. Introduction
Weeds are an inextricable component of field phytocoenoses. Their role varies, but they typically compete with crops for light, water, nutrients, and other habitat resources, which in most cases reduces yields. According to Oerke (2006), weeds account for nearly 34% of global crop yield losses. They are characterised by versatility, adaptability, and resilience to habitat conditions and cropping systems, which makes them difficult to control (MacLaren et al., 2020).
Crop rotation, through its diversity of crops and management practices, tends to increase weed diversity and reduce their competitiveness with crops (Chauhan et al., 2006; Garnier & Navas, 2012; Woźniak & Soroka, 2022). In contrast, reduced tillage practices and continuous cereal monoculture favour the specialisation of phytocoenoses and the predominance of a few species within weed communities (Hobbs et al., 2008; Gruber & Claupein, 2009). As noted by Neve et al. (2009), Hawes et al. (2009), and Garnier and Navas (2012), this results in agricultural landscapes dominated by a small number of difficult-to-control weeds, thereby diminishing biodiversity.
Agricultural practices, particularly soil tillage, influence the distribution of weed seeds in the soil and ultimately shape community composition (Tracy & Davis, 2009; Swanton et al., 2015; Feledyn-Szewczyk et al., 2020). No-till systems promote species with small seeds that germinate rapidly from the soil surface, whereas conventional tillage favours those emerging from deeper layers (Hernández Plaza et al., 2015). Consequently, no-till systems encourage the proliferation of wind-dispersed grasses (Feledyn-Szewczyk et al., 2020; MacLaren et al., 2020). Melander et al. (2008) and Woźniak (2023) also reported the highest density of Apera spica-venti in cereal monocultures under no-till systems.
Although herbicides remain effective for weed control, their use alters species composition and fosters the emergence of herbicide-resistant weeds (Mohler, 2001; Hicks et al., 2018; Bourgeois et al., 2019; Heap, 2020). Swanton and Weise (1991) argued that the most effective weed control relies on integrated weed management, combining a range of complementary methods, tools, and techniques. However, as emphasised by Mortensen et al. (2012), crop rotation remains central to integrated strategies, since crop succession and associated practices increase weed diversity while reducing competitiveness with crops (Liebman & Dick, 1993; Davis et al., 2012; Adeux et al., 2019; Weisberger et al., 2019).
Taken together, previous findings suggest that weed communities in crop rotations are more diverse than those in monocultures, with monocultures favouring the dominance of a few species. On this basis, the present study aimed to evaluate weed community diversity in a four-field crop rotation and in a 36-year winter wheat monoculture.
. Materials and methods
. Experimental site and design
The long-term field experiment was established in 1988 at the Uhrusk Experimental Farm (51°18'N, 23°36'E), belonging to the University of Life Sciences in Lublin, south-eastern Poland. Data for the present study were collected in 2024. The experiment was conducted with winter wheat (Triticum aestivum L.) as the test crop, cultivated under two cropping systems: (i) a crop rotation comprising potato (Solanum tuberosum L.) – winter durum wheat (Triticum turgidum L. subsp. durum (Desf.) van Slageren) – pea (Pisum sativum L.) – winter wheat, and (ii) a continuous 36-year monoculture of winter wheat.
The experiment was arranged in a randomised block design with three replications, with each plot measuring 25 m × 6 m. Winter wheat was grown under conventional tillage in both systems; in the crop rotation, it was sown following pea (Pisum sativum L.). The tillage sequence included shallow ploughing to a depth of 10 cm immediately after harvest in late July, followed by double harrowing (once directly after ploughing and again three weeks later), and pre-sowing ploughing at a depth of 18 cm in mid-September. These operations were performed using a stubble plough and a harrow. Winter wheat was sown in the last week of September at a density of 380 seeds m-2.
Prior to sowing, mineral fertilisers were applied at rates of 30 kg P ha-1, 85 kg K ha-1, and 20 kg Mg ha-1. Spring fertilisation involved nitrogen applied in three splits: 70 kg N ha-1 at tillering, 40 kg N ha-1 at stem elongation, and 20 kg N ha-1 at ear formation. Weed control relied exclusively on mechanical measures, namely harrowing at the tillering stage and again 7–10 days later. These treatments effectively eliminated a significant portion of short-lived weeds.
. Site characteristics
The experimental site lies within the Volyn Polesie macroregion and the Chełm Hills mesoregion (Kondracki, 2009; Pranagal & Woźniak, 2021). The area is characterised by heterogeneous soil cover and variable hydrological conditions (Dobrzański & Borowiec, 1961). According to the IUSS Working Group WRB (2022), the soil was classified as Rendzic Phaeozem, with a slightly alkaline reaction (pHKCL = 7.1). The soil texture comprised 52% of sand (2.0–0.05 mm), 25% of silt (0.05–0.002 mm), and 23% of clay (<0.002 mm). The soil was rich in available phosphorus (190 mg P kg-1) and potassium (200 mg K kg-1), with medium magnesium content (68 mg Mg kg-1). Total nitrogen amounted to 0.82 g N kg-1, and organic carbon content was 12.5 g C kg-1.
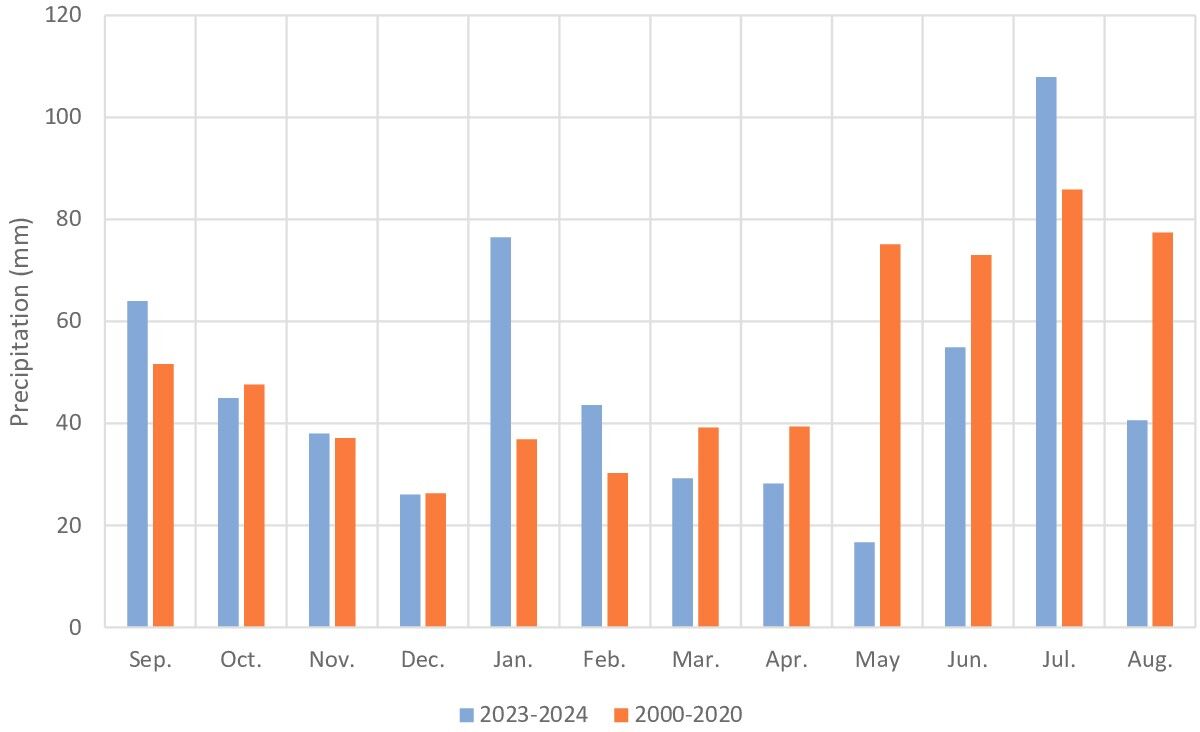
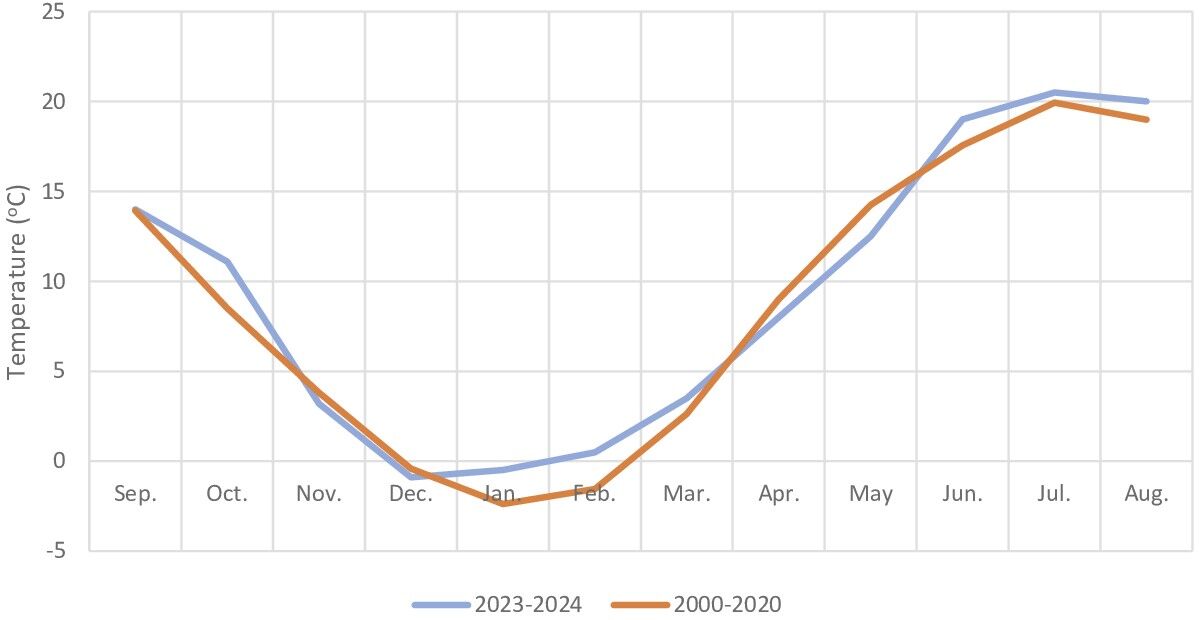
The growing season in this area lasts about 215 days and begins in the second half of March. In the 2023/2024 season (sowing of winter cereals in September 2023 and harvest in August 2024), total precipitation amounted to 571 mm, of which 312 mm fell between April and September (Figure 1). However, during the growing season, rainfall was lower than in the multi-year period 2000–2020. The warmest months were June to August, while the coldest were December to February (Figure 2). During the study period, the air temperature was similar to that in the multi-year period 2000–2020.
. Weed assessment and statistical analysis
Weed infestation was evaluated in both cropping systems using the following parameters: degrees of phytosociological constancy, percentage contribution of individual species in the community, Shannon–Wiener diversity index, Simpson dominance index, Sørensen similarity index, Jaccard similarity index, Bray–Curtis similarity index, evenness index (based on Shannon’s index), and species richness index.
Phytosociological surveys were conducted at the developmental stages of winter wheat: tillering (25 BBCH), stem elongation (32 BBCH), ear formation (55 BBCH), flowering (65 BBCH), milk maturity (75 BBCH), and full maturity (99 BBCH) (BBCH Working Group, 2001). The Braun-Blanquet (1964) method was applied, and syntaxonomic classification followed Matuszkiewicz (2022) and Soroka (2008).
Weed constancy degrees were assessed using the scale of Wilmanns (1993): V (80–100%), IV (60–80%), III (40–60%), II (20–40%), I (<20%).
The contribution of individual species (D) was calculated as:
The Shannon–Wiener diversity index (H’) was calculated as:
The Simpson dominance index (S) was calculated as:
The Sørensen similarity index (Sc) was computed using the following formula:
The Jaccard similarity index (Kj) was expressed as:
The Bray-Curtis similarity index (BC) was calculated using the following formula:
The evenness index (calculated for the Shannon index) was computed from the following formula:
The species richness index (d) was calculated as:
The results were analysed statistically using Statistica.pl software. Analysis of variance (ANOVA) was performed, and differences between mean values were tested with Tukey’s HSD test at p < 0.05.
. Results
The weed communities in both crop rotation and monoculture belonged to five syntaxonomic classes, with the most abundant taxa from Stellarietea mediae (Table 1). Within this class, Apera spica-venti, Avena fatua, Lamium purpureum, Fallopia convolvulus, and Papaver rhoeas reached the highest phytosociological constancy (degrees V and IV) in crop rotation, whereas A. spica-venti, A. fatua, Tripleurospermum inodorum, and Chenopodium album were the most frequent species in monoculture. In crop rotation, species of the class Molinio-Arrhenatheretea, typical of meadow flora communities, were also present. Taxa of Artemisietea vulgaris, characteristic of perennial ruderal habitats, occurred in both systems.
Table 1
Syntaxonomic classes and constancy degrees of weed communities in crop rotation and monoculture.
The weed community in winter wheat under crop rotation comprised 27 species, including 25 short-lived and two perennial species (Cirsium arvense and Elymus repens) (Table 2). The most abundant species were A. spica-venti and A. fatua, together accounting for 55.6% of the population. Galium aparine and F. convolvulus were also relatively frequent. By contrast, the monoculture community comprised 23 species (21 short-lived and two perennial) and was strongly dominated by A. spica-venti and A. fatua, which together represented 74.5% of the weed population. G. aparine and Anthemis arvensis also occurred in notable numbers.
Table 2
Percentage contribution of individual weed species to weed communities in crop rotation and monoculture.
Crop succession had a marked effect on diversity indices (Table 3). The Shannon–Wiener index was significantly higher in crop rotation (0.83) than in monoculture (0.62), indicating greater species diversity in the rotational system. Conversely, the higher Simpson index in monoculture (0.36 vs. 0.22) reflected the prevalence of a few dominant species.
Table 3
Shannon–Wiener diversity index and Simpson’s dominance index of weed communities in crop rotation and monoculture.
| Specification | Cropping systems | |
|---|---|---|
| Crop rotation | Monoculture | |
| Shannon‑Wiener’s Diversity Index (H’) | 0.83 a | 0.62 b |
| Simpson’s Dominance Index (S) | 0.22 b | 0.36 a |
The Sørensen, Jaccard, and Bray–Curtis indices (Table 4) indicated relatively low similarity between the communities in crop rotation and monoculture, mainly due to the strong dominance of a few taxa in the latter.
Table 4
Sørensen’s, Jaccard’s and Bray–Curtis’ similarity indices of the weed communities in the crop rotation and monoculture.
| Specification | Cropping systems | |
|---|---|---|
| Crop rotation | Monoculture | |
| Sørensen’s Similarity Coefficient (Sc) | 0.53 | |
| Jaccard’s Similarity Coefficient (Kj) | 0.38 | |
| Bray‑Curtis’ Similarity Index (BC) | 0.73 | |
Species distribution, assessed using the evenness index (Table 5), showed that higher values reflected balanced species proportions, whereas lower values indicated dominance. In this study, both the evenness and richness indices were significantly higher in crop rotation, supporting the conclusion that rotation fosters more balanced and diverse communities.
. Discussion
The structure of weed communities is strongly shaped by habitat conditions and farming practices (Soroka, 2008). Soil tillage, herbicide use, poorly diversified cropping systems, and monoculture frequently lead to dominance by one or a few species best adapted to prevailing conditions (Storkey et al., 2010; Bourgeois et al., 2019; Woźniak & Soroka, 2015). This pattern was clearly observed here, as Apera spica-venti and Avena fatua accounted for 74.5% of the weed population in winter wheat monoculture. Moreover, lower rainfall totals during the growing season than during the multi-year period (Figure 1) could have limited the occurrence of short-lived spring weeds.
Both occupy the upper canopy layer, exerting strong competitive pressure on the crop (Gaba et al., 2017; Woźniak & Soroka, 2022). By contrast, weeds beneath the canopy generally display lower competitive ability (Marshall et al., 2003; Storkey, 2006; Garnier & Navas, 2012), while fast-growing species typically outcompete smaller, slow-growing taxa.
Weed dynamics are also influenced by the soil seed bank, emergence timing, fecundity, and dispersal (Davis, 2017). In this study, 27 species were identified under rotation and 23 under monoculture. Although both systems contained typical cereal weeds, rotation also supported taxa associated with root crops and ruderal habitats, thereby enhancing floristic diversity. Similar results were reported by Clements et al. (1994) and MacLaren et al. (2020), who showed that rotation increases species richness while reducing competitive pressure on crops.
The diversity indices calculated here corroborate these patterns. The Shannon–Wiener and richness indices were significantly higher in rotation, confirming greater floristic diversity. In contrast, the higher Simpson index in monoculture reflected the predominance of a few well-adapted species. The evenness and similarity indices (Jaccard, Sørensen, Bray–Curtis) further highlighted differences between systems, with rotation supporting more balanced communities.
These results demonstrate that crop diversification plays a critical role in preventing the simplification of weed communities. By disrupting the life cycles of dominant species and creating heterogeneous ecological niches, rotation promotes biodiversity (Marshall et al., 2003; MacLaren et al., 2020). This is particularly relevant for sustainable agriculture, where maintaining species diversity enhances ecosystem resilience (MacLaren et al., 2020). Conversely, the prevalence of aggressive taxa in monocultures poses risks for weed management, as these species may evolve herbicide tolerance, posing persistent challenges (Storkey et al., 2010; Heap, 2020; Woźniak & Soroka, 2022). Our findings therefore provide further empirical evidence that crop rotation remains one of the most effective strategies for sustainable weed management and biodiversity conservation (Neve et al., 2009).
. Conclusion
In both cropping systems, weed communities were dominated by species of the class Stellarietea mediae, particularly Apera spica-venti and Avena fatua, with the dominance being especially pronounced in monoculture. Crop rotation supported greater species richness and floristic diversity, whereas monoculture was characterised by the prevalence of a few well-adapted species. Crop succession fostered more balanced and heterogeneous weed communities. Overall, the findings demonstrate that crop rotation plays a key role in maintaining biodiversity and ensuring sustainable weed management, while long-term monoculture promotes the homogenisation of weed flora.
