, 1 g of MgSO₄, 1.5 g of KH₂PO₄, 1 g of NaNO₃, and 5 g of agar in 1,000 mL of distilled water. This medium was selected for its suitability in supporting the growth of endophytic fungi during in vivo testing. A completely randomized design (CRD) was adopted, with treatments containing distinct endophytic fungal isolates compared to controls, and replicated three times. Each fungal isolate was added to a glass bottle with OMA medium. The inoculated bottles were incubated at room temperature for 5 days to allow for sufficient fungal growth before testing.
. 2.4.1. Germination of Chinese cabbage seeds on water agar medium
Chinese cabbage seed germination was carried out on a water agar (WA) medium made by dissolving 7.5 g of agar powder in 500 mL of distilled water. Germination began on the second day of fungal incubation. Prior to sowing, Chinese cabbage seeds were surface sterilized by immersing them in 70% ethanol for 1 min, then treated with 1% sodium hypochlorite (NaOCl) for 5 min, and lastly rinsed three times with sterile distilled water. The sterilized seeds were dried on sterile filter paper and then sown on water agar, with about 20 seeds per Petri plate. This method was repeated in 8 dishes, yielding 160 seeds. The plates were incubated for three days, or until visible roots and shoots appeared.
. 2.4.2. Inoculation of Chinese cabbage seedlings
Chinese cabbage seedlings were inoculated with endophytic fungal cultures on the fifth day of fungal incubation. Three-day-old, germinated seedlings were aseptically transferred to sterile glass jars containing endophytic fungus isolates grown on OMA medium. Each jar got three seedlings, and each fungal isolate was tested three times. Control treatments were Chinese cabbage seedlings placed in jars containing uninoculated OMA media, with one control jar per replicate, for a total of three control jars. Jars were sealed and stored on a culture rack at room temperature (16–18 °C) under controlled light conditions (18 hours of darkness and 6 hours of light each day). The assay was carried out over 14 days, with seedling development monitored and recorded to determine if each endophytic fungal isolate promoted or inhibited plant growth.
. 2.4.3. Determination of fresh and dry weight of Chinese cabbage seedlings
After 14 days of incubation, the fresh (wet) and dry biomass of Chinese cabbage seedlings were measured. The seedlings were carefully removed from the culture vessels, and any fungal mycelia or residual media were gently wiped away. Fresh weight was instantly recorded using an analytical balance. The seedlings were placed in paper envelopes and dried in an oven at 60 °C for 72 h to determine their dry weight.
The growth of seedlings infected with endophytic fungi was compared to that of uninoculated controls. Fungal isolates were termed growth-promoting if they produced more dry weight in seedlings than controls while showing no obvious evidence of pathogenicity.
. 2.5. Screening of endophytic fungi as inhibitors of P. capsici in chili in vivo
Endophytic fungal isolates previously identified as growth promoters were tested for their ability to reduce P. capsici, the causal agent of root, stem, and foliar rot in red chili pepper, using in vivo methods. Chili plants infected with a specific endophytic fungus were compared to non-inoculated control plants.
Chili seeds cv. ‘Tanjung 2’ were seeded in 50 well seedling trays with rice husk charcoal as the germination medium. Each well was fertilized with AB Mix, and two seeds were put per hole. The tray was kept in appropriate circumstances for 21 days, with constant watering during the seedling period. Chili’s planting medium was a mixture of burnt rice husks, bran, and compost (1:2:7). This mixture was homogenized in a large container, and 500 g portions were placed in heat-resistant plastic bags. The medium was sterilized for two days using an autoclave at 121 °C and 1 atm pressure. To test growth, each 8 × 7 × 5.5 cm pot received 100 g of planting media (1:1 mixture of rice husk charcoal and field soil). An additional 10 g of the inoculated medium was incorporated into each pot, resulting in a final total of 110 g of planting substrate per pot. Each treatment was replicated three times, with four pots per replicate. Two chili seedlings were transplanted into each pot for subsequent observation and evaluation.
. 2.5.1. Inoculation of P. capsici and disease assessment
Endophytic fungal inoculum was created by scraping the surface of fungal colonies grown on solid media (Figure 2). Six mL of sterile distilled water was added to the culture, and the mycelia were homogenized by grinding with an additional 44 mL of distilled water, for a total volume of 60 mL. This fungal suspension was thoroughly mixed into the sterilized planting media and incubated for 24 hours to allow for initial fungal growth. Phytophthora capsici was inoculated into chili plants 35 days after planting. The pathogen suspension was created by crushing P. capsici cultures on CMA media and suspending them in 2 mL of sterile distilled water per culture plate. A 5 mL aliquot of this suspension, containing active zoospores, was applied to the planting medium in each pot using a macropipette to ensure uniform distribution.
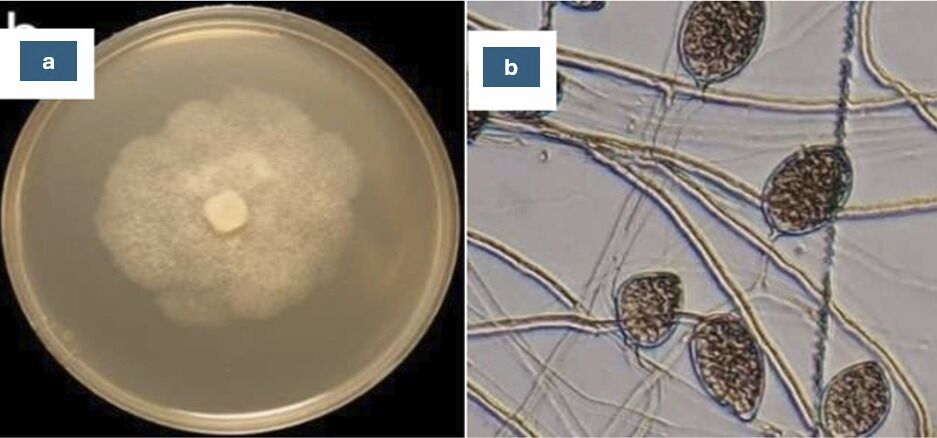
Figure 2
Phytophthora capsici pathogen isolated from chili plants: (a) macroscopic characteristic; (b) microscopic characteristic.
The parameters evaluated included disease incidence, disease severity, stem height, root length, fresh weight, and dry weight of both shoot and root tissues. The plant vigor index was determined using the formula of Abdul-Baki & Anderson (1973):
Disease progression was tracked every three days from day three to eighteen following inoculation. Disease severity was rated using a 0–5 scoring system, where: 0 = healthy plant, 1 = yellowing of leaves, 2 = slight necrosis, 3 = moderate stem necrosis with wilting, 4 = severe necrosis with pronounced wilting, 5 = plant death. The following formulas were used to compute disease incidence (DI) and severity (DS):
Disease Incidence:
Disease Severity:
where:
ni
vi
N
V
The biocontrol efficacy of endophytic fungi was determined by comparing the disease progression curve in treated plants with that in non-inoculated controls following the formula of cumulative area under the plant height progress curve (AUDPC) (Simko & Piepho, 2012):
. 2.6. Macroscopic and microscopic characterization of endophytic fungi
Selected endophytic fungi were described using both macroscopic and microscopic traits. Macroscopic observations included colony color (upper and reverse sides), surface texture, form, and margin characteristics. For microscopic examination, a small amount of fungal mycelium was collected using a sterile toothpick and transferred into a 0.2 mL microtube containing sterile distilled water. The suspension was gently stirred to prevent clumping, and two drops were placed on a glass slide. A coverslip was placed, and the slide was examined with an Olympus CX22 light microscope at 1000 × magnification. Morphological structures, including hyphae, spores, were detected and documented for future identification.
. 2.7. Molecular identification of growth-promoting endophytic fungi
Fungi were cultivated in potato dextrose broth (PDB) and incubated for 7 days at room temperature at 100 rpm using an orbital shaker. The resulting mycelia were collected, stored in sterile plastic, and mashed with a pestle to a fine consistency. Genomic DNA was isolated from superior endophytic fungus isolates following the protocol of the Plant Genomic DNA Mini Kit (Geneaid). DNA purity was determined using a NanoDrop 2000 spectrophotometer.
DNA amplification was carried out using the Polymerase Chain Reaction (PCR) with a Thermal Cycler, employing the universal fungal primers ITS1 (Forward: 5′-TCC GTA GGT GAA CCT GCG G-3′) and ITS4 (Reverse: 5′-TCC TCC GCT TAT TGA TAT GC-3′), as described by White et al., (1990). The PCR’s thermal cycling conditions were as follows: Initial denaturation at 94 °C for 5 min, 35 cycles of denaturation at 94 °C for 30 sec, annealing at 52 °C for 30 sec, and final extension at 72 °C for 7 min. Electrophoresis was performed at 100 V for 10 min, and the resulting DNA bands were visualized under UV light using a gel documentation system. A 1 kb DNA ladder was used as a molecular weight marker.
. 2.8. Data analysis
The sequence data were examined using the National Center for Biotechnology Information’s Basic Local Alignment Search Tool (BLAST). The retrieved ITS sequences were compared to the NCBI Nucleotide database to find the closest matches and establish the identity of the fungal isolates based on sequence similarity between the query and subject sequences.
The percentage of fungal isolates with growth-promoting activity and inhibitory effects against P. capsici was quantified using one-way analysis of variance (ANOVA) with SPSS software at a 95% confidence level. After detecting a significant effect, Duncan’s Multiple Range Test (DMRT) was used at a 5% significance level (α = 0.05) to compare treatment means.
. Results
. 3.1. Isolation of endophytic fungi from terrestrial orchid roots
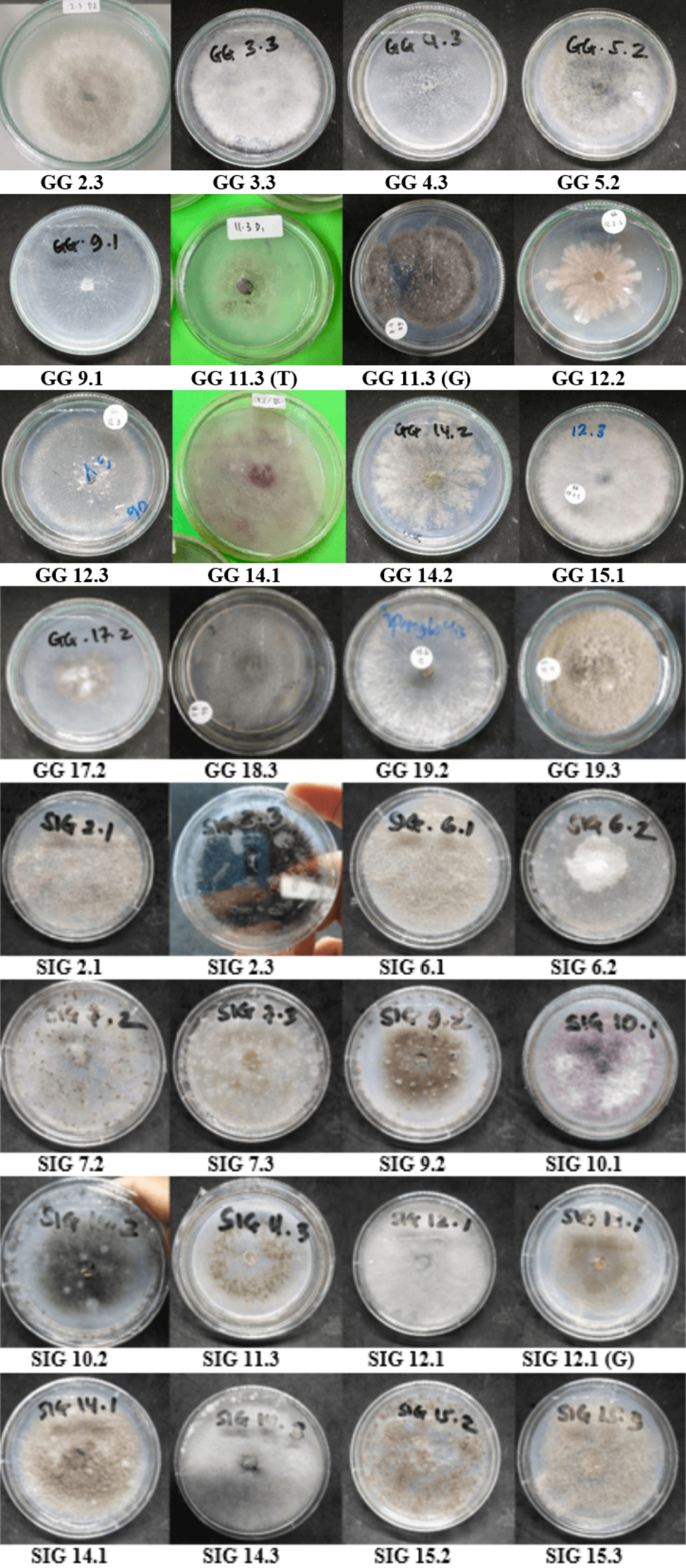
Endophytic fungi isolated from terrestrial orchid roots during field research were classified according to macroscopic characteristics (morphotypes), such as colony surface and reverse colors, texture, form, and margin features. Colonies with comparable physical characteristics were pooled, and one representative isolate was chosen from each group. Based on this method, a total of 34 different morphotypes were identified: 18 from Cibodas and 16 from Situ Gunung resorts (Table 2; Figure 3).
Table 2
The isolated codes of endophytic fungi from Cibodas and Situ Gunung Resort, TNGGP.
. 3.2. In vitro screening of growth-promoting and non-pathogenic endophytic fungi
In vitro screening of endophytic fungi for growth-promoting capacity and non-pathogenicity revealed three isolates that outperformed the 31 other isolates evaluated in terms of Chinese cabbage seedling development (Figure 4). These isolates are noted as GG 5.2, GG 14.2, and SIG 10.2. Over a 14-day observation period, Chinese cabbage seedlings treated with these three isolates grew vigorously, having fresh and green leaves throughout the experiment. The healthy look and continued growth of Chinese cabbage seedlings indicate that these endophytic fungi are not harmful and have a beneficial effect on plant development. Plant growth responses were measured in terms of seedling weight (Figure 5).
Figure 4
In vitro evaluation of endophytic fungi isolated from terrestrial orchids on the growth of Chinese cabbage seedlings: (a) Control (without endophytic fungi application); (b) Isolate GG 5.2; (c) Isolate GG 14.2; (d) Isolate SIG 10.2.

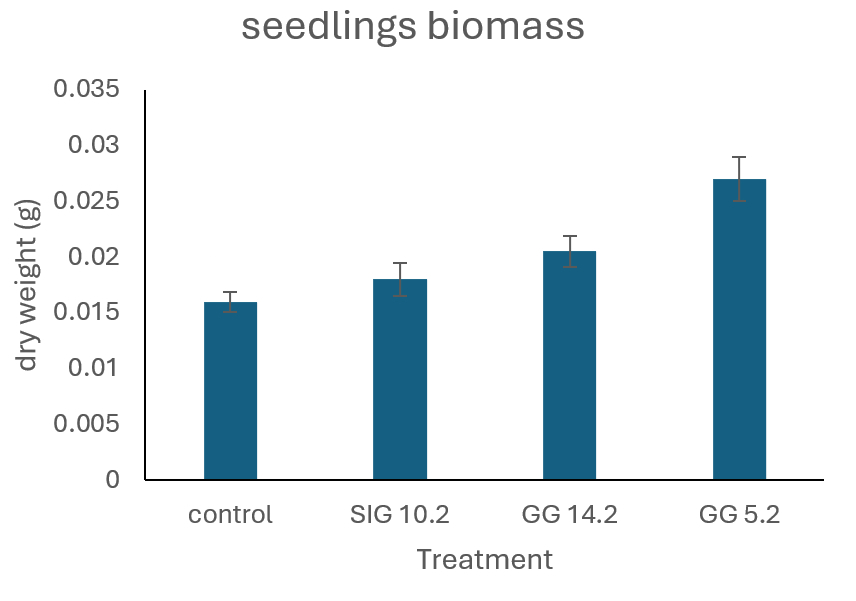
Dry weight measurements revealed that isolate GG 5.2 had the strongest growth-promoting impact when compared to isolates GG 14.2 and SIG 10.2. The control group had the lowest average dry weight of all treatments. Chinese cabbage treated with any of the endophytic fungal isolates consistently had a higher dry weight than the control. This rise in dry weight suggests that the use of endophytic fungi greatly improves Chinese cabbage growth.
. 3.3. Screening of endophytic fungi as biocontrol agents against P. capsici on chili in vivo
. 3.3.1. Effect of endophytic on the growth of chili seedlings: plant height and dry weight
The in vivo screening assessed the effectiveness of endophytic fungi in suppressing P. capsici and their impact on chili plant growth.
Figure 6A presents the mean average root length of chili plants across different treatments, while Figure 6B shows the corresponding stem height measurements. The data demonstrate notable differences in both root and shoot development, suggesting that the treatments had measurable effects on plant growth.
In Figure 6A, the mean root length of chili plants differed considerably between treatments. The treatment group(s) inoculated with beneficial bacteria or growth-promoting chemicals to fungi had significantly longer root lengths than the uninoculated control group. The increase in root length implies better root development, most likely due to greater nutrient uptake or hormonal stimulation caused by the treatment. Similarly, Figure 6B shows a constant trend in stem height. Plants given bioinoculant or biostimulant treatment had considerably taller stems than the control group. This rise in shoot height could be related to increased overall plant vigor and growth promotion, which could be the result of improved root system development that allows for greater water and nutrient uptake.
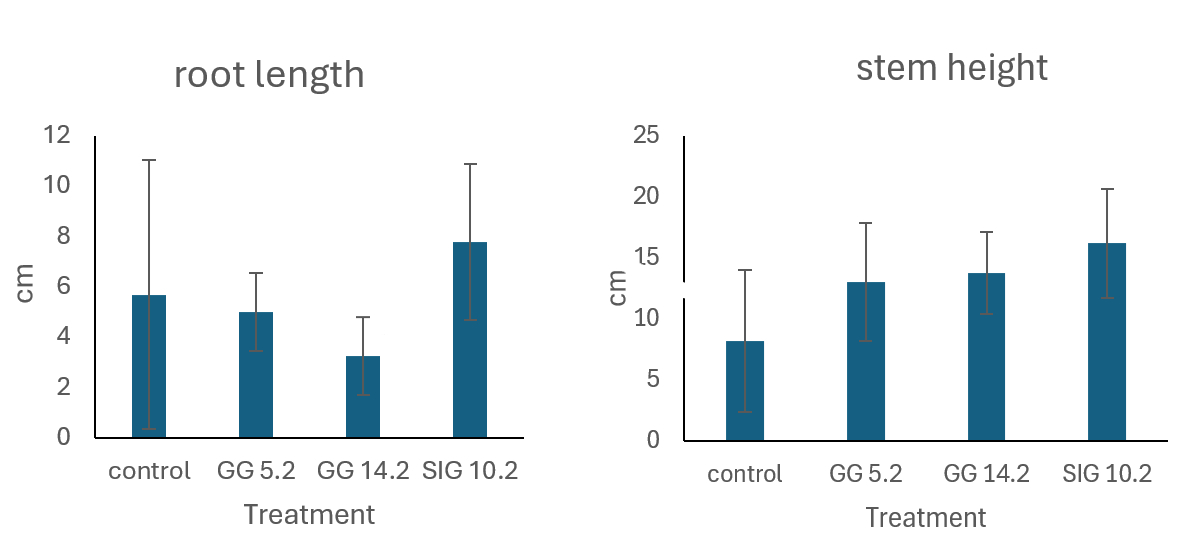
Table 3 depicts the effect of the treatments on the seedling vigor index (SVI) of chili plants. The findings clearly show that all examined endophytes increased seedling vigor in comparison to the uninoculated control. The control group had the lowest SVI value of 1384, which served as a baseline for comparison. Among the treated groups, the SIG 10.2 strain had the greatest impact, obtaining a substantially higher SVI of 2396, or a 73.1% rise over the control. This significant improvement implies that SIG 10.2 may have strong plant growth-promoting characteristics, such as phytohormone synthesis, greater nutrient absorption, or increased stress tolerance.
Table 3
Effect of endophytic on the seedlings’ vigor index of chili.
| Treatment | Seedlings vigor index (SVI) | Difference (%) |
|---|---|---|
| Control | 1384 | – |
| GG 5.2 | 1804 | 30.3 |
| GG 14.2 | 1699 | 22.7 |
| SIG 10.2 | 2396 | 73.1 |
The other treatments also had positive effects: GG 5.2 increased the SVI to 1804, a 30.3% improvement, and GG 14.2 increased the SVI to 1699, a 22.7% rise over the control. Although these levels are lower than those in SIG 10.2, they nevertheless show positive impacts on early seedling development.
The results revealed differences in the average dry weight of roots and stems across treatments. Plants treated with the endophytic fungus designated SIG 10.2 had the greatest average dry weight, at 0.023 g for roots and 0.075 g for stems. In comparison, the control group had the lowest average dry weights (0.007 g for roots and 0.028 g for stems). The treatments with endophytic fungi GG 14.2 and GG 5.2 also determined average dry weights higher than those of the untreated plants, albeit not showing significant differences from the control (Figure 7A-B).
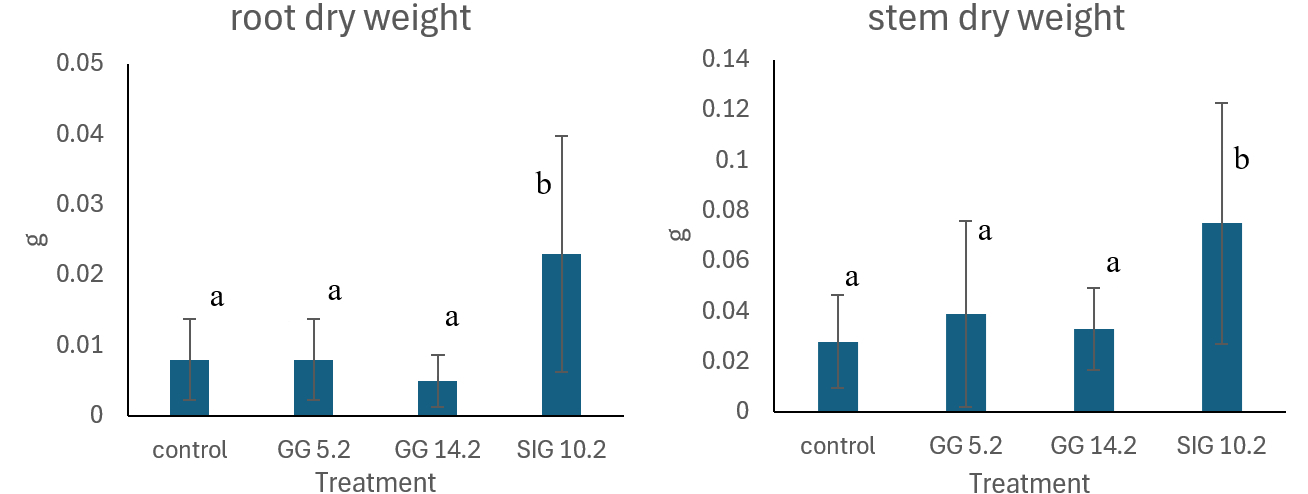
Statistical analysis using SPSS software and one-way ANOVA on root and stem dry weights gave p-values of 0.014 and 0.019, respectively. These values, which are less than the conventional significance level of 0.05, demonstrate substantial differences between the treatment groups and the control. In ANOVA, a p-value less than 0.05 means rejecting the null hypothesis (H0), suggesting the treatment factor significantly affects the measured parameters. The Duncan Multiple Range Test (DMRT) showed that the SIG 10.2 isolate outperformed other endophytic fungi treatments and the control, indicating superior efficacy in promoting growth and suppressing P. capsici infections.
. 3.3.2. Disease incidence and severity of P. capsici in chili
In vivo studies demonstrated substantial variations in disease incidence between chili plants treated with endophytic fungi and untreated controls (Figure 8A). The control group had the highest disease incidence, reaching 70.8% by day 3 post-inoculation, while all endophyte-treated groups kept incidences below 20%.
The disease incidence increased consistently throughout the monitoring period. By the end of the trial, the control group had the highest incidence (95.8%), indicating that the pathogen had infected virtually all of the plants.
Disease severity in chili plants differed significantly between treatment groups and controls. The control group had the highest severity levels, with 39% on 3 DAI reaching 90% on the last day (Figure 8B). Throughout the study period, disease severity in control plants was roughly twice that of chili plants treated with endophytic fungus. This suggests that chili plants that have not been treated with endophytic molds are extremely sensitive to P. capsici infection and do not grow.
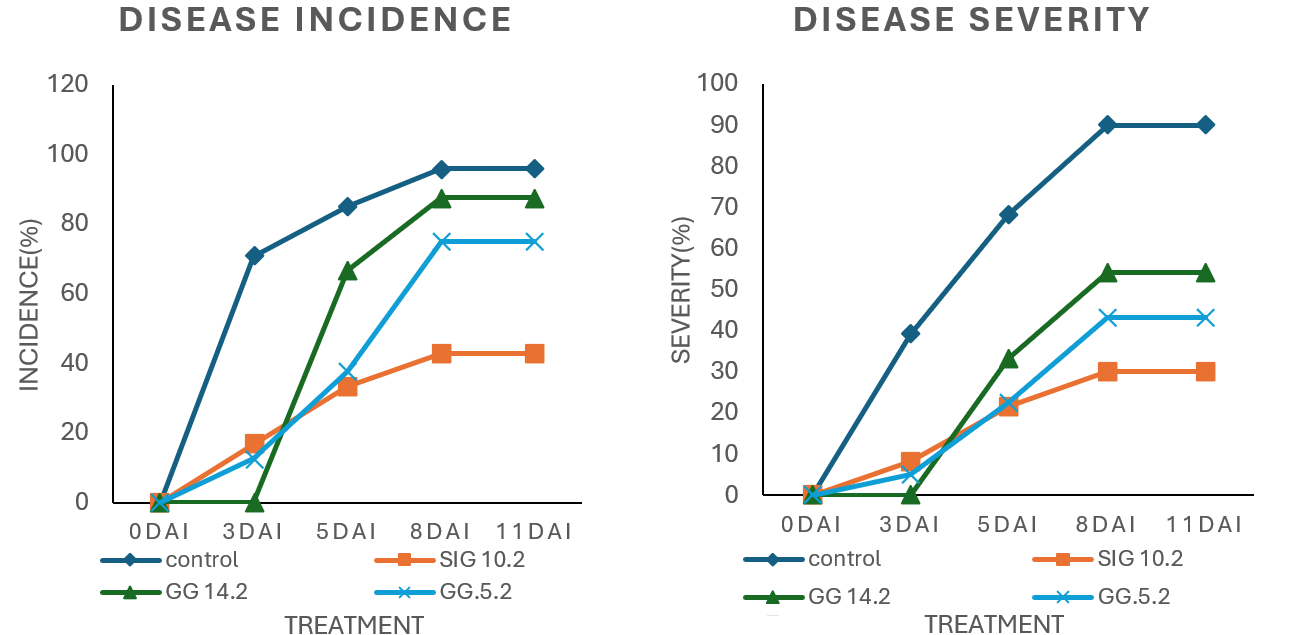
Among the treatment groups, chili plants inoculated with the endophytic isolate SIG 10.2 had the lowest disease severity, with 8.3% on 3 DAI, 21.7% on 5 DAI, and 30% on the last day of observation. In contrast, plants treated with isolate GG 14.2 showed a significant rise in disease severity from 0% on 3 DAI to 33.3% on 5 DAI, and 54.2% on 8 and 11 DAI. Although initially low, the severity increased dramatically during this time.
Older chili plants have greater defenses because their root systems are more established, and they produce more secondary metabolites. Nonetheless, when compared to the control, the use of endophytic fungi (particularly isolates SIG 10.2, GG 14.2, and GG 5.2) greatly reduced P. capsici infection. If GG 5.2, GG 14.2, and SIG 10.2 are novel isolates – especially if they are derived from terrestrial orchids or another underexplored plant – this represents a first report of their application against P. capsici in chilli.
The varying degrees of AUDPC reduction across the isolates (ranging from 52.21% to 68.86%) highlight the significance of selecting and optimizing certain endophytic strains for biocontrol. The endophytic isolates, particularly SIG 10.2, significantly reduced disease severity, with a 68.86% lower AUDPC than the untreated control (Table 4).
. 3.4. Macroscopic and microscopic characteristics of potential endophytic fungi
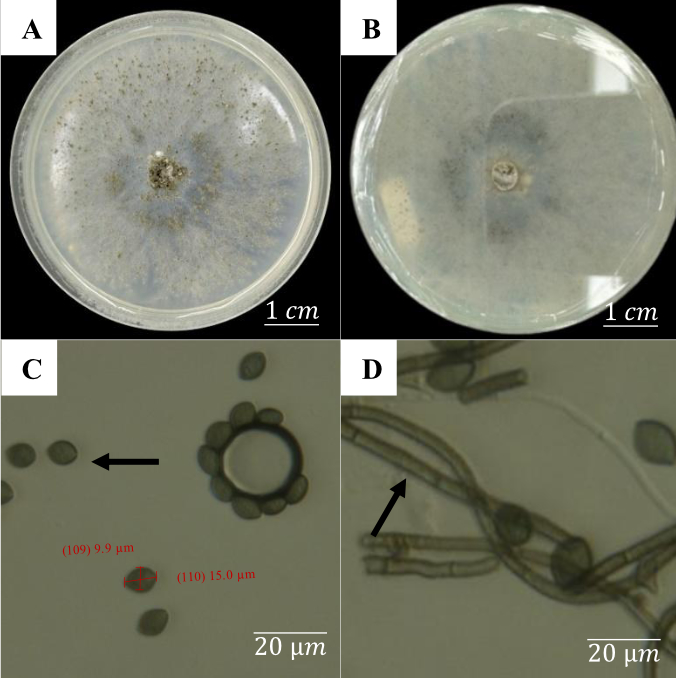
Figure 9 depicts the fungus Chaetomium globosum, which has peculiar macroscopic and microscopic characteristics. Macroscopically, the fungal colony appears white at first and gradually turns greenish gray on both the upper and lower surfaces over time. During growth on the medium, the colony develops an irregular, lobate border with a fibrous feel similar to cotton. Furthermore, water droplets accumulate atop this fibrous structure, creating the impression of crystal-like reflections when examined closely.
Microscopic inspection (Figure 9) revealed that C. globosum produces oval-shaped spores of approximately 15.0 µm by 9.9 µm. The hyphae are septate and found as thread-like structures. The spores are oval to spherical, brownish green in color, and measure around 15.0 µm × 9.9 µm.
Figure 9
Macroscopic and microscopic characteristics of Chaetomium globosum after 7 days of incubation: (A) Upper view of the colony; (B) Lower view of the colony; (C) Oval-shaped conidia; (D) Septate hyphae.
Colletotrichum gloeosporioides has different macroscopic and microscopic characteristics after 7 days of growth, as illustrated in Figure 10. Macroscopically, colonies are circular, with the surface color changing from grayish white to dark gray in the center by the sixth day. This fungus grows quickly, reaching a thickness of about 0.2 cm at 7 days after incubation.
Figure 10
Macroscopic and microscopic characteristics of C. gloeosporioides after 7 days of incubation: (A) Upper view of the colony; (B) Lower view of the colony; (C) Conidia; (D) Septate hyphae.
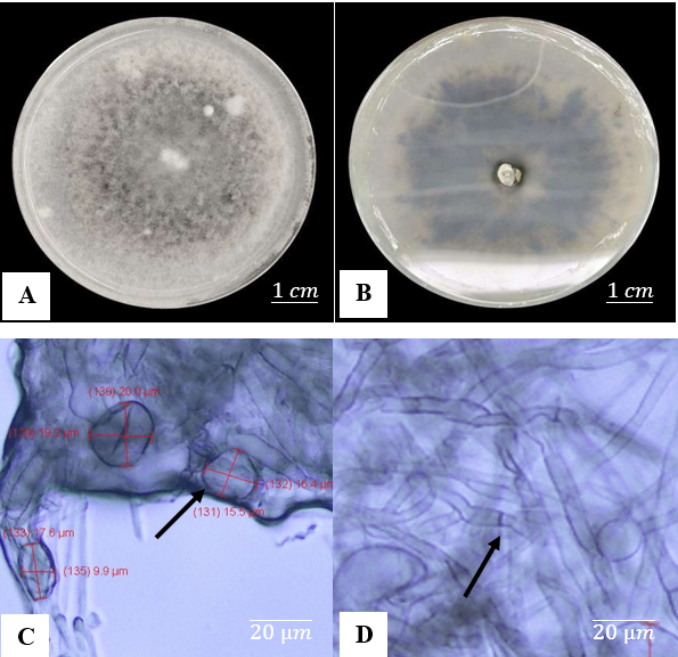
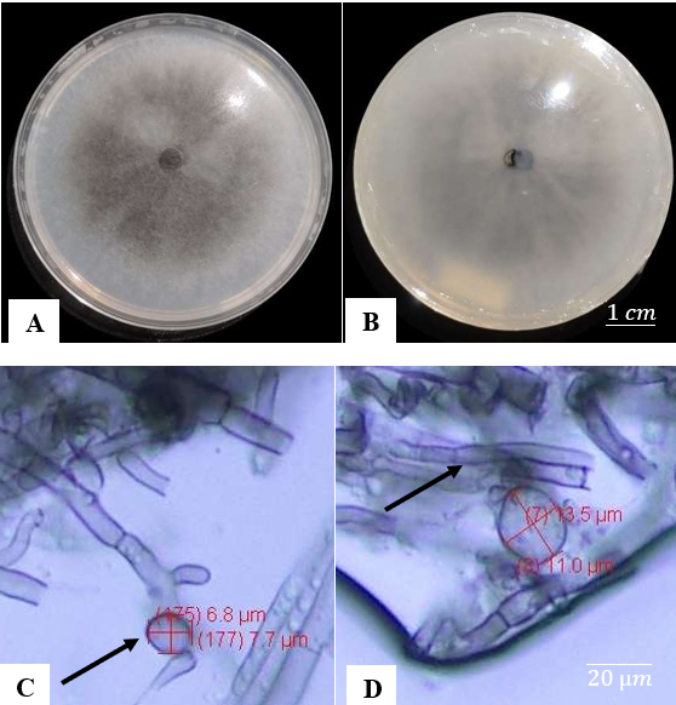
A microscopic examination reveals a layer of unevenly dispersed mycelium beneath the glass slide that covers the conidia and hyphae. This aggregation can interfere with the clear viewing of tiny details. Colletotrichum gloeosporioides conidia are cylindrical, with rounded ends and slightly tapering at the base, measuring 15–20 µm in length. The species produces conidia measuring 16.1 × 5.6 µm and grows at a pace of approximately 12.5 mm each day. Macroscopic examinations (Figure 11) reveal that C. arxii has a grayish surface with a thin, smooth, fiber-like texture on the upper side. The colony edges are complete, and the reverse has a similar gray hue to the top. Colletotrichum arxii grows at a slower rate than C. gloeosporioides and C. globosum, as shown by mycelium that has yet to completely cover the growing medium surface. Colletotrichum arxii conidia are spherical and tiny, ranging from 3 to 10 µm.
. 3.5. Molecular identification
BLAST analysis was used to identify potential endophytic fungi that promote growth and inhibit P. capsici. The BLAST analysis results based on the ITS rDNA region (Table 5) showed that isolate GG 14.2 has a very high genetic similarity to Chaetomium globosum, with a similarity percentage and query cover both at 100%. A high similarity percentage indicates substantial sequence identity between the query and database sequences, while a high query cover means that most of the query sequence aligns with the database entry.
Table 5
Identification of endophytic fungal isolates based on the ITS rDNA region.
For isolate GG 5.2, the ITS rDNA BLAST findings revealed a strong genetic similarity to C. gloeosporioides, with a query cover of 97.04%, indicating that the majority of the isolate’s sequences match this species. The extraordinarily low E-value of 0.0 verifies the statistical importance of this match, excluding coincidence.
The study provided by Liu et al. (2019) reported the most comprehensive molecular framework to date for Colletotrichum classification, setting a new standard for taxonomy and identification. They also clarified that, due to the lack of a universal barcode marker, species-level identification based solely on ITS sequencing should be considered provisional, especially within complex species groups such as C. gloeosporioides. Therefore, molecular identification was additionally conducted to verify and support the ITS-based identification of the pathogen.
. Discussion
Terrestrial orchids, including species from the genera Spathoglottis, Arundina, Arachnis, and Macodes, thrive in the highland tropical rainforests of Gunung Gede Pangrango National Park, especially at elevations above 900 meters above sea level. Due to their unique ecological adaptations and root morphologies, these orchids have dense endophytic communities, making them ideal for isolating beneficial fungi. Orchid root architecture, soil interactions, and habitat conditions, such as pH, aeration, and moisture, all play important roles in supporting different microbial symbionts (Herrera et al., 2022). Species-level identification of the orchid host is crucial for future studies and for advancing the understanding and application of associated endophytic fungi. Therefore, it is needed for detailed taxonomic identification in future research.
The observed disparities in endophytic fungus colonization levels between the two orchid collection sites, Cibodas Resort and Situ Gunung, can be explained in part by the diversity of host plant species present in each location. This is consistent with the previous findings that parameters such as host plant species, plant part sampled, and geographic location had a substantial impact on the diversity and abundance of endophytic fungi. Murphy et al. (2013) and Hardoim et al. (2015) stressed that successful endophyte colonization is dependent on a variety of factors, including plant genotype, tissue type, the associated microbial population, and both biotic and abiotic environmental circumstances. Chand et al. (2020) conducted a comprehensive study aimed at isolating, identifying, and functionally characterizing endophytic fungi associated with the roots of the wild Himalayan orchid Vanda cristata. The study emphasized the role of these endophytes in promoting plant growth, producing bioactive compounds, and enhancing orchid propagation. Endophytic colonization may occur through natural openings or plant wounds, whether induced by agronomic practices such as transplanting and hilling or formed naturally on roots. Once inside plant tissues, endophytic fungi can form mutualistic interactions that provide significant protection and growth benefits (Li et al., 2025).
Treatment with plant growth-promoting fungi (PGPF) may improve plant growth, enhancing biomass and development when compared to untreated controls. Bhullar (2024) classified an isolate as PGPF if it significantly improves one or more growth indices. This study indicates that the three studied isolates, particularly GG 5.2, as excellent growth stimulators for Chinese cabbage in vitro.
Several studies have found that endophytic fungi improve host plant tolerance to environmental stress by releasing phytohormones, which augment the plant’s defensive systems (Hiruma et al., 2016; Lubna et al., 2018). Shah et al. (2022) reported that colonization by non-mycorrhizal culturable endophytic fungi plays a crucial role in orchid growth promotion, primarily through increased IAA synthesis and improved nutrient assimilation. Phytohormones such as IAA, cytokinins, and gibberellins, particularly auxins, stimulate cell differentiation and elongation in host plant tissues, resulting in enhanced lateral root development and shoot length. These hormones help endophytic fungi by promoting plant growth, germination, reproduction, and resistance to both biotic and abiotic stressors (Egamberdieva et al., 2017). Among these, IAA is one of the most widely investigated metabolites found in endophytic fungi, demonstrating its importance in plant growth and defense mechanisms (Herrera-Parra et al., 2017). Shrestha et al. (2018) revealed that the roots of Dendrobium orchids serve as rich reservoirs of endophytic fungi with significant antibacterial potential. The study highlighted that the fungi are valuable sources of bioactive compounds, meriting further chemical and molecular characterization to identify specific metabolites responsible for the antimicrobial effects.
Endophytic fungi isolated from orchids show intriguing antibacterial capabilities. For instance, Khaterine & Sri Kasiamdari (2016) reported that fungal isolates from Phalaenopsis amabilis roots inhibited Fusarium oxysporum with inhibition rates up to 71.8%. This supports the premise that orchid-associated endophytes can be developed as biocontrol agents. Endophytic fungi help plants thrive in addition to suppressing pathogens. They can releasing phytohormones, increase nutritional absorption, and boost stress resilience. Dark Septate Endophytes (DSEs) are known to mineralize complex organic matter and aid in nutrient absorption in difficult environmental circumstances (Deng et al., 2020; Vergara et al., 2018). DSEs have also been shown to tolerate environmental stresses such as drought, salinity, and heavy metal contamination (Gaber et al., 2020; Gehring et al., 2020; Zhang et al., 2008).
Furthermore, the synergistic interaction of endophytic fungi and endophytic bacteria may increase their bioactivity. These bacterial symbionts can alter their fungal hosts’ metabolic output, including the synthesis of beneficial secondary metabolites (Vannini et al., 2016). Future studies should include detailed biochemical analyses to better elucidate the mechanisms underlying the plant growth-promoting and biocontrol effects of the isolated endophytes.
Endophytes’ potential to improve nutrient uptake, stress tolerance, and pathogen inhibition makes their use as biofertilizers and biopesticides extremely promising for sustainable agriculture. These benefits are especially important in chili farming because the crop is susceptible to both biotic and abiotic stresses (Pang et al., 2023; Zhai et al., 2016).
Symptoms of P. capsici infection differ depending on the affected plant section. Leaf infections appear as black, serrated patches, which are more visible when backlit. Even when the leaves are green, infected plants wilt and die, with the wilted leaves dangling before dropping off. Under humid conditions, the basal of diseased stems darkens and exudes a distinctive bluish fluid. Root infections are as severe, causing slow yellowing and leaf abscission, ultimately leading to plant death (Olmedo et al., 2020). The pathogen can cause total plant death within 10 days in susceptible cultivars. Meanwhile, in resistant varieties, the disease may still manifest as localized lesions on the secondary roots (Dunn & Smart, 2011).
Disease scoring of P. capsici-infected chili seedlings is an important approach for evaluating plant health, with two main factors to consider: disease incidence and severity. Disease incidence shows the percentage of plants affected, whereas severity assesses the level of infection on each plant, which is normally graded on a disease scale of 1 to 5. Plants with rot disease often develop dry stems and eventually perish. Rot symptoms in mature plants can extend to stems and fruits, causing considerable fruit loss at harvest. Disease severity is affected by environmental circumstances and plant age (Quesada-Ocampo et al., 2023). Disease incidence and severity can be modified by pathogen type, environmental factors, and host genetics.
The significant AUDPC reductions suggest that these endophytes could be useful as components in integrated pest management (IPM) systems, particularly in the development of biological control agents, which is consistent with worldwide trends toward sustainable agriculture and reduced chemical use. The substantial reduction in AUDPC values by endophytic isolates, especially SIG 10.2 (68.86%), demonstrates strong antagonistic activity and a high biocontrol against P. capsici. Such high performance is uncommon in endophyte-based disease management, underlining the possibility for use as biological control agent in crop protection strategy. To our knowledge, this is the first report of these specific endophytic strains being employed for disease suppression in chilli. Their efficacy not only underscores their biocontrol potential but also introduces a novel, eco-friendly avenue for managing P. capsici, supporting future integration into sustainable agricultural practices.
These findings are consistent with Harman (2011), who stated that endophytic fungi can restrict pathogen growth, reduce disease severity, stimulate plant growth, and improve nutrition by forming symbiotic relationships with host plants. Metabolites created during this mutualistic interaction contribute to the positive benefits. Furthermore, endophytic fungi benefit from residing within plant tissues, which allows them to avoid biotic stress and efficiently transport metabolites to the host.
Biological control utilizing endophytic fungi provides an environmentally benign alternative to chemical pesticides. Endophytes not only inhibit infections, but they also increase plant resistance by releasing bioactive and activating host defense mechanisms. For example, genera like Colletotrichum and Chaetomium, which were discovered in this study, are known to have significant antagonistic qualities that lead to increased plant health and growth. Chaetomium globosum is a filamentous fungus that lives in soil, plant debris, animal dung, straw and other cellulose-containing substrates, which is able to control fun gal pathogens primarily through (Linkies et al., 2021). Metabolites such chaetomin and chaetoviridins, as well as glucanolytic activity, have been demonstrated to be effective inhibitors of P. capsici (Kumar et al., 2020; Madbouly & Tamim, 2020). Other studies have confirmed that crude extracts from C. globosum effectively control Phytophthora species such as P. palmivora and P. nicotianae (Hung et al., 2015).
Furthermore, C. globosum metabolites influence the formation of pathogen pigments like indigoidine, which helps to restrict P. capsici growth and development. Aside from pathogen reduction, C. globosum and its metabolites enhance plant growth, as shown in citrus plants (Kumar et al., 2020). C. globosum produces phytohormones such as IAA and gibberellin, which boost host growth by increasing shoot elongation, biomass, leaf area, and chlorophyll content in comparison to controls. These characteristics identify C. globosum as a PGPF (Latif Khan et al., 2012). Therefore, through its diverse metabolites, C. globosum shows strong potential as an effective and comprehensive biocontrol agent against P. capsici and similar pathogens (Madbouly & Tamim, 2020).
Several endophytic fungi, including Cryptosporiopsis cf. quercina and Colletotrichum species, have shown efficiency against pathogens like P. capsici (Li et al., 2000; Lu et al., 2000). In field trials, endophytic fungal treatments reduced stem rot disease incidence in chili by 13.7–27.5 percent (Ramdan et al., 2018). This inhibitory effect is attributable to competition among fungi that share the same development environment. Both fungi struggle for limited space and nutrients, resulting in antagonistic relationships in which one fungus inhibits or kills the other by secreting inhibitory chemicals.
Colletotrichum gloeosporioides is identified macroscopically by white colonies with a cottony feel and uneven borders. Colletotrichum gloeosporioides has smaller spores that are also cylindrical (Ramdial et al., 2015; Weir et al., 2012). The hyphae are thick and septate, with a distinctive grayish hue (Ramdial et al., 2015). The colony’s underside is darker, almost black, and has the feel of a cotton ball. The colony margins are notably paler in color than the center.
Colletotrichum species are plant-pathogenic fungi that are known to cause significant damage to a variety of plants, including cereals, fruit trees, vegetables, and ornamentals (Masi et al., 2017). Colletotrichum gloeosporioides is the principal agent of anthracnose disease in chili plants, causing significant crop damage. According to Masi et al. (2017), Colletotrichum metabolites are often phytotoxins that are harmful to plants. Secondary metabolites from plant pathogenic fungi such as Colletotrichum are typically divided into two categories: host-specific toxins and non-host-specific toxins (Lou et al., 2013). Host-specific toxins exclusively affect the pathogen’s host plants, whereas non-host-specific toxins can harm a wide variety of plants, regardless of whether they are hosts.
Riga et al. (2019) studied the metabolites of C. gloeosporioides isolated from jackfruit (Artocarpus heterophyllus) leaves. Their research revealed that two weeks of cultivation were ideal for creating secondary metabolites, particularly from ethyl acetate extracts (EtOAc). Phytochemical analysis revealed that these extracts contained terpenoids, alkaloids, and phenolic compounds, all known for their diverse biological activities, including cytotoxic and antibacterial effects. These chemicals influence plant development through a variety of mechanisms. Terpenoids, for example, help plants defend against pests and pathogens, promote plant-to-plant communication by attracting pest predators or repelling competitive plants, and control root and leaf growth. Alkaloids also aid plant defense by repelling herbivores and diseases, facilitating plant-microbe interactions, and affecting root and leaf development. Phenolic chemicals are powerful antioxidants that protect plants from oxidative stress, aid in pest and disease defense, and influence plant-microbe interactions. Several phenolic compounds also influence root and leaf development (Srivasatava, 2022). Chapla et al. (2014) found that crude EtOAc extracts from Colletotrichum have modest antifungal efficacy against the phytopathogenic fungi Cladosporium cladosporioides and Cladosporium sphaerospermum. Furthermore, these extracts inhibited acetylcholinesterase (AChE) activity and had anticancer potential in Saccharomyces cerevisiae models.
To yet, no studies have specifically determined which metabolites from C. gloeosporioides can suppress Phytophthora species. However, it is now recognized that the fungal secondary metabolites serve critical roles in plant growth and development, notably root and leaf production. These metabolites may indirectly encourage plant development due to their complex activities.
BLAST allows for the investigation of phylogenetic similarity between species in the Genbank database. Values below 95% for either parameter may suggest the match lacks significant biological relevance (Pearson, 2013). Similarity and query cover values more than 95% in NCBI BLAST analysis are required to demonstrate the genetic relatedness between the studied sequences and the reference sequences in the database. Isolate SIG 10.2 has a high genetic similarity (98.33%) to C. arxii, with 99% query coverage, showing a robust alignment between the isolate and reference sequences in the database. The biological activity of secondary metabolites of C. arxii’s has not been studied extensively. Nonetheless, this research suggests that C. arxii has a unique ability to promote plant growth while suppressing pathogens such as P. capsici in chili plants. Although the precise metabolites responsible have yet to be determined, this discovery paves the way for future research on the secondary metabolites of C. arxii involved in plant growth promotion and pathogen control.
This is the first study to report the isolation and characterization of three endophytic fungi from terrestrial orchid plants, C. arxii (SIG 10.2), C. gloeosporioides (GG 5.2), and Chaetomium globosum (GG 14.2), that promote plant growth while suppressing P. capsici in chili under in vivo conditions. While these fungal species have previously been linked to positive plant-microbe interactions, their combined growth-promoting and biocontrol capability against P. capsici has not been demonstrated.
Among the isolates, SIG 10.2 (C. arxii) showed the greatest increase in plant biomass and disease suppression, establishing it as a novel and promising candidate for sustainable chili agriculture. The utilization of terrestrial orchids as a unique supply of endophytes fills a hitherto undiscovered ecological niche, emphasizing the research’s novel nature. In addition, there is a need for detailed taxonomic identification this isolate in future research.
Collectively, these findings underline the potential of using endophytic fungi, particularly in combination with other biocontrol agents like Trichoderma spp., as part of an integrated disease management (IDM) strategy against P. capsici.
Pant et al. (2017) stated that orchid endophytes represent a largely untapped biological resource with significant implications for plant growth promotion, biocontrol, and sustainable ecosystem management, and need to deeper exploration of these symbiotic relationships using modern molecular, biochemical, and ecological tools to fully harness their potential in agriculture and biotechnology.
More study is needed to identify the most beneficial strains, enhance application procedures, and understand the processes that underpin their protective benefits. Field validation, formulation development, and research into the induction of systemic resistance in plants should all be prioritized.
. Conclusion
This study successfully identified three endophytic fungal isolates from terrestrial orchid plants, i.e., SIG 10.2, GG 5.2, and GG 14.2, that demonstrated dual potential for promoting plant growth and suppressing P. capsici infection in chili under in vivo conditions. These three isolates were chosen from a group of 34 due to their superior performance in increasing plant biomass and lowering disease incidence and severity when compared to untreated controls. Among them, SIG 10.2 exhibited the most pronounced efficacy across all evaluated parameters, indicating its strong biocontrol and growth-promoting potential.
Molecular analysis revealed that the isolates include C. arxii (SIG 10.2), C. gloeosporioides (GG 5.2), and Chaetomium globosum (GG 14.2), three fungal species previously reported to have favorable features in plant-microbe interactions. However, the particular processes underlying their apparent effects, such as the generation of secondary metabolites, phytohormones, or enzymes, have yet to be determined.
Therefore, further investigations are warranted to characterize the bioactive compounds produced by these endophytes and to unravel the biological pathways involved in their interaction with both host plants and pathogens. Understanding these pathways is critical for optimizing their use in sustainable agriculture, especially as natural biostimulants and biocontrol agents. The insights gained from this study lay the groundwork for the development of eco-friendly strategies to enhance crop health and productivity.
Acknowledgments
We acknowledge funding to obtain resources of the isolates from’ Pendanaan Ekspedisi dan Eksplorasi 2022 (LPDP-BRIN), for a project titled: Eksplorasi dan koleksi keragaman jenis anggrek tanah dan mikroba simbionnya yang terdapat pada Taman Nasional Gunung Gede Pangrango (TNGGP) dan Gunung Salak Halimun (TNGHS)’. We also acknowledge the help and support provided by the staff of TNGGP during the collection of resources (orchids).
