. Introduction
As a pivotal early branch in the phylogeny of land plants, liverworts offer unique insights into the evolutionary origins of terrestrial life on Earth. The genus Pellia, belonging to the family Pelliaceae, is a small yet widespread group of liverworts found in temperate and cool regions of the Northern Hemisphere (Schuster, 1992). Representatives of this genus are considered the most basal lineage of Jungermanniopsida because of numerous plesiomorphic traits, such as a wedge-shaped apical cell, a thallus lacking a midrib, a spherical capsule, and a massive seta (Schuster, 1992). However, this apparent simplicity has proven profoundly deceptive, masking a far more complex evolutionary history (Konstantinova et al., 2023; Schuster, 1981, 1992; Schütz et al., 2016).
The core challenge in studying Pellia has always been its high degree of phenotypic plasticity, where environmental factors induce significant variation in form, coupled with a low number of reliable morphological characters (Konstantinova et al., 2023; Schuster, 1981, 1992; Schütz et al., 2016; Zieliński, 1987). This combination has led to a long and often contentious taxonomic history, with boundaries between species frequently debated and redefined (Schuster, 1992). It has become increasingly clear that traditional morphological approaches are insufficient to resolve the true diversity and phylogenetic relationships within what was known as the Pellia sensu lato complex (Zieliński, 1987).
The advent of the molecular era has fundamentally reshaped our understanding of liverworts. The application of molecular phylogenetics has not only clarified species boundaries but has also revealed previously unimaginable levels of hidden diversity. The most significant outcome of this study is the division of traditional Pellia sensu lato into two distinct, well-supported genera: Pellia Hedw. (sensu stricto) and the newly recognized Apopellia Grolle. Furthermore, these studies have consistently uncovered widespread cryptic speciation, demonstrating that what was once considered a single species is often a complex of morphologically identical but genetically divergent lineages (Konstantinova et al., 2023; Schütz et al., 2016; Zieliński, 1987).
Beyond resolving these critical taxonomic puzzles, the Pellia complex is rapidly emerging as a valuable model system for addressing fundamental questions in molecular biology and evolution research. Its unique position in the plant tree of life makes it an ideal subject for comparative genomics, representing the broader understudied liverwort class, Jungermanniopsida. This is particularly important because Jungermanniopsida, with over 7,000 species (Söderström et al., 2016), is far more speciose than the well-studied Marchantiopsida, which comprises approximately 380 species, including the model organism Marchantia polymorpha (Bowman et al., 2017).
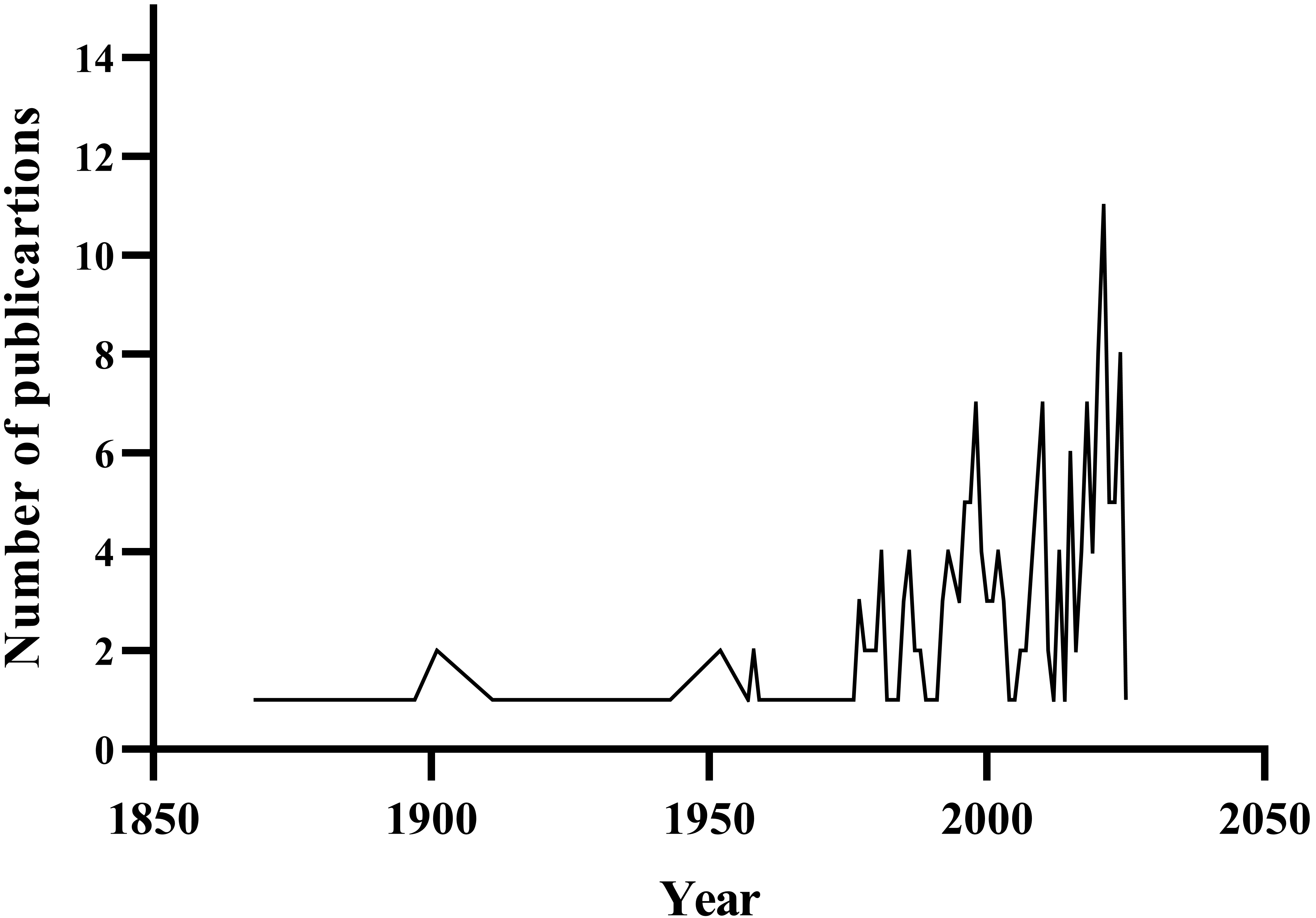
Scientific interest in the genus Pellia and order Pelliales has increased over the past few decades. An analysis of scientific publications featuring the keywords Pellia, Pelliales, or Apopellia reveals that research on this group was limited until the mid-20th century. Dynamic research development began in the 1970s, with notable intensification after 2000. The marked rise in studies over the last decade can be attributed to new systematic approaches, the use of molecular methods, and taxonomic revisions that led to the segregation of the genus, Apopellia. This trend suggests the growing importance of this plant group in research on the evolution, phylogeny, and biogeography of liverworts (Figure 1).
Figure 1
Bibliometric analysis of publication output concerning Pellia, Apopellia, and Pelliales (1850–2025). Data derived from Scopus database searches employing the query ‘pellia’ OR ‘apopellia’ OR ‘pelliales’. The temporal distribution demonstrates a pronounced upward trend commencing in the 1970s, with an accelerated publication frequency observed post-2000. The search was conducted on 05.04.2025.
This review synthesizes these recent and transformative developments. We traced the journey of Pellia from its confusing classical taxonomy to its modern reclassification based on robust molecular evidence. We detail the phylogenetic studies that led to the recognition of Apopellia, explore the biological and systematic implications of cryptic speciation within the complex, and finally, summarize the burgeoning field of molecular biological studies that are positioning these humble liverworts at the forefront of evolutionary research.
. Taxonomic history and classification
The earliest literary mentions of liverworts from the genus Pellia date back to 1616, when Colonna described them as Lichen alter minor caule calceato. The genus Pellia was formally established by Raddi in 1818, and the first critical taxonomic studies were conducted by Jack (1895). These foundational studies led to the distinction of three species: P. epiphylla, P. neesiana, and P. endiviifolia. Before 1876, three other described species were later synonymized: Pellia fabbroniana Raddi and Pellia calycina (Tayl.) Nees (both now considered synonyms of P. endiviifolia), and Pellia neesii (Lindb.) Limpr. (a variant of P. neesiana). Currently, the genus Pellia sensu lato comprises eight species. Five species belong to the genus Pellia (P. appalachiana, P. epiphylla, P. borealis, P. columbiana, and P. neesiana), while three belong to the genus Apopellia (A. alpicola, A. megaspora, and A. endiviifolia) (Söderström et al., 2013; Stotler & Crandall-Stotler, 2017).
Morphological separation of taxa within Pellia sensu lato has always posed a significant challenge. The simple thalloid morphology of these liverworts, combined with their vast distribution across the Northern Hemisphere, complicates their identification. This high structural simplicity can mask genetic differences, limiting their phenetic expression and resulting in a lack of clear diagnostic characters required for classical taxonomic classification. Consequently, a high degree of genetic variability is often concealed by structural homogeneity and significant phenotypic plasticity (Schuster, 1981).
Research initiated by Schuster (1981), encompassing studies on geographic distribution, cytogenetics, and genetic variability, led to the identification of several new species within Pellia sensu lato. These studies also laid the groundwork for the subsequent elevation of Apopellia from a subgenus to distinct genus.
The subgenus Apopellia Grolle, established in 1983, initially contained only Pellia endiviifolia. In 2016, Apopellia was elevated to the rank of genus based on detailed morphological and molecular analyses by Schütz et al. (2016), which demonstrated that the family Pelliaceae comprises two distinct, monophyletic lineages. This study provides a more accurate classification, offering greater insight into the phylogenetic relationships, ecological preferences, and geographical distribution of Pelliaceae species. Morphological data corroborated the genetic results, confirming distinct diagnostic characteristics for both Pellia and Apopellia (Table 1). Biogeographical analysis has further revealed different distribution patterns, with Pellia species being widespread in temperate regions, whereas Apopellia species show more restricted distributions (Schütz et al., 2016).
Table 1
Selected characters of the genera Apopellia (Grolle) Nebel & D. Quandt and Pellia Raddi. Based on Schuster (1981) and Schütz et al. (2016).
The genus Pellia is distinguished by several key features: a hemidiscoidal apical cell; clustered two-celled slime hairs on both the ventral and dorsal surfaces of the thallus, as well as on the calyptra, pseudoperianth, and at the base of the archegonia; thickening bands in the thallus cell walls; and papilliform cells in the male tubercle aperture (Table 1, Table S1). The archegonia are grouped and arranged horizontally or ascending. In contrast, the genus Apopellia is characterized by clustered multicellular slime hairs on the ventral side of the thallus, a ciliate pseudoperianth, and vertically grouped archegonia (Table 1; Table S1) (Schütz et al., 2016).
. Species of the genus Pellia
The genus Pellia (family Pelliaceae, order Pelliales) encompasses a small number of liverwort species characterized by simple thalloid gametophytes, specific anatomical features, and high phenotypic plasticity (Table 1). Representatives of this genus are found mainly in the temperate zone of the Northern Hemisphere, inhabiting moist terrestrial habitats with variable microclimates, such as forests, meadows, and stream and river banks. However, their elusive diagnostic features make identification challenging.
Species-level identification within Pellia can be problematic because of their morphological homogeneity and the frequent absence of reproductive structures in herbarium specimens (Schuster, 1981; Konstantinova et al., 2023). Historically, Pellia sensu lato included a larger number of taxa; however, the advent of molecular and phylogenetic studies has led to a significant revision. Consequently, the separate genus Apopellia was established, and the current concept of Pellia sensu stricto includes five species: P. epiphylla, P. borealis, P. neesiana, P. columbiana, and P. appalachiana (Schütz et al., 2016).
This subchapter aims to present the currently recognized species of Pellia, with a focus on their morphological features, geographical distribution, and ecological preferences. Where relevant, systematic difficulties associated with the delimitation of individual taxa are also discussed.
Pellia epiphylla (L.) Corda
The first species of this genus to be described, Pellia epiphylla, was originally named Jungermannia epiphylla by Linnaeus in his 1753 work, Species Plantarum. The species was later reclassified in 1829 by August Carl Joseph Corda, who placed it in the genus Pellia and established the currently accepted binomial nomenclature.
This species is characterized by a simple thallus that can reach over 1 cm in width and several centimeters in length (Figure 2). The thallus is typically green but may develop red or purple tints. Pellia epiphylla is a monoecious species with antheridia scattered along the midrib and a single archegonium at the apex, surrounded by a vertical, flap-like involucre. Monoiciousness and the structure of female sex organs are diagnostic characteristics that facilitate the taxonomic identification of this species during field observations. However, it is not uncommon to find large populations of Pellia that do not develop sexual organs, which makes it almost impossible to reliably identify them at the species level (Atherton et al., 2010).
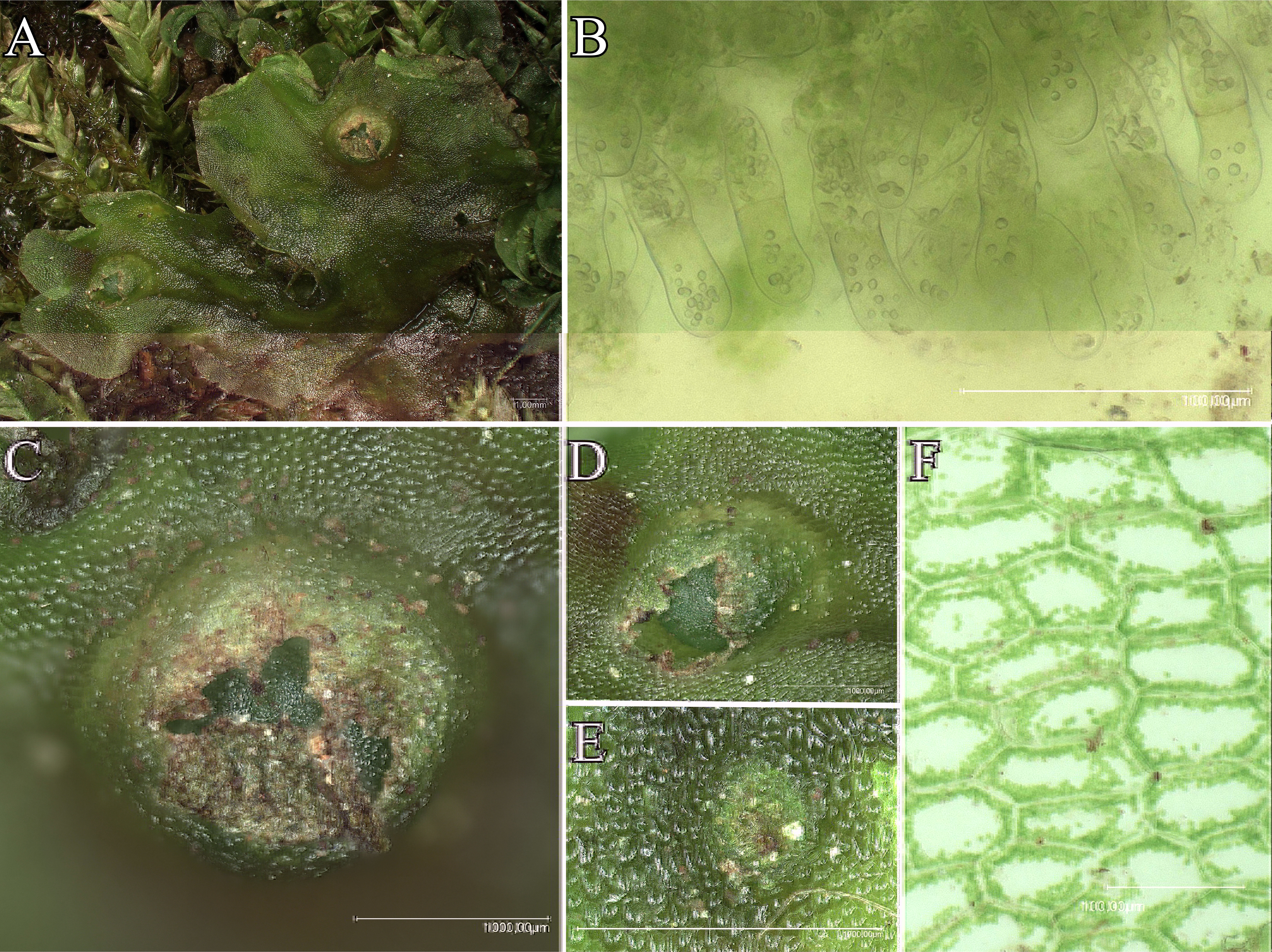
Figure 2
Pellia epiphylla: (A) General view of the thallus; (B) Slime hairs; (C, D) Archegonia on the thallus surface (close-up view); (E) Antheridium; (F) Thallus cells.
Pellia epiphylla is one of the most widely distributed species in the genus, with a nearly cosmopolitan distribution (Figure 3). It is found in Europe, Asia, North America, North Africa, the Azores, Madeira, and Greenland (Schütz et al., 2016). This species thrives in soils with varying degrees of moisture and acidity, including nutrient-poor substrates. It is commonly found on clay and sand and often grows on thin soil layers over rocks. Its habitats include the banks of streams, lakes, and marshes, as well as eroded banks, shaded forests, and disturbed areas such as ditches and wet paths (Schütz et al., 2016).
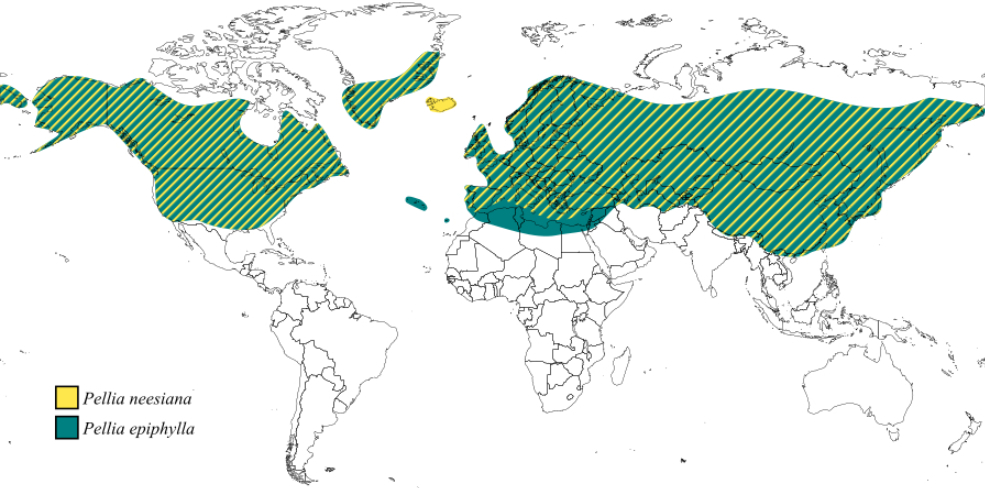
Modern molecular studies using RAPD techniques have revealed the existence of two cryptic species within the P. epiphylla complex, designated as species N and S (Zieliński, 1984, 1987). The distribution of these cryptic species has thus far been described only in Poland, where they exhibit an allopatric pattern: the N form is confirmed in the north, whereas the S form dominates in the south. Given the vast geographical range of P. epiphylla, further research is needed to determine the distribution of these cryptic lineages across the entire species range, as such data are currently unavailable.
Pellia neesiana (Gottsche) Limpr.
Pellia neesiana was first described by Gottsche in 1867 as a form of Pellia epiphylla. Although Limpricht (1877) elevated it to a separate species, its taxonomic status remained debatable until the early 20th century. Although Douin (1903) did not recognize P. neesiana as distinct, Frye and Clark (1937) later solidified its status as a separate species. In their study, they distinguished it based on key morphological features: a wide thallus (3–7 mm), dioicous sexual system, and an involucre forming a tube that is shorter on the side facing the thallus apex, with an exerted calyptra.
Pellia neesiana is characterized by a simple thallus, approximately 1 cm wide, which often develops red or purple pigmentation along the midrib. It can also be distinguished by its characteristic sharp odor. Despite their morphological similarity to Pellia epiphylla, the two species have different sexual systems. Pellia neesiana is dioicous, meaning that male and female reproductive structures develop on separate gametophytes. Consequently, field observations frequently reveal populations consisting of a single sex. The female involucre, which forms a tube that is shorter on its frontal side, is a key diagnostic feature (Atherton et al., 2010; Frye & Clark, 1937).
Pellia neesiana has a wide circumpolar distribution in Europe, Asia, North America, Greenland, and Iceland (Figure 3) (Schütz et al., 2016). It prefers moist to wet, nutrient-poor soils with a neutral to moderately acidic pH and clay or sandy-clay texture. This species tolerates seasonal flooding and is commonly found along streams, lakes, marshes, ditches, and wet paths, particularly within forests. It is occasionally found on gravelly forest tracks (Atherton et al., 2010; Schütz et al., 2016).
Pellia borealis Lorb.
Pellia borealis was first described by Lorbeer (1934) as a polyploid variant of Pellia epiphylla and immediately became the subject of extensive research on its origin, genetic structure, and taxonomic classification. Morphologically, it was distinguished from P. epiphylla by its larger gametophyte cells and higher average number of oil bodies. Additional diagnostic features include differences in the shapes of marginal thallus cells and epidermal capsule cells (Müller, 1954; Schuster, 1992). Comparative microstructural analyses have also revealed subtle differences in the distribution of papillose structures on the dorsal thallus surface and capsule surface cell patterns (Orzechowska et al., 2006).
The taxonomic status of P. borealis has long been debated because of its strong morphological similarity to P. epiphylla. Early cytological studies provided the first clues regarding its unique origin. Jachimsky (1935) discovered that P. borealis possesses a unique chromosome set in which it was difficult to identify the homologous pairs typical of autopolyploids. Based on this observation, he hypothesized that P. borealis was an allopolyploid that originated from a hybridization event between P. epiphylla and another, then-unknown Pellia species. Although Newton (1986b, 1986a) later confirmed the chromosomal differences between P. epiphylla and P. borealis, she could not definitively resolve whether its origin was auto- or allopolyploid, only excluding P. neesiana as a potential parent.
A breakthrough in understanding the origin of P. borealis came with the discovery of cryptic species within P. epiphylla. Isozyme analyses demonstrated that all analyzed samples of P. borealis exhibited heterozygous phenotypes that combined enzymatic variants characteristic of the two cryptic species, P. epiphylla S and P. epiphylla N. This provided strong evidence that they were the parental species (Odrzykoski et al., 1996). Subsequent research using molecular techniques, including the analysis of tRNAGly (UAA) gene intron sequences, confirmed this hybridization-based origin. In P. borealis, one copy of the sequence matched that of P. epiphylla species S, while the other matched P. epiphylla species N, proving its allopolyploid nature.
Pellia borealis is found in Ireland, Great Britain, France, Belgium, Germany, Poland, northern Russia, Norway, Sweden, and Finland. Its habitat preferences closely resemble those of P. epiphylla, indicating a significant overlap between their ecological niches (Figure 4) (Schütz et al., 2016).
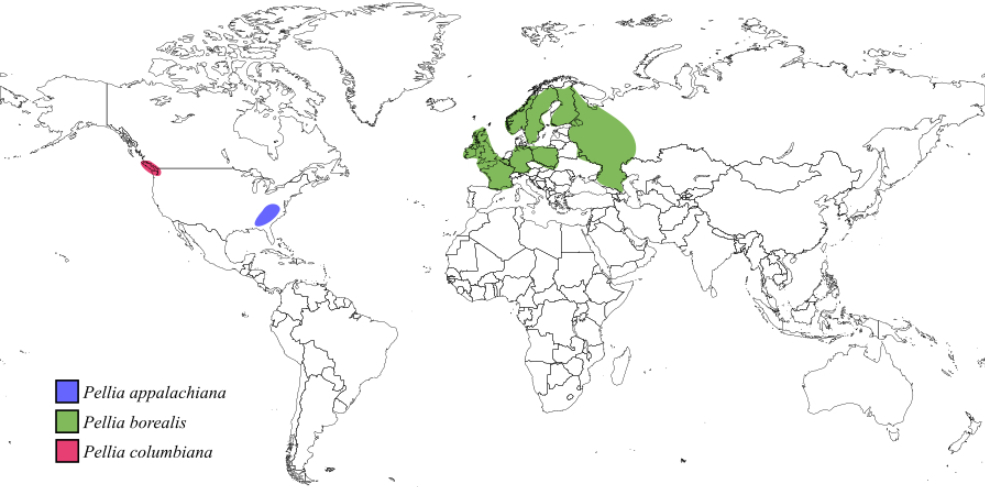
Pellia columbiana Krajina & Brayshaw
In the mid-20th century, Krajina and Brayshaw discovered a new liverwort species during field studies in British Columbia, which they named Pellia columbiana, and described it. This species is distinguished from other known Pellia taxa by a unique combination of morphological features, including thalli lacking thickening bands, spores larger than those of other Pellia species, an exerted tubular calyptra, and unilayered inner walls of the sporangium with semiannular and annular thickenings. The taxonomic status of Pellia columbiana has been changing over the course of research on this species.
It was initially reclassified as a taxonomic synonym of P. neesiana by Prosauker (1963). Later, Schuster (1981) redefined it as a subspecies, Pellia neesiana subsp. columbiana; however, this name was not validly published because the basionym was not cited. However, following comprehensive molecular and morphological analyses by Schütz et al. (2016), recent taxonomic treatments have recognized P. columbiana as a distinct species based on its distinguishing characteristics (Stotler & Crandall-Stotler, 2017).
Pellia columbiana is distributed mainly along the Pacific Northwest coast of North America, with confirmed locations including Vancouver in British Columbia (Canada) and Grays Harbor in Washington (USA) (Figure 4). It typically grows in forested areas along ditches and streams and prefers moist environments at elevations of 40–100 m above sea level. This species occurs abundantly near water bodies with a neutral or slightly acidic pH but does not tolerate very low pH conditions (Schütz et al., 2016).
Pellia appalachiana R.M. Schust. ex L. Söderstr., A. Hagborg & von Konrat
Pellia appalachiana was first discovered in southern Appalachia (Schuster, 1991). It exhibits morphological characteristics that are intermediate between those of the two existing species, Pellia neesiana and P. epiphylla. Like P. neesiana, it is dioecious, but its perichaetium – a posterior lamella that is absent anteriorly – resembles that of P. epiphylla. Schuster (1991) noted that P. appalachiana grew alongside both putative parent species, leading him to speculate that it might be a stabilized hybrid.
Reflecting this hypothesis, Schuster (1991, 1992) initially proposed the name Pellia X appalachiana. However, this designation was inconsistent with nomenclatural rules, leading to the later validation of the name Pellia appalachiana by Söderström et al. (2013). To date, there is no molecular evidence to support a hybrid origin for this species (Natcheva & Cronberg, 2004).
Pellia appalachiana is a thalloid liverwort that forms mats of overlapping thalli, sometimes arranged in rosettes. The thallus is up to 1 cm wide and several centimeters long, typically green, but capable of producing red pigments when exposed to direct sunlight. The midrib is not sharply demarcated and is “rather ill-defined” (Schuster, 1992). Pellia appalachiana is dioicous, with archegonia located on the dorsal surface of the thallus, protected by a posterior lobulate-incised involucre, antheridia sunken in dorsal pustules scattered or clustered along the thallus midline (Schuster, 1991).
Pellia appalachiana is an endemic species, with distribution limited to the Appalachian Mountains in the southeastern United States, including Alabama, Georgia, Kentucky, North Carolina, South Carolina, and Tennessee (Figure 4). It typically co-occurs with P. neesiana and P. epiphylla, suggesting that they share similar ecological preferences (Schütz et al., 2016).
. Species of the genus Apopellia
Species currently classified within the genus Apopellia were initially categorized within the genus Pellia, which belongs to the Pelliaceae family. This classification was primarily based on morphological characteristics, which were the fundamental criteria for distinguishing species in the 19th and early 20th centuries (Table 1). The subsequent elevation of Apopellia to genus rank was influenced by Schuster’s research from 1981, which provided the foundation for molecular studies that demonstrated its phylogenetic distinctiveness (Schütz et al., 2016). These seemingly simple organisms represent interesting evolutionary challenges. Pellia sensu lato abounds in evolutionary paradoxes: many taxa in this group show structural simplicity (such as Aneura pinguis), which is often mistakenly interpreted as primitiveness, yet they have remarkably wide geographic distributions. Some species appear to have slowed down or ceased evolutionary changes, especially unisexual taxa that reproduce asexually (such as Metzgeria leptoneura); however, two factors complicate this matter. 1) Structural simplicity may limit the visible phenetic expression of genetic differences in the species. The genus Pellia sensu lato can be highlighted as a case in which genetic variation exists but is “masked” by structural uniformity, making species difficult to distinguish when they are sterile. 2) Dramatic climate change and continental fragmentation over the past 65 million years have caused population isolation. Consequently, this has led to the elimination of “old” taxa in most of their range and the fragmentation of once continuous populations into genetically isolated units. Geographical isolation could have led to increased genetic discontinuity and, consequently, the differentiation of new species (Schuster, 1981).
Data presented in a series of papers on the Pellia endiviifolia complex (Krzakowa, 1981; Mendelak, 1981; Newton, 1981; Schuster, 1981) indicate that these factors influenced speciation within Pellia sensu lato; however, in this morphologically simple genus, genetic variation is “masked” by a high level of structural homogeneity (both within species and within the genus), combined with substantial phenetic plasticity. Based on these conclusions, Schuster indicated the distinctiveness of Pellia endiviifolia and Pellia megaspora, which provided the basis for the later differentiation of these species as a separate genus Apopellia – Apopellia endiviifolia and Apopellia megaspora. Currently, three species that were previously included in Pellia are classified under the genus Apopellia: A. endiviifolia, A. megaspora, and A. alpicola.
The aim of this chapter is to provide a synthetic overview of the current knowledge on species of the genus Apopellia, considering their diagnostic features, ecology, distribution, and systematic significance. Special attention is also given to issues related to taxonomic revisions.
Apopellia endiviifolia (Dicks.) Nebel & D. Quandt
Apopellia endiviifolia was first described by Dickson (1801) as Jungermannia endiviifolia. Over 180 years later, Grolle (1983) established the subgenus Apopellia to accommodate this species, emphasizing its unique morphological features that distinguish it from other taxa within Pellia. Subsequently, based on molecular and morphological evidence, Apopellia was elevated to the rank of genus, resulting in the current name Apopellia endiviifolia (Schütz et al., 2016).
This species has a green thallus up to 10 mm wide, which can develop red pigmentation when exposed to sunlight. A key diagnostic feature is the production of multiple apical proliferations (branches) in autumn, which can become so prolific that they obscure the maternal thallus (Figure 5). As a dioicous species, its antheridia are surrounded by a vertical, toothed, tubular involucre (Atherton et al., 2010).
Figure 5
Apopellia endiviifolia: (A) General view of the thallus; (B) Slime hairs; (C) Apical proliferation; (D) Archegonium; (E) Thallus cells.
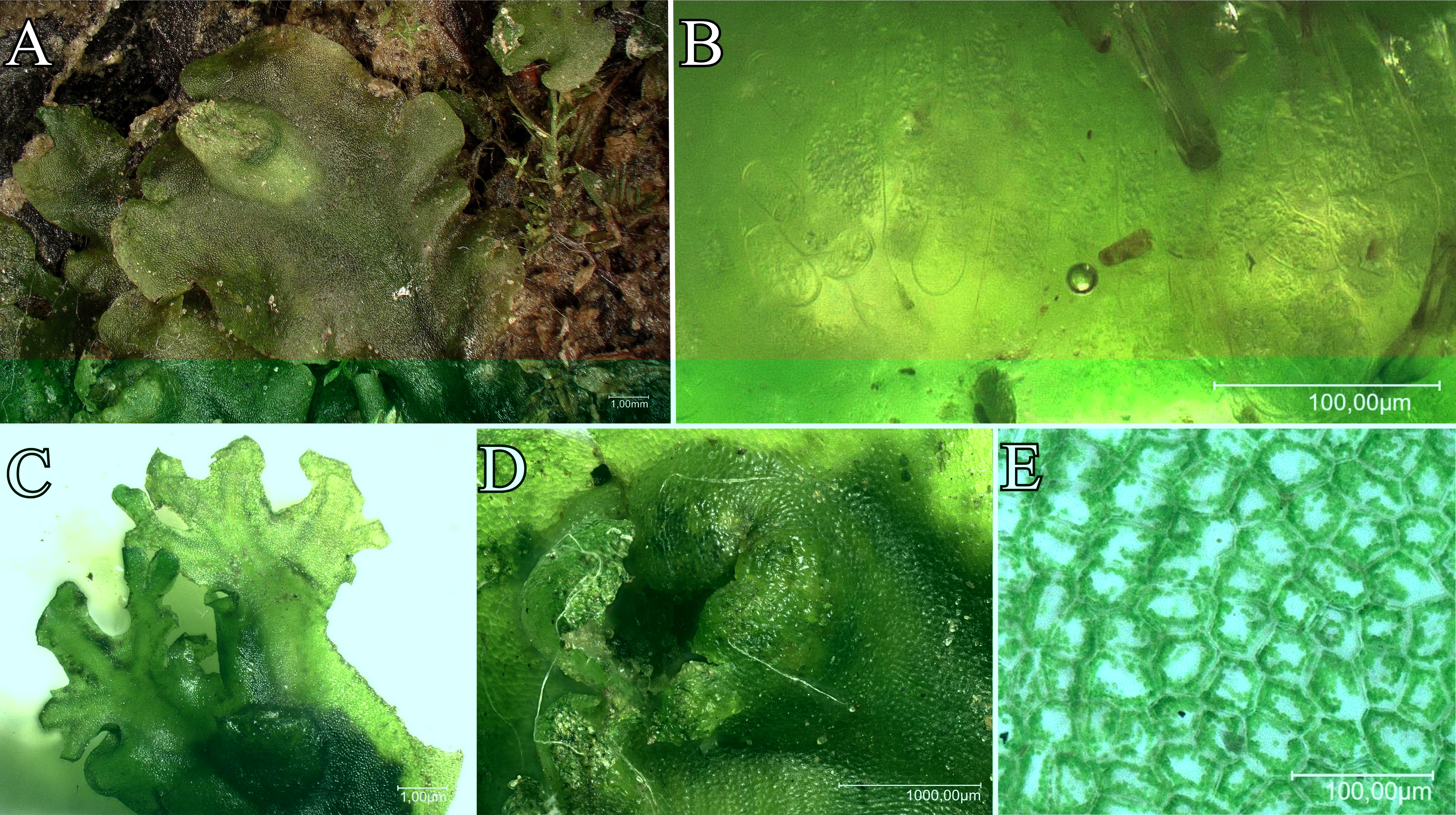
Apopellia endiviifolia is widely distributed in the temperate regions of the Northern Hemisphere (Figure 6). Its range extends from the lowlands of southern and western Europe to high mountains (2500 m in the Alps, 2300 m in the Caucasus) (Konstantinova et al., 2023). It is also found in the Mediterranean Basin, North Africa, southern Siberia, Central Asia, China, Korea, Japan, the Himalayas, and Oman, reaching as far north as 66°15' N in Scandinavia (Konstantinova et al., 2023).
Figure 6
Distribution of Apopellia endiviifolia, Apopellia alpicola, and Apopellia megaspora based on literature.
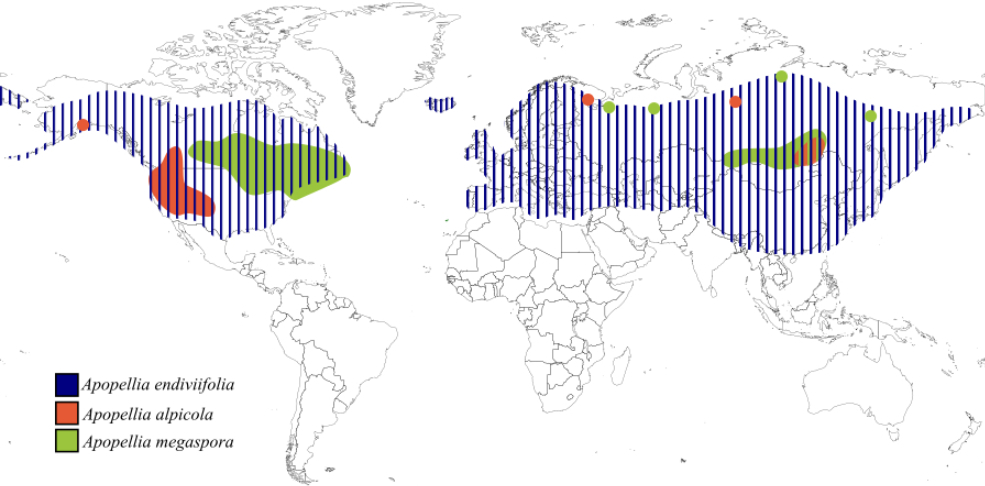
Ecologically, A. endiviifolia prefers nutrient-rich, calcareous, and clayey substrate. It frequently colonizes disturbed and anthropogenically altered environments, such as concrete canal banks, masonry around springs and damp path edges. It is also common in natural habitats, including stream banks, springs, and marshes, and can be found in forested areas along ditches and gravel roads (Schuster, 1981; Schütz et al., 2016).
Apopellia megaspora (R.M. Schust.) Nebel & D. Quandt
The description of Apopellia megaspora emerged from Schuster’s (1981) extensive research on the Pellia endiviifolia complex. Initial studies of populations from Europe, North America, and Japan suggested that American and European populations represent distinct evolutionary lineages.
This hypothesis was inspired by Schuster’s long-term field observations, spanning over 15 years, of a liverwort population along the Green River in Massachusetts, USA. He noted that these eastern North American plants consistently lacked the autumnal proliferations and red pigmentation characteristic of typical Apopellia endiviifolia. These observations prompted a collaborative, integrative study that led to the description of Pellia megaspora as a new species, supported by morphological, cytogenetic, and isoenzymatic evidence (Krzakowa, 1981; Mendelak, 1981; Newton, 1981; Schuster, 1981). A. megaspora is characterized by several key features: exceptionally large spores, lack of cell wall pigmentation, dioicous (unisexual) reproductive system, and a strongly incised and laciniate-dentate perichaetium mouth (Schuster, 1981).
Cytogenetic analyses provided further evidence of its distinctiveness. Although A. megaspora and A. endiviifolia share the same haploid chromosome number (n = 9), they differ significantly in karyotype symmetry and the presence of a sexually dimorphic ninth chromosome (m-chromosome) in A. endiviifolia (Newton, 1981). These findings were corroborated by isozyme analyses, which demonstrated clear biochemical differentiation (Krzakowa, 1981), and by detailed chromosome measurements that quantified the differences between the karyotypes of the two species (Mendelak, 1981).
Apopellia alpicola (R.M. Schust. ex L. Söderstr., A. Hagborg & von Konrat) Nebel & D. Quandt
Schuster (1992) first documented populations in the Canadian Cordillera at elevations of 2,134–2,896 m that exhibited morphological divergence from typical Apopellia endiviifolia. These high-elevation plants are characterized by their inability to form autumnal apical innovations, the presence of lower, frontally compressed juvenile gynoecia (similar to Pellia neesiana), and their exclusive occurrence in acidic regions. This montane taxon was initially described as Pellia endiviifolia subsp. alpicola Schust., a name that was later validated by Söderström et al. (2013). However, because a type specimen deposit was not designated, the name was not considered to be validly published (Stotler & Crandall-Stotler, 2017). The taxon was finally accepted at the species rank, as Apopellia alpicola, based on a comprehensive molecular phylogenetic study (Schütz et al., 2016), and its distinctiveness has been confirmed by subsequent phylogenetic studies.
The principal diagnostic features of Apopellia alpicola include unisexual thalli with short, angled to erect, laciniate involucres, which separate this species from other species of the genus; lack of thickening of cell walls in the mid thallus; and three or more cells long slime hairs (Schütz et al., 2016).
Apopellia alpicola shows subarctic-montane and nearly circumpolar distribution (Figure 6). Initially recorded in western Canada, it has been confirmed at high elevations in California (~2682 m), New Mexico (>3048 m), and Alaska (600–900 m) (Konstantinova et al., 2023; Schuster, 1992). It is also found in southern Siberia and the Murmansk region, extending beyond the Arctic Circle in the north. A. alpicola is scattered throughout Eurasia and appears widespread in western North America but hasn’t been found in eastern North America. (Konstantinova et al., 2023).
Apopellia alpicola develops in late snow areas, alpine meadows, and tundra vegetation in the far north. This species mainly occupies acidic environments and occurs primarily at elevations of 1800–2600 m above sea level in the alpine and subalpine regions. In the high Arctic latitudes, it occurs at lower elevations.
Apopellia alpicola has a subarctic montane and nearly circumpolar distribution. Initially recorded in western Canada, its presence has since been confirmed at high elevations in California (~2682 m), New Mexico (>3048 m), and Alaska (600–900 m) in North America (Schuster, 1992; Konstantinova et al., 2023). It is also found in southern Siberia and the Murmansk region, with its range extending beyond the Arctic Circle in the north. Although it is scattered throughout Eurasia and widespread in western North America, it has not been found in eastern North America (Konstantinova et al., 2023).
. Taxonomic challenges and revisions in the shade of cryptic speciation
The identification of Pellia sensu lato species poses a major challenge, as distinguishing them based on their morphological features presents many problems. The great similarity of thalli, caused by their morphological simplicity, makes species identification based on macroscopic features impossible when specimens are sterile. Identification issues stem from the high phenotypic plasticity of Pellia sensu lato, combined with the rarity of sporophyte production in the wild. This leads to difficulties in distinguishing Pellia from other morphologically similar genera such as Aneura. Identifying sterile thalli of Aneura and Pellia species can be challenging and often leads to misidentifications, particularly because their aquatic forms inhabit similar environments as each other. While microscopic observation of slime hairs on the ventral side of the apical thallus is considered the best method for distinguishing Pellia sensu lato species (Frahm, 2013), high phenotypic plasticity across wide geographical ranges and environmental variability introduce uncertainty. The significant number of misidentifications in herbarium specimens further highlights the difficulties in identifying Pellia sensu lato species (Konstantinova et al., 2023). Therefore, researchers have begun searching for alternative methods to enable the reliable identification of individual Pellia sensu lato species. The application of isoenzymatic markers is a response to this emerging need. Research on genetic variability has enabled the discovery and description of cryptic species of P. epiphylla and A. endiviifolia.
The first documented case of cryptic speciation within Pellia concerned Pellia epiphylla. Population studies conducted in Poland revealed the presence of two peroxidase phenotypes, which were initially considered to represent two distinct, previously unrecognized sibling species (Zieliński, 1984, 1987). These forms were characterized by allopatric distribution: the P. epiphylla N form dominated in the north of the country, whereas the P. epiphylla S form dominated in the south. The enzyme systems examined included peroxidases (PX), esterases (EST), aspartate aminotransferase (GOT), glutamate dehydrogenase (GDH), NADP-dependent malic dehydrogenase (ME), and acid phosphatases (AP). Subsequent genetic analyses showed significant differences between these two groups, with a genetic distance (Dn = 0.552) comparable to that typically found between species rather than populations of vascular plants (Zieliński, 1987). Zieliński (1987) proposed that these lines might represent separate biological species. This hypothesis laid the foundation for future genetic studies on cryptic speciation within only the genus name (Pellia) should be in italics. The presence of cryptic species in the Pellia epiphylla complex was confirmed through biochemical studies, which validated significant biochemical differences between these sister taxa. This strengthens the idea that geographical isolation and ecological differentiation are key factors driving cryptic speciation in Pellia (Prus-Głowacki et al., 1998).
With the increasing availability of molecular research methods, subsequent studies have consistently confirmed the existence of cryptic species within P. epiphylla. Molecular analyses of Pellia have utilized various genetic markers, enabling the differentiation of closely related cryptic species. Fiedorow and Szweykowska-Kulińska (1998) were the first to employ intergenic tRNA sequences as phylogenetic markers for Pellia species, using PCR to amplify the nuclear tRNALeu sequences. Simultaneously, the presence of distinct cryptic species was confirmed by studies using the Random Amplified Polymorphic DNA (RAPD) technique, which demonstrated significant genetic divergence within the P. epiphylla complex (Pacak et al., 1998). Phylogenetic studies of the Polish Pellia population using tRNALeuCAA intergenic sequences as molecular markers facilitated a more precise phylogenetic analysis of European Pellia species, including those from the P. epiphylla complex, thereby confirming the cryptic nature of P. epiphylla S and P. epiphylla N (Fiedorow et al., 2001). Szweykowska-Kulińska et al. (2002) introduced new chloroplast (cp) and mitochondrial (mt) tRNA molecular markers in their research: cp tRNALeuUAA, cp tRNAGlyUCC, cp tRNALysUUU, mt tRNASerGCU and mt coxIII. Their results indicated that tRNALeu intron sequences are identical in the closely related species P. borealis, P. epiphylla S, and P. epiphylla N and therefore cannot serve as markers for their differentiation. However, the remaining sequences studied successfully allowed for the distinction of closely related cryptic species, thus serving as highly effective marker sequences for differentiating cryptic species of P. epiphylla (Szweykowska-Kulińska et al., 2002).
Similarly, cryptic species have also been found for Apopellia endiviifolia. Studies on the Polish population of Pellia endiviifolia (now classified as Apopellia endiviifolia) distinguished two groups based on peroxidase variation analysis, which were described as “sister species” (Zieliński, 1987). They were named Pellia endiviifolia species A (water form) and Pellia endiviifolia species B (terrestrial form). These species exhibit differences in their range of occurrence and environmental preferences (Szweykowski et al., 1995; Zieliński, 1987). P. endiviifolia sp. A occurred at sites near water courses, shallow water bodies, and mountain springs, growing around the water surface line and in periodically flooded areas. In contrast, P. endiviifolia sp. B occurred on moderately moist substrates (Szweykowski et al., 1995). Phylogenetic studies of the Polish Pellia population using tRNALeuCAA molecular markers confirmed the occurrence of the cryptic species P. endiviifolia sp. A and sp. B, while also indicating the paraphyletic nature of these taxa (Fiedorow et al., 2001). Immunological analyses and studies of nuclear and chloroplast DNA have further confirmed their genetic distinctness, verifying the ecological and genetic separation of these forms (Polok et al., 2005; Sawicki et al., 2021).
Research on cryptic species within Pellia sensu lato has been conducted exclusively in Poland, which represents only a small fragment of the total distribution range (as discussed in the Taxonomic History and Classification section), which encompasses almost the entire Northern Hemisphere. Consequently, the distribution of cryptic species throughout their entire range remains unknown. A compelling parallel can be drawn from Aneura. In the case of Aneura pinguis, studies using DNA barcoding revealed that the nominally cosmopolitan A. pinguis was composed of 10 cryptic species, of which only five had been previously described (Bączkiewicz et al., 2017). A probable scenario is that Pellia sensu lato also conceals more cryptic species; however, it is essential to conduct analyses using material from a broader geographical range than has been previously studied. Such an analysis could reveal patterns indicating the habitat preferences of individual cryptic species. Habitat adaptation may be the main driving force of cryptic speciation within the taxon, as indicated by the aforementioned studies on Aneura pinguis (Bączkiewicz et al., 2017).
The description of cryptic species enabled the identification of the origin of the only described polyploid species in the genus Pellia: Pellia borealis. This species was identified as diploid (n = 18), with twice the number of chromosomes as P. epiphylla (n = 9) (Newton, 1986b, 1977). Since its initial description, its distinction as a separate species has been debated. The taxonomic distinctness of P. borealis has been revealed in genetic studies, which have shown differences in peroxidase genotypes and phenolic compounds, suggesting that P. borealis is not merely a polyploid variant but a distinct taxon (Krzakowa & Szweykowski, 1977; Szweykowski & Krzakowa, 1977). Two main hypotheses have emerged regarding its polyploidy: autopolyploidy (chromosome duplication within one species) and allopolyploidy (hybridization between two species followed by polyploidization). Early skepticism toward the allopolyploidy hypothesis stemmed from the lack of evidence for a second parent species and the absence of known hybridization mechanisms in liverworts (Mendelak, 1972).
Chromosome staining techniques have demonstrated chromosomal differences between Pellia epiphylla and Pellia borealis (Newton, 1986b). Pellia borealis chromosomes exhibit unique structural arrangements and heterochromatin distribution. Cytogenetic studies subsequently excluded P. neesiana as a potential parent species in the hypothesized allopolyploidy scenario. The final verification of the allopolyploidy hypothesis was made possible by the discovery of cryptic species within the Pellia epiphylla complex. The cryptic species P. epiphylla S and P. epiphylla N were identified as the parental species of P. borealis (Odrzykoski et al., 1996). Isoenzymatic analyses have demonstrated that all polyploid samples of P. borealis from various geographic populations exhibit heterozygous phenotypes, combining enzymatic variants characteristic of the parental cryptic species (Odrzykoski et al., 1996). Furthermore, molecular analyses of tRNA intron sequences provided evidence that P. epiphylla N is the maternal organism from which P. borealis inherited exclusively the chloroplast and mitochondrial genomes through uniparental organelle inheritance (Pacak & Szweykowska-Kulińska, 2000).
To further investigate the polyploid genome of Pellia borealis, newer cytological and molecular techniques have been employed, such as the Random Amplified Polymorphic DNA (RAPD) technique (Pacak et al., 1998). Recent phylogenetic studies have supported the hypothesis of an allopolyploid origin, confirming its status as a distinct species. Analysis of tRNAGlyUAA gene intron sequences (Fiedorow et al., 2001) allowed for the differentiation of cryptic species through the insertion/deletion of three nucleotides (GCT). In P. borealis, one copy of the sequence matched P. epiphylla S, and the other copy matched P. epiphylla N. This confirmed that P. borealis originated through hybridization between these two cryptic species (Fiedorow et al., 2001). Subsequent studies using the RAPD-tRNA marker further confirmed that Pellia borealis is a hybrid between the two cryptic species, P. epiphylla S and P. epiphylla N (Pacak et al., 2002). Comparative karyotype analysis of P. endiviifolia and P. borealis using fluorescent differential staining, C-banding, silver staining, and FISH provided cytological confirmation of the allopolyploidy of P. borealis (Orzechowska et al., 2010).
Modern molecular methods have enabled the taxonomic validation of Pellia species using many complementary approaches. The integration of isoenzymatic methods, cytogenetics, and classical botanical methods is crucial for the unambiguous acceptance of newly discovered species. This multifaceted approach is essential, as demonstrated by the case of Apopellia megaspora (formerly Pellia megaspora), which required years of observation to draw conclusions about its distinctiveness (Schuster, 1981). Cytogenetic analyses have provided detailed evidence for the distinctiveness of A. megaspora as a species. Although P. megaspora and P. endiviifolia have the same number of haploid chromosomes (n = 9), the ninth chromosome of P. megaspora does not show sexual dimorphism. Additionally, P. megaspora exhibits a more symmetrical karyotype with a predominance of acrocentric chromosomes, whereas P. endiviifolia shows greater chromosome asymmetry (Newton, 1981). These cytogenetic data were also confirmed through several methods: isoenzyme analysis (Krzakowa, 1981), which demonstrated biochemical differentiation; morphological analysis (Schuster, 1981), which documented structural differences; and detailed chromosome measurements (Mendelak, 1981), which quantified karyotypic differences. The distinction of A. megaspora as a separate species provided the basis for further research confirming the distinctiveness of Apopellia as a taxon.
. Phylogenetic relationship
One of the first comprehensive phylogenetic studies of the genus Pellia was initiated by Schütz et al. (2016), who in her paper conducted a detailed phylogenetic analysis based on morphological and molecular markers. The foundation for this research stemmed from earlier analyses, including those by Schuster (1981), who repeatedly pointed out the difficulties associated with correctly identifying species, especially when they are sterile. In their study, the authors posed several key questions: What are the phylogenetic relationships within the genus Pellia? Are the species within this genus monophyletic? Is there any evidence of infraspecific and geographical differentiation? In their study, the authors used 61 samples from the Pelliaceae family, collected from across each area of occurrence (Schütz et al., 2016).
The Pelliaceae split into two maximally supported lineages: Pellia and Apopellia (new rank proposed in that study) (Schütz et al., 2016). Analysis of three plastid loci (rpl16, rps4, and trnL-trnF) showed maximum support for previous reports regarding the distinctiveness of these two genera. The level of genetic separation between these two groups was greater than that observed within either clade. The results of the genetic analysis were strongly supported by morphological data, which indicated that each clade was distinguished by distinct, unique morphological characters. Furthermore, the division into two clear evolutionary lineages reflects the ecological patterns of each group, as Apopellia prefers to grow on calcareous substrates, whereas Pellia is associated with more acidic environments (Schütz et al., 2016).
Within the genus Pellia, the first clade includes P. epiphylla and P. borealis, whereas the second is composed of P. neesiana and P. appalachiana (Schütz et al., 2016). These species exhibit close phylogenetic relationships within their clades. Pellia borealis was classified as a variety of P. epiphylla because of the lack of sufficient genetic and morphological support for their distinctness. The results of this study indicate that Pellia epiphylla is a monophyletic taxon. The analysis showed that Pellia neesiana is a well-supported species and is sister to Pellia appalachiana (Schütz et al., 2016). The previous hypothesis regarding the origin of P. appalachiana as a hybrid between P. epiphylla and P. neesiana (Schuster, 1991) was not confirmed in this study. Hybridization between closely related species is possible; however, there is a high probability of overlooking this phenomenon because of the absence of diagnostic morphological characters (Schuster, 1992).
Within the genus Apopellia, two main clades were identified, corresponding to the species A. endiviifolia and A. megaspora (Schütz et al., 2016). The monophyletic A. endiviifolia was further differentiated into three subclades based on geographical origin: European (EU) and two Asian (A1 and A2). A. megaspora was divided into two subclades – eastern (E) and western (W) – a division which called its monophyly into question. Despite suggestions of paraphyly, the authors decided to maintain A. megaspora as a single species, arguing that a greater amount of genetic and morphological data were necessary to clearly differentiate these two clades (Schütz et al., 2016).
An integrative taxonomic study by Konstantinova et al. (Konstantinova et al., 2023) significantly supplemented and expanded our knowledge regarding the phylogenetic relationships of Pellia sensu lato and robustly supported the divergence of Apopellia as a distinct genus. Analyses were conducted based on two molecular markers: ITS1-2 nrDNA and trnL-F cpDNA. Sequencing of 36 new specimens from Alaska and Siberia, along with morphological analysis, provided new phylogenetic data and clarified the geographical distribution of the studied species. This work led to the description of the Apopellia megaspora W subclade as Apopellia alpicola (Konstantinova et al., 2023). According to this study, Pellia neesiana was divided into three subclades, which reflected the geographical distribution of the examined specimens (Konstantinova et al., 2023). This study also confirmed the occurrence of three subclades of A. endiviifolia.
The division into Pellia and Apopellia, initially revealed based on the analysis of nuclear and chloroplast genome regions (Konstantinova et al., 2023; Schütz et al., 2016), was subsequently confirmed by analyses of complete plastid and mitochondrial genomes (Paukszto et al., 2023). In these studies, 22 Pellia and Apopellia samples were subjected to barcoding analyses. Based on this work, the three subclades of A. endiviifolia – described as typical, aquatic, and spring forms – were recognized as monophyletic (Paukszto et al., 2023). Furthermore, the cryptic species P. epiphylla N (northern line) and P. epiphylla S (southern line) formed monophyletic, maximally supported groups that shared a common clade.
. Reproductive biology and sex determination
In an evolutionary context, sex determination in the genus Pellia sensu lato is particularly interesting because it encompasses both monoicous and dioicous species. In contrast to seed plants, dioicy is significantly more common in liverworts, with approximately two-thirds of all liverwort species being dioicous (Bell & Hemsley, 2000). A similar pattern is observed in the genus Pellia sensu lato, where the only monoicous species are P. epiphylla and P. borealis. The dioicous Pellia sl. species include: A. endiviifolia, A. megaspora, A. alpicola, P. neesiana, P. appalachiana and P. columbiana.
In bryophytes, dioicy is considered the primitive state, whereas monoicy is a secondary trait (Laenen et al., 2016; Longton & Schuster, 1983; Singh et al., 2023). For this reason, dioicous species can be viewed as evolutionarily older than forms that produce reproductive organs of both sexes on one individual. Within the genus Pellia sensu lato, heterogeneity in reproductive systems has been observed. In dioicous species, male and female thalli are phenotypically identical until the differentiation of sexual organs – antheridia and archegonia. The sexual organs of Pellia sensu lato are located on the dorsal surface of the costa of the main axis. Antheridia are colorless, immersed, and scattered in one to three irregular rows. The archegonia are solitary in cavities covered by a hemispherical tubercle with a small apical aperture (Paton, 1999).
Dioicous liverworts of the genus Pellia sensu lato encounter several obstacles that limit their reproductive success (Laenen et al., 2016). As in other bryophytes, sexual reproduction requires water, which means that increasing the distance between individuals of different sexes decreases the probability of fertilization. The distance of effective sexual reproduction in Pellia sensu lato is not well known, but it may not exceed 20 cm (Anand et al., 2013). The reproductive success of dioicous species may be further limited by two additional factors (Laenen et al., 2016): uneven sex distribution in populations, where one sex significantly predominates numerically (Bisang et al., 2014), and the tendency of males and females to colonize different habitats according to their distinct ecological requirements (sexually dimorphic niche partitioning) (Brzyski et al., 2014; Stark & McLetchie, 2006).
These obstacles result in rare sexual reproduction in dioicous bryophytes (Vanderpoorten & Goffinet, 2009). The sexual system has a decisive impact on reproductive success, with monoicous species characterized by a significantly higher frequency of successful fertilization and sporophyte formation than dioicous species (Longton, 1997). This difference is so pronounced that a significant proportion of dioicous moss and liverwort species have never been observed in the reproductive stage (Laenen et al., 2016). Among the liverworts of the genus Pellia sensu lato, vegetative reproduction clearly dominates, and the production of sexual organs occurs sporadically. For example, in P. neesiana, colonies consisting exclusively of thalli of one sex are frequently observed (Atherton et al., 2010). This pattern is consistent with the general tendency observed in dioicous species, where asexual reproduction constitutes an effective strategy for rapid population spread, whereas sporophyte production remains rare (Mishler & Newton, 1988).
The predominance of vegetative reproduction in Pellia sensu lato has been confirmed by observations of herbarium materials, where most specimens remain sterile, show only male characteristics, or contain unfertilized female structures (Schuster, 1981). A revision of Apopellia endiviifolia specimens by Konstantinova et al. (2023) demonstrated the scale of this phenomenon: among 100 examined specimens, only five had sporophytes, approximately 20 had pseudoperianths, and approximately 30 showed autumn proliferations. Furthermore, intensive vegetative reproduction was documented in populations of Pellia borealis (n = 18). Thalli growing in close proximity showed only one of two mutually exclusive peroxidase alleles, which is indicative of a clonal structure often extending over the entire colonies of P. borealis (Zieliński, 1986).
Although earlier studies indicated a strong correlation between dioicy and the production of specialized asexual reproductive structures (During, 2007; Hedderson & Longton, 1995; Longton & Schuster, 1983), contemporary phylogenetic analyses of mosses question this relationship (Crawford et al., 2009), pointing instead to other compensatory strategies. Dioicous species often achieve larger sizes than bisexual species and invest in intensive vegetative growth. This pattern, also known in angiosperms (De Jong, 2000), suggests that limitations in sexual reproduction are balanced by increased organism size and extended lifespan (Devos et al., 2011).
The occurrence of both monoicous and dioicous species within the genus Pellia creates broad possibilities for research on the mechanisms of sex determination, which has become the basis for intensive research on sex determination mechanisms in liverworts. The striking morphological similarity of these species, in contrast to their reproductive diversity, makes Pellia an exceptionally valuable model for understanding the evolution of sexuality in these plants.
Because liverworts are considered one of the oldest evolutionary lineages of currently living land plants, they can serve as a valuable model for studying the molecular foundations of mechanisms associated with sexual reproduction. However, knowledge of the molecular mechanisms of sex determination in liverworts remains modest. Currently, information on sex determination is available only for A. endiviifolia (Sierocka et al., 2011, 2020). Ultimately, these findings emphasize the need for further research to fully understand the complexity of the sex determination process and the development of monoecism in liverworts, using Pellia sensu lato as an example.
In the male gametophytes of Apopellia endiviifolia, Sierocka et al. (2011) identified four genes that were exclusively expressed: PenB_TUA1, PenB_Raba1/11, PenB_HMG-box, and PenB_MT. The expression of PenB_TUA1 and PenB_Raba1/11 was observed in male thalli, irrespective of whether antheridia were present, suggesting a role in the growth and development of general male gametophytes. In contrast, PenB_HMG-box and PenB_MT were expressed exclusively in male thalli producing antheridia and only under natural conditions. This indicates their involvement in reproductive organ development, which is influenced by environmental factors (Sierocka et al., 2011).
Using Representational Difference Analysis of cDNA (RDA-cDNA), Sierocka et al. (2014) identified three genes exclusively expressed in the female gametophytes of A. endiviifolia: PenB_CYSP, PenB_MT2, and PenB_MT3. These genes play roles in female gametophyte growth and differentiation, particularly in the formation of archegonia, the female reproductive organs. All three genes exhibited a tenfold higher expression in archegonial tissues than in vegetative tissues within female gametophytes grown in natural habitats (Sierocka et al., 2014).
A subsequent study by Sierocka et al. (2020) identified 87 genes with similar expression patterns during the reproductive phase of growth in Apopellia endiviifolia and Marchantia polymorpha. This suggests a common mechanism governing sex organ development in both species. Specifically, five female-specific and two male-specific Differentially Expressed Genes (DEGs) showed enriched expression in archegonia and antheridia, respectively. These DEGs included genes involved in various functions, such as ferric chelate reductase, potassium ion transporter, and chitinase class I (Sierocka et al., 2020).
. Transcriptomic analyses
Liverworts, among other bryophytes, represent one of the most ancient lineages of land plants. This makes them a valuable group for comparative genomic studies, allowing for a better understanding of plant evolution. Microtranscriptome studies provide crucial information on evolutionary connections among bryophytes and between bryophytes and their algal ancestors. Analysis of the bryophyte microtranscriptome suggests that the basic molecular frameworks for microRNA (miRNA) biogenesis, degradation, and function were established in the common ancestor of all land plants (Pietrykowska et al., 2022). Understanding these mechanisms is crucial for comprehending how plants adapt to life on land and how they develop complex regulatory networks based on miRNAs, which currently control their development and response to environmental stresses. Research on conserved miRNAs within bryophytes has indicated that six miRNA families conserved in all bryophytes likely appeared before the common ancestor of all land plants (Pietrykowska et al., 2022). These conserved miRNAs generally target conserved mRNAs, suggesting that they play important roles in regulatory networks throughout plant evolution.
Some miRNAs were shared between the liverwort species Marchantia polymorpha and Pellia endiviifolia. Analysis of microtranscriptomic data from these two species showed that miRNA Pen-miR8163 (P. endiviifolia) shares a similar sequence with miRNA Mpo-miR11737a (M. polymorpha). Furthermore, M. polymorpha contains a short RNA sequence similar to Pen-miR8170 miRNA from P. endiviifolia, and Mpo-miR11889 (M. polymorpha) is homologous to Pen-miR8185 (P. epiphylla) (Pietrykowska et al., 2022). The presence of these three miRNAs, shared by M. polymorpha and P. epiphylla, is highly significant, as it may indicate their important role in liverwort physiology. Conversely, the limited number of shared species-specific miRNAs supports the model of the frequent gain and loss of microRNA genes during evolution (Pietrykowska et al., 2022).
Research on Apopellia endiviifolia has shed new light on the evolutionary and functional aspects of miRNAs, challenging previous assumptions regarding their distribution and conservation in green algae and land plants. Earlier studies have suggested a lack of shared miRNA genes between these groups, as no evidence of overlap has been observed (Cuperus et al., 2011; Nozawa et al., 2012). Using Illumina-based next-generation sequencing, the identification and analysis of miRNAs in Apopellia endiviifolia enabled the discovery of hundreds of conserved plant miRNA families and the identification of new liverwort-specific miRNAs (Alaba et al., 2015). Analysis of the RNA degradome, which represents a collection of mRNA fragments cleaved by miRNAs, revealed that the target mRNA of only three miRNAs (miR160, miR166, and miR408) was conserved between liverworts and other land plants. This suggests that miRNA-mediated gene regulation has undergone significant diversification during the evolution of land plants.
Crucially, three miRNAs were identified in Apopellia endiviifolia with significant sequence similarity to miRNAs and small RNAs (sRNAs) found in the green alga Chlamydomonas reinhardtii. This discovery provides the first evidence for the existence of conservative miRNA species in different plant lineages and suggests a common evolutionary origin. The miRNAs in Pellia and C. reinhardtii were identified with a high degree of confidence, allowing for up to two mismatches in their sequences. One of these miRNAs (pen-miR1444) was previously described in algae, whereas the other two (pen-miR8183.2 and pen-miR8185) are novel miRNAs in Pellia that share homology with algal sRNAs. Experimental validation using northern hybridization confirmed their presence in Pellia, even when it was grown in controlled, contamination-free environments. These data strongly suggest that these miRNAs belong to a category common to liverworts and green algae, providing insight into the evolutionary relationship between land plants and their algal ancestors (Alaba et al., 2015). Broader studies of microtranscriptomes in other liverwort species are needed to enable a fuller understanding of the molecular mechanisms that facilitated the transition from aquatic to terrestrial ecosystems.
. Organellar genomes, RNA editing and nuclear genome structure
Technological development and increased access to molecular biology methods have enabled the expansion of research on Pellia sensu lato to include the study of organellar genomes, plastid and mitochondrial. Thanks to these advances, it has become possible to sequence the complete organellar genomes of Pellia sensu lato representatives. The first complete plastid genome to be described was that of Apopellia endiviifolia, with a size of 120,546 bp (Grosche, 2012). The A. endiviifolia chloroplast genome is the largest known chloroplast genome of Pellia sensu lato (Table 2). In subsequent years, the chloroplast and mitochondrial genomes of Pellia neesiana, Pellia borealis, and cryptic species belonging to the Apopellia endiviifolia and Pellia epiphylla complexes were also described (Paukszto et al., 2023).
Table 2
Size of chloroplast and mitochondrial genomes (based on Paukszto et al.,2023).
Among Pellia sensu lato species, distinct differences exist in mitochondrial genome sizes, ranging from the smallest mitogenome in Apopellia endiviifolia with a size of 109,389 bp to 163,996 bp in Pellia epiphylla (Table 2). Despite its relatively small size, the A. endiviifolia mitochondrial genome contains all protein-coding genes and the full intron set known from other Jungermanniopsida, including 41 protein-coding genes, three rRNAs, and 26 tRNAs (Sawicki, 2023). Flow cytometry analyses of genome sizes in species of the genus Pellia sensu lato have also revealed differences in their dimensions (Temsch et al., 2010), which may reflect differences in the amount of repetitive DNA and the accumulation of non-coding sequences.
Comparative organellar genomics can serve as a valuable tool for delimiting Pellia and Apopellia species, including cryptic species, using organellar genomes as ‘superbarcodes’. The identification of Molecular Diagnostic Characters (MDCs) in the mitogenome and plastome allowed for the successful identification of most evolutionary lineages of Pellidae, with the exception of the allopolyploid P. borealis. The mitogenome was the most effective for distinguishing species within the P. epiphylla complex, whereas the plastome was more effective for the A. endiviifolia complex (Paukszto et al., 2023).
The availability of complete organellar genome sequences enables their characterization and the study of post-transcriptional processes, particularly RNA editing. RNA editing is a post-transcriptional process that involves changes in single nucleotides in RNA sequences. This process is observable in the chloroplast genome, where it affects protein expression and the stabilization of secondary RNA structures (Huang et al., 2022). Among Pellia sensu lato species, RNA editing sites have been localized in the chloroplast genome of Apopellia endiviifolia and the mitochondrial genome of Pellia epiphylla (Grosche et al., 2012; Myszczyński et al., 2019).
In the chloroplast genome of Pellia endiviifolia, 54 RNA editing sites have been identified, all of which exclusively involve C-to-U transitions (Grosche et al., 2012). These modifications lead to nonsynonymous codon changes that have functional significance. The exchange of amino acids in codons can restore or improve the functionality of encoded proteins. RNA editing is particularly important in cases which RNA editing restores the correct start codon (methionine), thereby enabling the translation of a functional protein (Grosche et al., 2012). However, not all chloroplast genes undergo RNA editing; for example, the ndhB gene, which encodes a subunit of the NADH dehydrogenase complex. This variation suggests that RNA editing patterns may differ between individual genes and in Pellia lineages. Unlike M. polymorpha, Pellia retained RNA editing capabilities, supporting the hypothesis that RNA editing was present in all early land plants and was subsequently lost in specific lineages (Grosche et al., 2012).
Analysis of mitochondrial genomes revealed that Pellia epiphylla is also a liverwort species rich in RNA editing sites. Comparative studies have demonstrated an evolutionary trend concerning the decreasing number of predicted editing sites along with diversification observed in the leafy liverworts group (Myszczyński et al., 2018; Ślipiko et al., 2017). This phenomenon may be related to structural changes in mitochondrial genes, such as the loss of introns. This trend is illustrated by comparing the early divergent Haplomitrium hookeri, in which 732 editing sites were predicted, with the simple thalloid P. epiphylla, in which 325 editing sites were predicted (Myszczyński et al., 2019).
Detailed analyses of plastidial genomes have revealed the occurrence of heteroplasmy in the mitochondrial genome of Apopellia endiviifolia, which has significant implications for understanding liverwort evolution (Sawicki et al., 2023). Intra-individual variability caused by this phenomenon may constitute a hidden source of material for natural selection, potentially increasing the adaptive plasticity of liverworts in response to variable environmental conditions. In the A. endiviifolia mitogenome, heteroplasmy manifested as a significant number of indels, up to 475 variants including those in coding regions, of which 30 potentially caused frameshift mutations (Sawicki et al., 2023). Notably, heteroplasmy has not been observed in the chloroplast genome of A. endiviifolia (Sawicki et al., 2023).
Recent advancements in long-read sequencing technologies have facilitated the expansion of genomic research on Pellia sensu lato at the nuclear level. A complete telomere-to-telomere (T2T) genome assembly was recently achieved for the haploid liverwort Apopellia endiviifolia, providing the first chromosome-scale nuclear genome reference for this lineage (Szablińska-Piernik et al., 2025). The assembled genome was 2,914,960,273 bp in size and organized into nine chromosomes, for which telomeric and centromeric regions were identified in silico. Genome annotation, supported by RNA-seq data, led to the identification of 34,615 genes, the majority of which were protein-coding. The analysis revealed substantial variation in gene density among the chromosomes, ranging from 5.52 to 9.78 genes per 500 kbp. The validated centromeres exhibited length variations ranging from 1.9 to 5 Mbp (Szablińska-Piernik et al., 2025). The availability of a complete nuclear genome complements earlier studies on the plastid and mitochondrial genomes of Pellia sensu lato, providing a broader framework for interpreting the evolutionary dynamics of this genus. Together, the complete organellar sequences and T2T nuclear assembly of A. endiviifolia contribute to a comprehensive understanding of genome organization in the early diverging lineages of land plants.
. Ecological significanc
Pellia species play an important role in various ecosystems. They are commonly found in damp, sheltered habitats such as streambanks, rivers, and ditches (Atherton et al., 2010; Schuster, 1992; Schütz et al., 2016). They also occur in wet woodlands, marshes, and wet rocks. Pellia species are particularly abundant in areas with acidic or neutral substrates and are often found growing in dense mats that cover extensive areas. These mats provide habitats for various invertebrates and contribute to nutrient cycling and soil stabilization. Furthermore, Pellia species are sensitive to environmental changes and can be used as indicators of water quality and habitat disturbance (Schuster, 1981).
Pellia endiviifolia, in particular, exhibits a preference for base-rich sites, growing near watercourses, springs, flushes, shaded tracks, woodland rides, fens, dune slacks, moist floors of chalk quarries, and dripping rock outcrops (Schuster, 1981; Schütz et al., 2016). This species’ specific habitat preference further highlights the ecological diversity within the genus Pellia.
. Future research directions
Despite the significant progress made in understanding the genus Pellia, several areas require further research. These include:
Species delimitation and taxonomy: The exact number of species within Pellia remains uncertain, and further research is needed to clarify the boundaries between closely related species and to resolve cryptic species complexes.
Phylogeny and biogeography: Although molecular studies have provided valuable insights into the phylogenetic relationships within Pellia, further research is required to fully resolve the evolutionary history of the genus and understand the factors that have shaped its current distribution.
Ecological interactions: The ecological roles of Pellia species are not fully understood, and further research is needed to investigate their interactions with other organisms and their responses to environmental change.
Physiological adaptations: Pellia species exhibit a range of physiological adaptations that enable them to thrive in diverse habitats. Further research is needed to understand the mechanisms underlying these adaptations and their roles in the ecological success of this genus.
. Conclusion
The genus Pellia represents a fascinating group of liverworts with a complex taxonomic history, diverse morphology, and important ecological role. Taxonomic changes, cryptic speciation, phylogenetic studies, and molecular research have significantly contributed to our understanding of the genus. Although significant progress has been made, several questions remain unanswered. Future research integrating molecular, morphological, and ecological approaches will be crucial to further unravel the mysteries of Pellia and gain a deeper appreciation of its contribution to the biodiversity of our planet.
. Supplementary material
The following supplementary material is available for this article:
Table S1. Comparative morphological and reproductive characteristics of Pellia and Apopellia species. The table presents key diagnostic features, including sexual systems, chromosome numbers, cellular structures, reproductive organs, and spore characteristics. Data compiled from taxonomic and morphological studies (1966, 1981; Schütz et al., 2016). Unknown values indicate insufficient data in the literature.
