Introduction
This paper is a continuation of a previous study (Nobis et al., 2023), and is dedicated to new data on the distribution and taxonomy of vascular plants and bryophytes, or simply, to plants new to the flora of selected countries (or their significant regions) in the Old World (Europe, Asia, and Africa; Sawicki et al., 2023). Here, we present data on 35 species that have been newly reported as components of the flora of 18 countries or their significant regions (provinces or republics). Vascular plant species are represented by 34 taxa that were reported from five European countries, seven Asian countries, and six African countries; additionally, one moss species was obtained from Central Asia (Uzbekistan). The list of taxa is given in alphabetic order; however, it begins with data on vascular plants, followed by data on bryophyte species. Unless otherwise stated, the nomenclature of species is provided following the POWO (2024) checklist, with minor corrections regarding problematic taxa.
VASCULAR PLANTS
Agrostis lazica Balansa, nom. cons. (Poaceae)
Contributor: Beata Paszko
New records
IraN [East Azarbaijan]: Mount Sabalan [Savalan in Azerbaijani], green alpine pasture. Water present in the soil, steep slope, close grazing dominated by Onobrychis and Crepis, 38°10'N, 47°40'E, alt. ca. 3352 m., July 23, 1961, G.N. Harrington 120, Kuh-i-Sabalan Expedition 1961 (K000482656!). Without precise locality: Perse [now Iran], s.d. [1782–1785], [A.] Michaux s.n. (P02224275!).
Taxonomic notes
The genus Agrostis in Iran is represented by 16 species, including Agrostis lazica, which has not yet been reported within the borders of the Iranian flora (Bor, 1970). An ongoing revision of the Agrostis olympica (Boiss.) Bor complex and its close relatives in the Himalayan region, Near East, and Central Asia (Paszko & Pendry, 2013; Paszko, 2014; Paszko in Nobis et al., 2016) revealed the first two records of A. lazica from Iran (Harrington 120, Michaux s.n.). The first record, Harrington 120, was collected during the Kuh-i-Sabalan Expedition in 1961 at Mount Sabalan [Savalan in Azerbaijani] on the Alborz Mountains. Previously, this specimen was misidentified by Bor as A. olympica, and as such cited it in Flora Iranica (Bor, 1970). The above record from Iran considerably expands the known distribution of A. lazica southeastwards. The second record, Michaux s.n. with a label note “Persen [now Iran]” and without a precise locality, was collected during a botanical mission to Persia undertaken by André Michaux (1746–1802) in the years 1782–1785 (Pluchet, 2014). Michaux’s specimen is the oldest collection of A. lazica, and it has not been critically studied since Michaux’s time.
Agrostis lazica was collected and described by Balansa (1874) based on a heterogeneous collection from the Rize district in northeastern Turkey. The name Agrostis lazica has recently been proposed for conservation with a conserved type (Shenzen ICN Art. 14.9) (Paszko & Wolski, 2023). Conservation, as proposed, was recommended (Applequist, 2024).
Agrostis lazica is often confused with other species, particularly A. balansae (Boiss.) Tzvelev (Doğan, 1985). This can also be confused with A. olympica, as was done by Bor. Agrostis lazica differs from A. olympica by longer callus hairs and anthers, shorter paleas, usually glabrous lemmas (hairy all over in A. olympica), and absence of mucros at the tips of lemmas (2–4 mucros up to 0.5 mm long in A. olympica) (Doğan, 1985).
Distribution and habitat
To date, A. lazica has been recorded in Armenia, Azerbaijan, Georgia, Iraq, Russia (Dagestan), and Turkey (Bitlis, Kayseri, Rize) (Rzazade, 1950; Tzvelev, 1976, 2006; Doğan, 1985; Nersesian, 2004; Kurchenko, 2010; Nakhutsrishvili, 2013; Paszko in Nobis et al., 2016). G.N. Harrington 120 is the first record of A. lazica in the East Azerbaijan region of northwestern Iran. Agrostis lazica grows in alpine meadows at about 2500–3300 m (Tzvelev, 1976; Doğan, 1985; Nakhutsrishvili, 2013), and is a characteristic element of alpine grasslands in the lower alpine belt, from 2400 to 2750 m (Zazanashvili et al., 2000). In Iran, it was collected at about 3350 m in an alpine pasture on a steep slope, with a share of Onobrychis and Crepis.
Agrostis × torgesii Portal (Poaceae)
Contributor: Beata Paszko
New record
Spain [Catalonia]: Pyrenees, Núria à Noufonts [Núria at the foot of the Pic de Noufonts Mountain]; alt. ca. 1990 m., September 4, 1913, F. Sennen s.n. (P03659161!).
Taxonomic notes
Agrostis × torgesii is of hybrid origin with A. rupestris All. s.s. and A. schraderiana Bech. and it shows intermediate phenotypic traits relative to those of their parents. The callus hair length to lemma length ratio of A. × torgesii is intermediate (0.2–0.5) compared to that of A. rupestris s.s. (0.05–0.15) and A. schraderiana (0.45–0.90).
Distribution and habitat
The genus Agrostis in the Iberian Peninsula is represented by 16 species, including A. rupestris All. s.s. and A. schraderiana Bech. (Romero-García et al., 1988). An ongoing revision of Agrostis revealed that the first record of A. × torgesii Portal from Spain (Sennen s.n.) was collected by Sennen in 1913 at Núria, at the foot of Pic de Noufonts mountain in the Catalan Pyrenees (Spain). Agrostis × torgesii was described by Portal (2009) in the Haute-Savoie Department in southeastern France. Portal also recorded it from other French departments: Hautes-Alpes, Alpes-Maritimes, Hautes-Pyrénées, and Savoie.
Agrostis × torgesii occurs in alpine and subalpine habitats, together with the parental species A. rupestris All. s.s., and A. schraderiana Bech., which are sometimes sympatric in the Alps and Pyrenees of Europe. If they occur in the same habitat, hybrid swarms are not rare (Portal, 2009).
Crocus heuffelianus Herb. (Iridaceae)
Contributors: Robert Zelek, Marian Szewczyk
New record
POLAND: Western Carpathian Mts, Beskid Niski Mts, Jaśliska Landscape Park, Wisłok Wielki, meadow, alt. 541 m, exp. N, FG 34 (ATPOL grid), March 9, 2024, R. Zelek s.n. (KRA).
Taxonomic notes
The genus Crocus includes over 235 species worldwide (Rukšāns, 2017). Mathew (1982) divided them into two sections: Crocus Mathew and Nudiscapus Mathew. One of the essential groups is the Verni series to which C. heuffelianus belongs. It mainly includes spring-flowering species from Central and Southern Europe, some of which are ornamental plants (Raca et al., 2023).
Crocus heuffelianus is a perennial plant with slightly flattened corms that are covered with fibrous remains of leaf sheaths. Leaves 2–4, narrowly lanceolate, up to 20 cm long and 9 mm wide, glabrous. The base of the stem is covered with a membrane-like sheath that does not cover the leaves. A flower with one bract and a perianth tube up to 15 cm in length. Corolla tepals purple, lilac, rarely white, ovoid, elongated, blunt, or slightly dissected, often with darker spots near the top, 3.5–4.5 cm long, 1.2–1.5 cm wide. The throat is naked. The stigmas are orange and divided into short jagged branches. The seeds are shortly pubescent. It blooms in March and April (Raciborski & Szafer, 1919; Szafer et al., 1986; Mosolygó-Lukács et al., 2016).
Crocus heuffelianus differs from C. scepusiensis (Rehm. & Woł.) Borb., a West Carpathian subendemic that also occurs in Poland, by having a glabrous instead of a hairy throat of the perianth tube (Figure 1).
Figure 1
Crocus heuffelianus in Wisłok Wielki (A); reproductive floral parts and throat of the perianth tube: Crocus heuffelianus (B); Crocus scepusiensis (C) (photo. R. Zelek, 2024).

Distribution and habitat
Crocus heuffelianus is distributed on the lower mountain elevations east of Austria, throughout the Balkan Peninsula, with the southern border in Albania (Gjeta & Hallac, 2018). It extends east to Romania, Ukraine, and northeast Italy (Peruzzi, 2016; Mosolygó-Lukács et al., 2016; Raycheva et al., 2023). The northern limit of its distribution is on the southern border of Poland (Brighton, 1976; Mathew, 1982; Mihaly & Kricfalusy, 1997; Štěpánková, 2011).
Recently, Crocus heuffelianus was found in the Beskid Niski Mts, Wisłok Wielki village, Jaśliska Landscape Park (Figure 1), which is the first locality of this taxon recorded in Poland. Approximately 380 flowering individuals were found in an area of 1200 square meters (the population is about 300 flowering specimens in a meadow (Phytosociological relevé 1 and 2, see below) with an area of 800 square meters, about 70 individuals on an area of about 250 square meters in a meadow with artificial plantings of Larix decidua and Abies alba, and about 10 blooming specimens on an area of 150 m2 in a pine forest with Betula pendula, and the farthest was 15 m from the forest edge during fieldwork in 2024. The population is limited from the east by a drainage ditch. A beneficial measure to maintain the entire population would be to cut down fast-growing trees (Salix caprea, Betula pendula) along the forest edge.
Phytosociological relevé 1. The middle part of the meadow, May 30, 2024 (* March 9, 2024), 25 sq. m, alt. 541 m, exp. N, incl. 5°, herbaceous plant coverage in layer C – 95%, mosses coverage in layer D – 15%; number of herbaceous plants – 52: Crocus heuffelianus 1*, Anthoxanthum odoratum 2, Avenula pubescens 2, Holcus lanatus 1, Trisetum flavescens +, Dactylis glomerata 2, Vicia sepium +, Ranunculus polyanthemos 2, Plantago lanceolata 1, Hypericum maculatum 2, Luzula multiflora 1, Potentilla erecta 2, Centaurea jacea 1, Lychnis flos-cuculi 1, Campanula patula +, Ranunculus acris +, Ranunculus auricomus 1, Veronica chamaedrys 1, Cirsium palustre +, Fragaria vesca +, Briza media +, Tragopogon orientalis +, Alchemilla sp. +, Knautia arvensis +, Prunus spinosa +, Ajuga reptans +, Rhinanthus minor +, Carex pallescens +, Cruciata glabra 1, Deschampsia caespitosa +, Trifolium medium +, Stellaria graminea +, Cirsium rivulare +, Polygala oxyptera +, Equisetum palustre +, Pilosella officinarum 1, Pimpinella major +, Pinus sylvestris r, Hieracium lachenalii +, Rumex acetosa +, Achillea millefolium +, Carex pilulifera +, Veronica officinalis +, Peucedanum palustre +, Festuca rubra 1, Pimpinella saxifraga +, Angelica sylvestris +, Cerastium holosteoides +, Platanthera bifolia r, Heracleum sphondylium +, Polygala vulgaris +, Poa pratensis +.
Phytosociological relevé 2. The middle part of a meadow with artificial plantings, May 30, 2024 (* March 9, 2024), 25 sq. m, alt. 540 m, exp. N, incl. 5°, shrubs coverage in layer B – 15%, herbaceous plant coverage in layer C – 100%; number of shrub species – 4: Abies alba 1, Larix decidua 2, Salix purpurea 1, Salix caprea 2; number of herbaceous plants – 35: Crocus heuffelianus +*, Deschampsia caespitosa 2, Arrhenatherum elatius 1, Holcus lanatus +, Mentha longifolia +, Myosotis palustris +, Hypericum maculatum 1, Agrimonia eupatoria 1, Scirpus sylvaticus 2, Galium verum +, Cirsium arvense +, Poa pratensis +, Chaerophyllum aromaticum 2, Cirsium rivulare 1, Galium mollugo +, Filipendula ulmaria 2, Abies alba +, Ranunculus acris +, Carex hirta +, Pimpinella major 1, Veronica chamaedrys +, Urtica dioica +, Rosa canina +, Poa trivialis 1, Dactylis glomerata +, Lathyrus pratensis +, Potentilla erecta +, Geranium pratense +, Trifolium medium +, Vicia sepium +, Equisetum palustre +, Rumex acetosa +, Prunus spinosa +, Alopecurus pratensis 1, Trifolium pratense + (nomenclature of species is given after Mirek et al., 2020).
Dysphania pumilio (R. Br.) Mosyakin & Clemants (Amaranthaceae)
Synonyms: Ambrina pumilio Moq., Blitum glandulosum Moq., Blitum pumilio C.A.Mey., Chenopodium carinatum f. rubellum Aellen, Chenopodium glandulosum (Moq.) F.Muell., Chenopodium pumilio R.Br., Chenopodium pumilio f. glandulosum (Moq.) Aellen, Chenopodium pumilio f. rubellum (Aellen) Aellen, Neobotrydium pumilio (R.Br.) M.L.Zhang & G.L.Chu, Teloxys pumilio (R.Br.) W.A.Weber.
Contributors: Julian Chmiel, Wojciech Rakowski, Stanisław Rosadziński
New records
POLAND: Wielkopolska region – Poznań-Górczyn, 7 Daleka Street (plot no. 21) – on a 100-metre-long section of pavement along an apartment building, between pavement blocks on driveways, at flower bed edges, and balcony boxes; regularly observed from 2018 to 2023, with a dramatic decline in 2019, elev. 83.5 m, ATPOL square BD0856, 52°23'0"N, 16°53'16"E, May 22, 2018, W. Rakowski, St. Rosadziński s.n. (POZ); Wola Wapowska village (plot no. 64/2) near Kruszwica, Inowrocław County, roadside flower bed along property no. 3, elev. 90.5 m, ATPOL square CD7911, 52°41'49"N, 18°26'26"E, June 16, 2023, J. Chmiel s.n. (POZ-V-0168977); Poznań-Spławie, plant nursery Dobrepole, ATPOL square BD0995, St. Rosadziński s.n. (POZ).
Taxonomic notes
Dysphania R. Br. comprises species characterized by glandules and aromatic secondary metabolites, which are absent in other species assigned to the earlier broadly defined genus Chenopodium (Mosyakin & Clemants, 2002). According to Plants of the World, the new genus comprises 44 taxa. Most originate from Australia and the tropical zones of the Americas, eastern Asia, and Africa. In circumpolar zones, these species are generally sporadic as introduced species. To date, nine species have been recorded in Europe (DAISIE, 2008), mainly in the southern part of the continent. At least four species of this genus occur in Poland: D. ambrosioides (syn. Chenopodium ambrosioides), D. botrys (syn. Ch. botrys), D. pumilio (syn. Ch. pumilio), and D. schraderiana (syn. Ch. schraderanum).
Distribution and habitat
Dysphania pumilio originates from Australia and Tasmania, from where it was unintentionally shipped with wool to New Zealand, Africa, North and South America, and Europe (Dostálek et al., 1990; Akeroyd, 1993; Uotila & Tan, 1997). In the late 19th century, it was accidentally introduced into Central Europe (Ascherson, 1895), but its record was misidentified as a similar species, Chenopodium carinatum (Chytrý, 1993). The presence of this new species has been confirmed e.g. in Italy (Iamonico, 2011), Bulgaria and Greece (Grozeva, 2007), Romania (Chytrý, 1993; Sîrbu et al., 2011), Czechia and Slovakia (Dostálek et al., 1990; Lhotska & Huny, 1979), Norway (Gederaas et al., 2012), Switzerland (Frey et al., 2015), Serbia (Bogosavljević & Zlatković, 2017), Belarus (Dzhus, 2011), and Hungary (Lengyel, 2013). Within its natural distribution range, it can grow on sandy and clayey soils, and sometimes even in saline soils. It can colonize riversides, the edges of water bodies, ruderal sites, and even cultivated fields (Aellen, 1960).
In Central Europe, this species was primarily observed in the plant communities of the alliances Sisymbrion officinalis, Chenopodion rubri, Malvion neglectae, and Polygonion avicularis (Dostálek et al., 1990; Oberdorfer, 1990).
In Poland, D. pumilio was first found in Proszowice near Rybnik, in 1957 (KRAM00028321-V), but misidentified as Chenopodium carinatum. The occurrence of D. carinata in Poland is doubtful based on this single, negatively verified piece of information (Mirek et al., 2020). In the following years, D. pumilio was recorded at two sites: 1974 in port areas in Gdańsk (Misiewicz, 1976; Misiewicz & Korczyński, 2003) and 2005 in Stryków near Łódź (Witosławski, 2009).
Floristic research in the Wielkopolska region from 2018 to 2024 has resulted in the discovery of three new localities of D. pumilio. In the first locality at Daleka Street in Poznań-Górczyn (Figure 2), the population is very large and stable, except for 2019, when its dramatic decline was observed because of the weeding of the green areas. This confirms that the D. pumilio population can regenerate quickly from the soil seed bank. The species composition of the vegetation patches with D. pumilio in Daleka Street includes Cardamine hirsuta, Eragrostis minor, Erigeron canadensis, Poa annua, Portulaca oleracea, and Sagina procumbens, but also many species of bryophytes: Ceratodon purpureus, Barbula convoluta, Tortula muralis, Funaria hygrometrica, Amblystegium serpens, Bryum argenteum, and Marchantia polymorpha. The second population was located in the village of Wola Wapowska near Kruszwica (Inowrocław County), on an unsurfaced road, partly covered with crushed stone, and partly with organic substrate resembling fertilized garden soil for flower beds. The population, composed in 2023 and 2024 of at least 500 individuals, was accompanied by Capsella bursa-pastoris, Chenopodium album, Echinochloa crus-galli, Eragrostis minor, Galinsoga parviflora, Lepidium ruderale, Poa annua, Portulaca oleracea, and Urtica urens. The third locality was found in 2024 in Poznań-Spławie, in the plant nursery Dobrepole, ATPOL square BD0995.
Figure 2
Fragment of a site of Dysphania pumilio (crevices between pavement blocks) on Daleka Street in Poznań (photo. S. Rosadziński, 2022).

According to Mirek et al. (2020), D. pumilio was classified as an ephemerophyte in Poland. However, Misiewicz & Korczyński (2003) suggested that the species can be regarded as naturalized because of its continuous occurrence in Gdańsk. This suggestion was strengthened by Witosławski (2009), who indicated that a future expansion of this species is possible in Poland – very much like in neighboring countries (especially in Czechia), where D. pumilio is classified as naturalized, locally showing signs of expansion. This species has probably been introduced accidentally to Wielkopolska in commercial topsoil. Considering the short observation period, the status of D. pumilio in the Wielkopolska region requires clarification in subsequent years.
Eragrostis virescens J. Presl (Poaceae)
Synonym: E. mexicana subsp. virescens (J. Presl) S. D. Koch & Sánchez Vega (1985)
Contributors: Ewelina Klichowska, Marcin Nobis, Orzimat Turginov, Komiljon Sh. Tojibaev
New records
UZBEKISTAN, Tashkent: near the Institute of Botany, Uzbekistan Academy of Science, on the side of the road, near the sidewalk, 41.3369172N / 69.3411618E, September 2, 2024, M. Nobis, E. Klichowska s.n. (KRA634316); near the Institute of Botany, Uzbekistan Academy of Science, on the side of the road, near the sidewalk, 41.33811400N / 69.3394000E, September 4, 2024, M. Nobis, E. Klichowska, A. Rudov s.n. (KRA634317); on the side of the road, near the sidewalk, Mirishkor ko’chasi St., 41°19'55.85"N 69°20'28.85"E, September 3, 2024, M. Nobis, E. Klichowska s.n. (KRA634318).
Taxonomic notes
Eragrostis Wolf (lovegrass) is one of the largest genera of the family Poaceae, which comprises over 400 taxa (Clayton et al., 2006). Within this genus, there are many groups of morphologically similar species. One of them is the E. pilosa-pectinacea complex, encompassing species from different continents and represented in Eurasia by seven wild occurring (E. pectinacea, E. pilosa, E. multicaulis, E. albensis, E. amurensis, E. imberbis, and E. virescens) taxa, as well as two crops E. tef, and E. barrelieri, E. pectinacea, E. pilosa, E. multicaulis, E. albensis, E. amurensis, E. imberbis., and E. virescens. According to the identification key provided by Martini & Scholz (1998), and the morphological description presented by Wróbel et al. (2021), the last mentioned E. virescens, differs from closely related species E. pilosa, E. imberbis, and E. tef, by having panicles with solitary (vs verticillate) inferior branches as well as subequal glumes (vs. unequal glumes). It can be distinguished from E. pectinacea by its longer culms (50–70 cm vs. 10–25 cm) and wider spikelets (1.5 mm vs. 1 mm). Whereas from E. multicaulis, differs by having tufts of long hairs at the throat of the leaf sheaths, which is lacking in E. multicaulis. In contrast to E. barrelieri, E. virescens has no ring of glandular tissue below the nodes on the culms. This species differs from E. amurensis in the absence of glands on leaf sheaths and blades, which are present in E. amurensis. Whereas from E. albensis it can be distinguished by having glabrous axils (or nearly so) vs. panicle axils with hairs.
Distribution and habitat
Lovegrasses are distributed from tropical to temperate regions of all continents, except Antarctica. They are mainly associated with dry, sandy areas or human-disturbed habitats. Some species of economic importance, such as E. tef, E. barrelieri, and E. curvula, have been intentionally introduced into many regions worldwide (Lee et al., 2013; Stoyanov, 2014; Cheng et al., 2017). While the first two species spread mainly in anthropogenic habitats (Weakley & Southeastern Flora Team, 2024), the second one is much more expansive, invading the gravelly floodplains of river valleys (Muranaka & Washitani, 2004). Other Eragrostis species accidentally introduced to areas beyond their natural range also show a tendency to be invasive, such as the Central Asian E. albensis, which is spreading rapidly in Europe (Wróbel et al., 2021). Another species, the South American E. virescens, has been introduced to North and Central America (Koch & Sánchez Vega, 1985), S Africa (Smook, 1990), Australia (Lazarides, 1997), many European countries (Tutin, 1980; Ryves et al., 1996; Lambinon, 1997; Martini & Scholz, 1998), and recently to Central Asia (Tajikistan; Nobis & Nowak, 2011), where it grows in human-disturbed habitats such as roadsides and sidewalks between pavement slabs, or wastelands.
In September 2024, a new locality for E. virescens was found in Tashkent (Uzbekistan), where the species had not been previously reported. In the capital city of Uzbekistan, the species spread along the sidewalks, from where it was collected (Figure 3).

Erigeron canadensis L. (Asteraceae)
Synonyms: Aster canadensis (L). E.H.L.Krause, Conyzella canadensis (L.) Rupr., Conyza canadensis (L.) Cronquist, Leptilon canadensis (L.) Britton
Contributors: Shukherdorj Baasanmunkh, Batlai Oyuntsetseg, Hyeok Jae Choi
New record
MONGOLIA: Selenge province, Yuruu soum, Bugant tosgon, Yuruu river; herbs, N 49°25'47.6" / E 107°22'37.4", elev. 1836 m, July 9, 2024, B. Oyuntsetseg, H. J. Choi, S. Baasanmunkh NW24_09 (UBU0036360-UBU0036364).
Taxonomic notes
Erigeron canadensis is an annual species that belongs to the Asteraceae family. Until recently, the species was classified as Conyza canadensis (L.) However, molecular studies have allowed its retransfer to the genus Erigeron (Noyes, 2000; Oh & Park, 2023). In general, based on morphological features, E. canadensis is similar to E. bonariensis L. and E. sumatrensis Retz. (Chen & Brouillet, 2011; Nobis et al., 2018) but can be easily distinguished by the bract of the involucre (Figure 4C) (glabrous vs. hairy or densely hairy), stems (glabrous vs. very hairy), and leaf margin (Figure 4F) (entire vs. coarsely serrate).
Figure 4
Erigeron canadensis in Mongolia: (A) general habit; (B) inflorescences; (C) involucre bracts; (D) receptacle and phyllaries; (E) seed; (F) leaves (photo. Bugant tosgon, Selenge province, by S. Baasanmunkh and D. Munkhtulga, 2024).

Distribution and habitat
The native range of E. canadensis includes North America, and its non-native range spans the rest of the Northern Hemisphere (Shah et al., 2014; Lucas et al., 2024). However, this species has not yet been recorded in Mongolia (Baasanmunkh et al., 2022). During our field expeditions, we found E. canadensis in the northern part of the country, which is a new record for Mongolian flora. In general, E. canadensis grows as an invasive species in open areas, such as roadsides, margins of farmland, and forests, causing ecological issues for native plants (Yan et al., 2020). In Mongolia, this species occurs in the mining areas of the Bugant tosgon, Selenge Province ( Figure S1). The population of E. canadensis in this area is less than 500 individuals.
Gagea tenera Pascher (Liliaceae)
Synonyms: Gagea kashmirensis Turrill; Gagea persica var. kashmirensis (Turrill) S.Dasgupta & Deb
Contributors: Serik Kubentayev, Daniyar Alibekov
New record
KAZAKHSTAN: Ulytau Region, Ulytau District, near the Kinembay reservoir, along the bank of the river, 48°39'37.6"N / 66°34'31.1"E, April 30, 2024, D.T. Alibekov et al. s.n. (NUR). Karaganda region, Aktogay district, Bektauata mountain, near Karabuta, rubbly slopes of foothills of granitoid hills, 47°22'20.3"N / 74°50'40.1"E, April 20, 2023, S.A. Kubentayev et al. 7085 (NUR).
Taxonomic notes
Gagea tenera belongs to the large section of Didymobolbos (K. Koch) Boissier, which includes more than 30 species (Peterson et al., 2008; Tison et al., 2013), is distributed mainly in Europe and along the Mediterranean coast of Africa. The main characteristic of G. tenera, which distinguishes it from other species of the Didymobolbos section, is the presence of a leaf bundle in a head-like cluster of vegetative bulbs on an underdeveloped peduncle. G. tenera in the new location in Central Kazakhstan is characterized by small size (3.5–8 cm), subtilty and few flowers (1–2, rarely 3 flowers). This can be explained by the more severe habitat conditions in Central Kazakhstan. Further molecular taxonomic studies of the Central Kazakhstan populations of G. tenera are needed. It is possible that G. tenera in Central Kazakhstan forms a separate isolated race, which can be classified as a subspecies or variation.
Distribution and habitat
The genus Gagea comprises more than 370 species that are morphologically, genetically, and arealogically delimited within its Eurasian range (Levichev, 1999, 2013). A significant part of the representatives of the genus (164 species) is concentrated in two modern centers of its diversity: the Western Tien Shan – 77 (21%) species and Pamir-Alai – 122 (33%) species. More than 60 species of Gagea grow in Kazakhstan (Kubentayev et al., 2021), of which four species are national endemics (Kubentayev et al., 2024). G. tenera grows in mountains and foothills on stony and rubbly slopes, as well as along rock crevices. The center of the species range includes the Western Tien Shan and Pamir-Alai; however, some marginal populations are also found in the Hindu Kush and Himalayas and westward to Elbrus and Transcaucasia up to Anatolia. In Kazakhstan, the species was previously known from the southern regions of the country of Northern and Western Tien Shan, including Dzungarian Alatau, Zailiyskiy Alatau, Karatau, and the Chu-Ili Mountains (Goloskokov, 1958). During field surveys in late April 2023, G. tenera was discovered for the first time in Central Kazakhstan in the Bektauata Mountains (Figure 5), and a year later, the species was found in the Ulytau Mountains. In Bektauata, the species grows on the fine-grained slope of the foothill terrace and granitoid uplands near a small stream. In the Ulytau Mountains, the species grows on the slope of a shallow gorge formed by the Usabaybulak River in fine-gravelly soil. The gorge is located below the dam of a small, unnamed reservoir. The species occurs in the community with Gagea bulbifera, G. fedtschenkoana, Alyssum sp., Tulipa patens, Elymus repens, Spiraea hypericifolia, Valeriana tuberosa. New finds of G. tenera are located at a considerable distance to the north from the known localities of this species in Kazakhstan. The closest known locality of G. tenera is in the Karatau Mountains (Kazakhstan) (Ebel et al., 2017) at a distance of more than 500 km from the new location in Ulytau and 700 km from the location in Bektauata. This species has not previously been reported in the Ulytau Region, Karaganda region, Kazakh Upland, and in general, in Central Kazakhstan (Goloskokov, 1958; Karamysheva & Rachkovskaya, 1973; Kupriyanov, 2020). New locations of G. tenera in Central Kazakhstan are the northernmost locality of G. tenera and provide new insights into the distribution and ecology of this species. The data from the new localities allowed us to significantly expand the range of this species to the north.

Grindelia squarrosa (Pursh) Dunal (Asteraceae)
Synonyms: Aurelia amplexicaulis Cass.; Aurelia squarrosa (Dunal) Cass. ex Steud.; Donia squarrosa Pursh; Grindelia angustifolia Kunth; Grindelia arguta A.Gray; Grindelia arguta Schrad. ex DC.; Grindelia perennis f. pseudopinnatifida D.Löve & J.-P.Bernard; Grindelia pinnatifida Wooton & Standl.; Grindelia serrulata Rydb.; Grindelia speciosa Gillies ex Hook. & Arn.
Contributors: Daniyar Alibekov, Serik Kubentayev
New records
KAZAKHSTAN: Akmola Region: Zharkain District, neighborhood of Derzhavinsk city, on the edge of willow thickets, in a low relief, 51°04'56.8"N / 66°17'31.1"E, July 22, 2021, S.A. Kubentayev, D.T. Alibekov s.n. (NUR); Astrakhan District, near Zhaltyr, 51°38'30.6"N / 69°49'25.1"E, July 30, 2024, D.T. Alibekov, Zh. T. Idrisova s.n. (NUR); Zharkain District, neighborhood of the town of Derzhavinsk, near a wheat field and a rubbish dump, 51°04'40.8"N / 66°17'14.6"E, July 30, 2024, D.T. Alibekov, Zh. T. Idrisova, K. S. Izbastina s.n. (NUR). Abay Region, Tarbagatai-Ayagoz Road, at the entrance to Ayagoz Town, bank of Ayagoz River, 47°56'50.60"N / 80°25'02.30"E, June 28, 2016, D.T. Alibekov s.n. (KG).
Taxonomic notes
Grindelia squarrosa is morphologically close to Grindelia hirsutula Hook. & Arn., which is often confused with. For example, in Germany, these two species have not been recognized for a long time, and it is considered that only G. squarrosa grows in this country. Although the identification key compiled by Jäger E. J. provides clear distinguishing features of these two species, this information has long been ignored (Jäger, 2008; Fischer et al., 2023). First, G. squarrosa is a biennial, approximately 40 cm high plant, that does not form a root rosette. Its leaves are obovate or obovate-lanceolate, flower heads are 2.5–3.5 cm width, and there are 12–40 ray flowers. Whereas, G. hirsutula is a perennial plant with a root rosette, 60–80 cm leaves are large, broad, fleshy with toothed margins; flower heads are 2.5 to 6 cm width, with 15–60 ray flowers (Jäger, 2008). Previously, G. nuda, which is characterized by discoid heads and blunt or rounded leaves, was synonymized with G. squarrosa in the Flora of North America (Strother & Wetter, 1993+). However, the latter is treated as a separate species (Powell & Worthington, 2018).
Distribution and habitat
Of the 44 species in the genus Grindelia (Steyermark, 1934), only G. squarrosa has spread to almost all continents. G. squarrosa is the native species of North America. Its natural range is probably in the Great Plains and Rocky Mountain regions (Strother & Wetter, 1993+). The species apparently naturalized in the eastern United States (Gleason & Cronquist, 1991) and was subsequently the species has been introduced to other parts of North America, South America, Europe, and Asia (Hansen, 1976; Shevera et al., 2019; Pliszko & Górecki, 2021). The species is currently naturalized in six European countries (Bulgaria, Romania, Moldova, Ukraine, Poland, and Russia) (Shevera et al., 2019; Zając & Zając, 2019). We first found G. squarrosa in Kazakhstan in 2016, near the town of Ayagoz (East Kazakhstan region), on the bank of the Ayagoz River, disturbed by grazing near a motorway. During a field trip in 2021, G. squarrosa was found near Derzhavinsk, Akmola region (Figure 6). The population of this species is located in a lowland, open glades of Salix sp. G. squarrosa grows in a ruderal community with Achillea nobilis, Aeluropus littoralis, Agropyron cristatum, Amaranthus albus, Artemisia austriaca, A. schrenkiana, Atriplex sagittata, Convolvulus arvensis, Descurainia sophia, Falcaria vulgaris, Galatella villosa, Lepidium latifolium, Lophiolepis serrulata, Palimbia turgaica and Sonchus arvensis. Despite its origin in the arid regions of North America, the species prefers moderately humid habitats. It is difficult to establish the original distribution center of G. squarrosa in Kazakhstan, but all locations are near railway stations; therefore, the species is spread by railway transport, transport of fodder, food grain, hay, etc. Grindelia squarrosa is a new genus and species for the flora of Kazakhstan.

Lilium buchenavii R.J. Koch (Liliaceae)
Contributor: Zbigniew Szeląg
New records
Poland: Masuria Lake District, N of Gołdapiwo Lake: 1. 700 m N of Jasieniec village, Pinus sylvestris-Acer platanoides forest margin by a road, 54°08'18.5"N 21°58'22.6"E, June 26, 2022; 2. 300 m E of “Aneta” camping site, openings in Pinus sylvestris-Quercus robur forest along a road, 54°07'59.7"N 21°58'03.0"E, June 19, 2022; 3. Jasieńczyk settlement, on over-grown meadow, 54°08'00.4"N 21°57'18.3"E, June 24, 2022; 4. 100 m W of a lock on the Sapina River, grassy slope, 54°08'06.4"N 21°55'35.9"E, July 1, 2022; 5. Przerwanki village, 150 m E of a bridge on the Sapina River, Pinus sylvestris forest margin, 54°08'10.9"N 21°55'35.3"E, July 5, 2022. 6. Masuria Lake District, SW of Gołdapiwo Lake, by a road from Kruklanki to Sałdany villages, Pinus sylvestris-Tilia cordata forest margin, 54°05'10.2"N 21°53'58.1"E, July 2, 2023 (no herbarium specimens and only living plants for future garden cultivation were collected).
Taxonomic notes
In 2022, a lily that looked like Lilium bulbiferum L. was found in the central part of the Masuria Lake District, NE Poland (Figure 7). Observation of these plants during cultivation showed that they differed from L. bulbiferum s.str. that occurs in Poland only in the Sudetes and Western Carpathians (Koczur & Świerkosz, 2014). A recently published paper on lilies of the L. bulbiferum aggregate in Germany (Koch, 2023) identified the plants found in Poland as L. buchenavii. As this beautiful plant was found in the Masuria Lakeland, the Polish name “lilia mazurska” is proposed.

The authorship of the name L. buchenavii requires comment. Koch’s (2023) intention was to validate of the forgotten name L. buchenavii, which was first used by Focke (1883) as provisional name for plants growing in the lowlands of northern Germany, and which were mistakenly treated as L. bulbiferum by Buchenau (1877). However, Koch (2023) described this species anew; therefore, the authorship of the name L. buchenavii must be attributed to him alone.
The morphological differences between L. buchenavii and L. bulbiferum are as follows:
1. Flowers orange-red. Bract glabrous on the lower surface. Axillary bulbils purplish-brown, only within inflorescence ………………………… L. buchenavii
– Flowers orange. Bract hirsute on the lower surface. Axillary bulbils white-green, in axils of stem leaves and within inflorescence ………………………… L. bulbiferum
Distribution and habitat
Lilium buchenavii was found in several places along the northern shore of Gołdapiwo Lake and near Kruklanki village to the south of the lake. The discovery of additional sites in this region is highly likely. It is very likely that the plants reported as L. bulbiferum from the vicinity of the towns of Mrągowo and Szczytno (Abromeit et al., 1940) are also L. buchenavii.
Hitherto, L. buchenavii was known only from Lower Saxony in Germany, where it grows on light soils in the old moraine area, on fallow land, embankments, and occasionally in gardens (Koch, 2023). Its occurrence in northern Poland is restricted to the moraine landscape of Masuria Lake District. Lilium buchenavii prefers sandy grasslands and abandoned meadows, as well as loose thickets and forest edges with a predominance of Pinus sylvestris. In all localities, it grows singly or in small clusters.
Neottia ovata (L.) Hartm. (Orchidaceae)
Synonyms: Listera ovata (L.) R. Br., Helleborine ovata (L.) F.W.Schmidt, Malaxis ovata (L.) Bernh., Epipactis ovata (L.) Crantz, Ophrys ovata L., Neottia latifolia Rich.
Contributors: Arkadiusz Nowak, Sebastian Świerszcz
New record
TAJIKISTAN: Chorbog, N 39.43741, E 67.73215, elev. 1182 m a.s.l., gaps in wooded area, June 5, 2019, A. Nowak s.n. (OPUN); Rarz, N 39.37981, E 68.75534, elev. 1541 m a.s.l., sparse tree stand, June 15, 2019, A. Nowak obs.
Taxonomic notes
Neottia ovata is a short-to-medium rhizomatous plant with an erect stem and hairy above. It has only two oval, dull green, conspicuously ribbed leaves and low at the base of the stem. Flowers are small, numerous in slender, spike-like racemes with tiny bracts, stalked, yellowish-green, 7–15 mm long, hanging, notched at the end, and spurless.
Distribution and habitat
Neottia ovata is mainly distributed in the Euro-Siberian and Mediterranean regions (POWO, 2024). The species is native to much of Europe, including the British Isles, Siberia, Central Asia, Southwest Asia, and Himalayas. It has been introduced in Ontario (Kotilínek et al., 2015; POWO, 2024). In Central Asia, the species has only been found in Kyrgyzstan, Uzbekistan, Kazakhstan and Afghanistan, e.g. from Dzungar Ala-too and Tian Shan (Bondarenko, 1971; H. Freitag, pers. comm.).
During fieldwork in western Tajikistan in June 2019, this species was found in an old apple orchard on the outskirts of the village of Chorbog (Figure 8; Nowak & Nobis, 2020). It grows in grassland vegetation in a tree gap on the slope of the southeastern exposition towards the Mogiendarya River. The size of the population was moderate, with approximately 50 inflorescences found in 2019. In Chorbog, the population of N. ovata was accompanied by: Achillea millefolium, Agrimonia asiatica, Aphanopleura capillifolia, Arenaria serpyllifolia, Avena trichophylla, Bromus lanceolatus, Bromus oxyodon, Crepis pulchra, Dactylis glomerata, Elytrigia trichophora, Hordeum bulbosum, Koelpinia linearis, Malus sieversii, Medicago orbicularis, Medicago sativa, Plantago lanceolata, Scabiosa songorica, Trifolium pratense and Vulpia persica. In Rarz, it grows in tree groves on the southwestern slopes with grassy undergrowth, dominated by Bromus spp. within the village limits. During a field survey in 2019, we observed ca. ten individuals.

Nonea lutea (Desr.) DC. (Boraginaceae)
Synonyms: Anchusa lutea (Desr.) M. Bieb., Echioides lutea (Desr.) Poir., Lycopsis lutea Desr.
Contributors: Artur Pliszko, Krzysztof Kapała, Bogusław Binkiewicz, Mykhaylo Chernetskyy, Marcin Nobis
New records
POLAND: Kraków Bridge, Kraków, Księcia Józefa Street, N50°02.670', E19°51.225' – roadside ditch, roadside verges, edge of thickets in a private garden, exposure: S, elev. 208 m, DF69 (ATPOL grid), April 16, 2023, A. Pliszko s.n. (KRA); Grzegórzecka Street N50°03.504', E19°57.348' – urban green area, elev. 209 m, DF69 (ATPOL grid), April 20, 2023, A. Pliszko obs.; Wawel Royal Castle, N 50°03.180', E19°56.154' – lawn on a slope, elev. 202 m, DF69 (ATPOL grid), May 1, 2023, M. Nobis obs., and April 7, 2024, A. Pliszko s.n. (KRA); Bochnia Foothills, Niepołomice, Plac Zwycięstwa Street, N50°01.962', E20°12.975' – neglected lawn and pavement, elev. 200 m, EF61 (ATPOL grid), April 7, 2024, A. Pliszko s.n. (KRA); Lublin, Nałęczów Plateau, Aleje Racławickie, N51°15.839', E22°30.884', lawns, FE27 (ATPOL grid), April 10 and 16, 2024, M. Chernetskyy obs.; Spadochroniarzy Street, N51°14.992', E 22°31.896' – lawns, FE27 (ATPOL grid), April 09 and 21, 2024, M. Chernetskyy obs.; Czwartaków Street, N51°14.795', E 22°31.855' – grassland slope and ruderal ground, FE27 (ATPOL grid), April 20, 2024, M. Chernetskyy obs.; Mariana Langiewicza Street, N51°14.720', E22°32.085' – edge of the sport field and grassland slope with ornamental plants, FE27 (ATPOL grid), April 14 and 20, 2024, M. Chernetskyy obs.; Głęboka Street, Akademicki Park, N51°14.691', E22°31.914' – park, FE27 (ATPOL grid), April 20, 2024, M. Chernetskyy obs.
Taxonomic notes
The genus Nonea Medik. belongs to the tribe Boragineae in the family Boraginaceae, and includes 46 species of herbaceous plants. It is characterized by an accrescent fruiting calyx, hairy appendages at the throat of the corolla, and glandular hairs on leaves, stems, and/or inflorescences (Selvi et al., 2006; Yeşil, 2017; POWO, 2024). In the flora of Poland, it is represented by one native species, N. pulla (L.) DC., and two alien species, N. lutea (Desr.) DC. and N. rosea (M. Bieb.) Link (Mirek et al., 2020). Nonea lutea is an annual plant that is up to 60 cm tall (Teofilovski & Ćušterevska, 2022). It can be distinguished from N. pulla and N. rosea by its yellow flowers and oblong-cylindrical longitudinally rugose nutlets (Pakravan et al., 2009).
Distribution and habitat
Nonea lutea is native to southeastern Europe, south-western, and central Asia. It was introduced in other parts of Europe and Asia, Australia, and North America (Eberwein, 2011; Randall, 2017; CABI, 2022; Teofilovski & Ćušterevska, 2022). In its native range, it occurs on steppes, farmlands, roadsides, and in oak forests (Eberwein, 2011; Teofilovski & Ćušterevska, 2022). In Western and Central Europe, it is usually found in botanical gardens, arable fields, roadsides, railway embankments, lawns, thickets, hedges, and waste areas (Galera & Sudnik-Wójcikowska, 2004; Eberwein, 2011; Dvořák et al., 2019; Krumbiegel, 2020; Teofilovski & Ćušterevska, 2022). It has also been naturalized in Croatia, Czechia, Germany, Italy, Macedonia, and Switzerland, as well as in some parts of Ukraine and Russia (Pyšek et al., 2012; Cecchi & Selvi, 2015; Randall, 2017; Dvořák et al., 2019; Krumbiegel, 2020; Teofilovski & Ćušterevska, 2022). It is not considered an invasive species in any country in the world (Randall, 2017; CABI, 2022), although it has become a troublesome weed in some botanical gardens (Eberwein, 2011).
In Poland, N. lutea has been known from three localities and its establishment has been overlooked. The first locality was found in Wrocław, south-western Poland, near the Botanical Garden of the Wrocław University (BGWU), in 1877, the second, in Kraków, southern Poland, in the vicinity of the Botanical Garden of the Jagiellonian University (BGJU), in 1950, and the third, in Lublin, south-eastern Poland, in the area and vicinity of the Botanical Garden of the Maria Curie-Skłodowska University (BGMCSU), in the 1990s (Kornaś, 1954; Galera, 2003). Moreover, it was observed in BGJU and its close vicinity in the 1990s (Galera, 2003). Surprisingly, N. lutea has been treated as a casual alien plant (Urbisz, 2011; Mirek et al., 2020), although it has spread spontaneously in BGJU and BGMCSU for many years (Galera, 2003; Guzik, 2006, 2012; Binkiewicz et al., 2017).
The presence of N. lutea was confirmed in BGJU, BGMCSU, and their surrounding areas. In addition, three new localities in Kraków, two new localities in Niepołomice and Zawoja, and five new localities in Lublin were recorded between 2023-2024. In Kraków, the most abundant stands of N. lutea (ca. 10 000 individuals) were observed under the planted trees of Metasequoia glyptostroboides Hu & W.C. Cheng in the north-eastern part of BGJU in 2023 (Figure 9). In addition, large patches were noticed in a ruderal ground and domestic garden along Żółkiewskiego Street, up to 50 m from the southwestern border of BGJU (ca. 1 000 individuals). Moreover, a few smaller patches were observed outside the fence of BGJU, namely on a lawn at the building of the Herbarium of the Institute of Botany of the Jagiellonian University at Kopernika Street (25 individuals), in an urban green area at Kopernika Street near the Mogilskie Roundabout (12 individuals), and on lawns at Kosińskiego Square (ca. 350 individuals). Going further, N. lutea was found in an urban green area at Grzegórzecka Street, approximately 265 m from the borders of BGJU (ca. 300 individuals), on a slope in the area of Wawel Royal Castle, about 2 km from the borders of BGJU (ca. 500 individuals) and roadside verges, in a roadside ditch, and on the edge of thickets in a neglected domestic garden at Księcia Józefa Street, approximately 8 km from the borders of BGJU (ca. 600 individuals) (Figure 9). In Niepołomice, N. lutea was recorded on a lawn and on a pavement (ca. 150 individuals), whereas in Zawoja, it was found on private property near the piles of firewood and building materials (three individuals). In Lublin, the largest population of the plant was observed in BGMCSU (ca. 1000 individuals), and in the remaining sites located within a distance of about 3 km from BGMCSU, the abundance ranged from a few to about 300 individuals. Most likely, the new localities of N. lutea in Kraków and Lublin are related to botanical gardens, which seem to be a source of the plant. However, the origin of this stand in Niepołomice remains unknown. Nevertheless, N. lutea seeds can be easily carried by humans over long distances, that is, on shoes, vehicles, or garden and construction machinery, as well as with garden soil, compost, and seed material (Eberwein, 2011; Randall, 2017; Dvořák et al., 2019; Teofilovski & Ćušterevska, 2022). Moreover, the seeds of the plant can be carried by ants over short distances (Dvořák et al., 2019), which may explain their presence outside flower beds and fences in BGJU and BGMCSU. Interestingly, N. lutea was accidentally introduced to Zawoja along with wood from BGJU, but this stand was ephemeral.
Figure 9
Nonea lutea in Kraków, southern Poland: (A, B) flowering individuals in the Botanical Garden of Jagiellonian University; (C) lawn at Kosińskiego Square near the fence of the Botanical Garden of Jagiellonian University; (D) roadside at Księcia Józefa Street (photo. A. Pliszko, 2023).

Considering the long period (over 20 years) of spontaneous occurrence of N. lutea in Kraków (Galera, 2003; Guzik, 2006, 2012; Binkiewicz et al., 2017) and Lublin (Galera, 2003), it can be classified as a locally established alien species in Poland. It prefers sunny or semi-shaded anthropogenic habitats with ruderal plants, such as Capsella bursa-pastoris, Cardamine hirsuta, Chelidonium majus, Glechoma hederacea, Poa annua, Senecio vulgaris, Stellaria media, and Taraxacum sect. Taraxacum, and Veronica hederifolia. Similar to other European botanical gardens (Eberwein, 2011), in BGJU and BGMCSU, N. lutea exhibits rapid growth, fast fruit ripening, and the deposition of large amounts of seeds in the soil seed bank, which makes it difficult to eradicate. However, it is regularly mowed and weeded from flower beds. Further spread of N. lutea in Poland is likely; therefore, it should be monitored and controlled to prevent its invasion.
Orobanche clausonis Pomel (Orobanchaceae)
Synonyms: Orobanche galii var. atlantica Batt., Orobanche lucorum var. hesperina J.A. Guim., Orobanche clausonis subsp. hesperina (J.A. Guim.) M.J.Y. Foley
Contributors: Óscar Sánchez Pedraja, Renata Piwowarczyk, Thorsten Usée
New records
FRANCE: Pyrénées-Orientales Department, Tour Carroig, Corbère, May 9, 1972, Loius Amandier n. 255 (MPU499695, as Orobanche caryophyllea Smith [sic]); Aude department, Fitou, 42°53'23.4"N 2°57'34.3"E, May 7, 2023, T. Usée (https://www.inaturalist.org/observations/161155907);
ITALY: Sicily, Trapani province, Monte Inici, 650 m, May 2013, Beppe Di Gregorio (https://www.actaplantarum.org/forum/viewtopic.php?f=166&t=57516 [as Orobanche caryophyllacea Sm. „(apocromatica)”, on Galium sp.]);
MOROCCO: Ifrane province, El-Hajeb to Ifrane, 14 km to Ifrane. Forest of Jaba, 33°32'08.0"N 5°14'26.6"W, perhaps loc. calcareous. (protruding rocks), May 23, 1961. Fl. yellowish white. parasite on Galium sp., J.J.F.E. et P.A.W.J. de Wilde et J. Dorgelo no. 2394 (WAG1141150, AMD84012 [as O. alba]);
TUNISIA: Ben Arous Governorate, Dj. Reças [Djebel Ressas, Dorsale Tunisienne], May 22, 1888 [Mission Botanique de Tunisie], G. Barratte (P03428435-hb. E. Cosson [as Orobanche („stigm. purpureo”)]; Figure 10).
Figure 10
Orobanche clausonis, Tunisia, Djebel Ressas (P03428435-hb. E. Cosson) (https://science.mnhn.fr/institution/mnhn/collection/p/item/p03428435).

Taxonomic notes
Currently, Orobanche clausonis is considered a separate taxon by most authors, although this is not always the case. The species was described by Pomel (1874–1875) on Algerian material from Ouled-Mazis y Mouzaïa, the material of the last place from a Clauson gathering, and hence the name of the species. Pomel considered it different from the rest of the genus, even though this species forms a different subgenus (Orobanche subg. Monoschiza). Later, Battandier (1890) described O. galii var. atlantica, considering Pomel’s species to be a form that must be included in the variety he described. Battandier apparently included his variety in O. galii for its similar habit, but smaller in all its parts, and also to be parasitic on species of the Rubiaceae family. Several years later, Guimarães (1904) described on Portuguese materials a new taxon O. lucorum var. hesperina. This new taxon was placed as a variety of O. lucorum, probably based on its yellowish coloration, because in the other characters, it bears little resemblance to the alpine plant, which Guimarães indicates never seen in Portugal. In his monographic study of the Orobanchaceae of the Iberian Peninsula, Foley (1996) considered different African forms of the Iberians; therefore, he retained the Pomel name for the Africans and created a new combination based on that of Guimarães (O. clausonis subsp. hesperina). After studying abundant material (Sánchez Pedraja et al., 2016+), we think that the differential characters indicated by Foley for his subspecies fit within the variability of this species, so we consider Foley’s name as a mere synonym of the Pomel plant.
Distribution and habitat
The species was described in Algeria by Pomel (1874–1875) and has a West Mediterranean distribution, spreading through many territories that surround the west of this sea (Algeria, Balearic Islands, France, Italy (Sicily), Morocco, Portugal, Spain, and Tunisia) (Sánchez Pedraja et al., 2016+). It is relatively abundant in the Iberian Peninsula, where its westernest distribution touches the coast of the Atlantic Ocean (Berlengas Islands) and the NW of Africa that borders the Mediterranean Sea. France and Italy (Sicily) in Europe and Tunisia in Africa now constitute its oriental distribution limit. With the discovery in the herbaria P and MPU and the photographs in the iNaturalist.org (2024) and Acta Plantarum (2024), we expanded the species’ distribution to these three countries, which we are now indicating as the oriental limit to its range. In addition, we provide the only concrete Moroccan appointment (no “calculated”) backed by herbarium material (Rumsey & Jury, 2002: 591; Dobignard & Chatelain, 2013: 20; Fennane, 2018).
Orobanche teucrii subsp. sideritidis (Huter, Porta & Rigo) Ó. Sánchez & Piwow. (Orobanchaceae)
Synonyms: Orobanche sideritidis Porta, nom. nud., Orobanche sideritidis Huter, Porta & Rigo, Orobanche caryophyllacea var. typica subf. thyrsiflora (Wallr.) Beck, p.p., Orobanche teucrii auct. (e.g., Carlón et al., 2008; Govaerts et al., 2021; Hassler, 2024), Orobanche caryophyllacea auct. (e.g., Caruel, 1885: 372; Beck, 1890: 157; Beck, 1930: 232; Uhlich & Rätzel, 2024)
Contributors: Óscar Sánchez-Pedraja, Renata Piwowarczyk
New records
ITALY: Calabria Region, Cosenza Province: Dirupata di Morano [north-west of Morano Calabro] (Caruel, 1885: 372, sub O. caryophyllacea; Porta, 1879: 268, sub O. sideritidis; Beck, 1890: 157, sub O. caryophyllacea var. typica subf. thyrsiflora; Huter 1907: 354, sub O. sideritidis; Carlón et al., 2008: 118, sub O. teucrii; Uhlich & Rätzel, 2024: e04155, sub O. sideritidis); Sila Grande ca. 14 km ENE of Camigliatello Silano, Arnocampo, 1550–1650 m s.m., 39°20'49.0"N 16°36'39.0"E, June 11, 1997, G. Cesca et al. no. 1063 (B101250727, sub O. caryophyllacea); “1–5 Italien-Calabria-Mormanno-Parco del Pollino [Mormanno comune, National Park of Pollino] 16.05.2006” by Ernst Gubler (http://www.alpenbotaniker.ch/Wuerger-Labkraut-Mi.html, sub O. caryophyllacea, apparently on Sideritis italica; accessed March 2024); Italy, Calabria Region, Catanzaro Province: Sila-Gebirge, Comune Tiriolo; etwa 0,7 km E über Tiriolo im südlichen Teil der Verebnungsfläche am Monte Tiriolo; 775 m, 38°56'53.5"N 16°30'58.8"E, August 5, 2014, K. Zernig no. 8827 (GJO0083714, sub O. caryophyllacea); Italy, Campania Region, Salerno Province: “Province of Salerno [40°34'06.3"N 15°29'10.5"E, near Caggiano], April 17, 2019” (https://www.inaturalist.org/observations/192616090 by https://www.inaturalist.org/people/yossarianc, sub O. teucrii; apparently on Sideritis italica). Italy, Basilicata Region, Potenza Province: Massiccio del Pollino, ca. 12 km NNW of Castrovillari, E of Serra del Prete, V. del Malvento, 1570–1600 m s.m., 39°54'30" N 16°08'10" E, 100 m, June 18, 1997, G. Cesca et al. no. 1806 (B101252370, sub O. caryophyllacea); “Pollino National Park, May 30, 2013” (http://www.natureofitaly.com/page716.html by Uvino Platino in Nature of Italy, accessed May 2016 [parasitic on Sideritis italica]).
Taxonomic notes
Orobanche teucrii subsp. sideritidis was collected on June 11, 1877, and was first distributed in exsiccata by Huter, Porta, Rigo (“N.º 357, Huter, Porta, Rigo ex itinere italica III”) under the name of Orobanche Sideritidis [sic], on the same label they also refer to a future description [“Descriptio serius”] of this species. As a taxonomic anecdote about the origin of its name, we can indicate that only know a sheet (CLF170345; Figure 12) that seems to be part of the same gathering, and therefore also syntype, which bears in its label the name of “Orobanche sideritis Huter” and as collectors to Porta and Rigo (Sánchez Pedraja et al., 2016+). The species was published later, but not validly (nomen nudum), such as O. Sideritidis by Porta (1879: 268), who indicated the host plant but did not provide a description, diagnosis, or reference to a description or diagnosis.
Figure 11
Orobanche teucrii subsp. sideritidis syntypes (CLF170345) (https://science.mnhn.fr/institution/clf/collection/clf/item/clf170345).

The origin of its erroneous synonymy, in our opinion, is the above-mentioned exsiccata no. 357. First, it was placed by Caruel in the synonymy of O. caryophyllacea (Caruel, 1885: 372). This material was later studied by Beck (1890: 157), who placed it in the synonymy of his O. caryophyllacea as O. caryophyllacea var. typica subf. thyrsiflora. In addition, he indicated that the host (S. brutia) seems to be false (“Fictae videntur”) because it is not among the hosts of O. caryophyllacea (Beck 1890: 160). After these monographers, many other authors followed the same concept for this species (e.g., Beck, 1930: 232; Uhlich & Rätzel, 2024).
A few years later, the taxon was correctly described with the species rank of Huter, Porta, and Rigo (Huter, 1907: 354), who considered it related to O. caryophyllacea and O. teucrii. These two species are very close to each other (Piwowarczyk et al., 2018) and are often difficult to distinguish (Foley, 2001: 63) if the host is not known or if they are not typical specimens, such as those that identify the usual keys (e.g., Foley, 2001: 35; Pusch, 2009: 16). If we attend to their hosts, in all their distribution ranges, then the distinction is clear; in the first case, it is an exclusive parasite of several species of Rubiaceae and the second is exclusive to the genus Teucrium (Lamiaceae) (Sánchez Pedraja et al., 2016+).
The fact that the host is Lamiaceae probably led other authors to consider it synonymous with O. teucrii (Carlón et al., 2008; Govaerts et al., 2021, sub O. sideritis [sic]; Hassler, 2024, sub O. sideritis [sic]), but as we have indicated, this species is only a parasite of species of the genus Teucrium. Although the treated taxon also has a unique and exclusive host plant (Sideritis italica), this specificity is probably the cause of its differentiation. In our opinion, this taxon seems to be a geographical vicariance with a different host association of the plant that is predominantly circumalpine in Italy, although at some point it seems to reach the Central Apennines. Therefore, we prefer to treat it as a subspecies.
In addition to its host and the characteristics indicated by their descriptors, it can be differentiated from O. caryophyllacea and O. teucrii in the following important diagnostic characteristics. First, it differs by its lowest size (up to 30 cm compared to 60 cm) and short and compact inflorescence against long and lax inflorescences in the other. Second, it differs by its usually most robust habit and its most compact inflorescence with flowers of corollas that are more campanulate and wide; in addition, the calyx segments are most conspicuous, about half as long as the corolla. In all three cases, its coloration can be a source of confusion, and we do not consider it a sufficiently diagnostic character.
New combination
Orobanche teucrii subsp. sideritidis (Huter, Porta & Rigo) Ó. Sánchez & Piwow., comb. & stat. nov. Basionym: Orobanche sideritidis Huter, Porta & Rigo in Oesterr. Bot. Z. 57: 354 (1907)
Homotypic synonyms: Orobanche sideritidis Porta in Nuov. Giorn. Bot. Ital. 11: 268 (1879), nom. nud. Heterotypic synonyms: Orobanche caryophyllacea var. typica subf. thyrsiflora (Wallr.) Beck in Biblioth. Bot. 19: 157 (1890), p.p. Type:—ITALY (lectotype designated by Uhlich & Rätzel, 2024: e04155). «A) ‘N. 357 Huter, Porta, Rigo, ex itinere italica III. / Orobanche sideritidis nobis / Descriptio serius / Calabria III orient. in mte Dirupata di Morano / parte austr.-orient. in [de]clivibus graminos petros.ad radices / Sideritis brutia caespitas integra vastans. sol. calcar. 12–1300 m / 11. Junio 1877’. B) ‘Staatl. Museum für Naturkunde Stuttgart / Isotype / Familie: Orobanchaceae / Art: Orobanche sideritidis Huter, Porta & Rigo ex Huter / Staat: Italy / Fundlokalität: Calabria; in mte. Dirupata di Moreno [sic] parte austr. in clivibus graminos. petros. ad radices Sideritsi [sic] brutia caespites integra vastans. sol. calcar. / Höhe: 12–1300 m NN / leg. R. Huter, P. Porta, G. Rigo Konvolut: Hegelmaier, F. 1907 / Sammeldatum: 11.6.1877’. C) ‘Orobanche caryophyllacea Smith / subforma thyrsiflora Wallroth / Martin Engelhardt 15.IX.2004.’ D) ‘Isotypus / Orobanche sideritidis / Huter, Porta + Rigo ex Huter, / Österreichische Botanische / Zeitschrift 57 (1907) / 354 / Arno Wörz 16.9.2004 / Staatliches Museum für Naturkunde Stuttgart’ (STU 78811/2011, Supporting information; after further examination of the situation and with reference to Art. 8.1 ICN, we choose – contrary to the previous labeling – the entire herbarium specimen as a lectotype; isolectotypes: K000759338, P04384679, P04384680, P04384682, P04393731, MPU021195)». Other syntypes: LY0477279, CLF170345.
Distribution and habitat
The overlooked subspecies were described from Dirupata di Morano [northwest of Morano Calabro, Cosenza Province, Calabria Region, Italy] in the southwestern portion of the Italian peninsula. Apparently, it is endemic of the Southern Apennines [Appennino meridionale] and nowadays, only known from a relatively small area, in the provinces of Cosenza and Catanzaro in the region of Calabria and Salermo province, in the Campania region, although it also reaches, in the Pollino National Park, the province of Potenza in the Basilicata Region. What we now prefer to treat as subspecies is an exclusive parasite of the Sideritis italica (Mill.) Greuter & Burdet (= Sideritis brutia Ten. [Lamiaceae]), another endemism from Southern Italy (incl. Sicily), which grows in calcareous soils.
Paspalum dilatatum Poir. (Poaceae)
Synonyms: Digitaria dilatata (Poir.) Coste, Paspalum moluccanum Huber
Contributors: Arkadiusz Nowak, Sebastian Świerszcz
New record
TAJIKISTAN: Dushanbe, city center, N 38.57985, E 68.78069, elev. 735 m, anthropogenic, ruderal vegetation, July 12, 2024, A. Nowak s.n. (OPUN – Herbarium of the University of Opole).
Taxonomic notes
Paspalum belongs to the tribe Paspaleae and subtribe Paspalinae, and is the largest genus in the subfamily Panicoideae, with 330–350 taxa (Galdeano et al., 2016; Soreng et al., 2017). It is easily distinguished from other genera by its spike-like, solitary, digitate, or racemose inflorescence (Norsaengsri & Chataranothai, 2008). Paspalum species are morphologically characterized by their plano-convex spikelets, with the margins of the upper lemma rolled over the palea (Rua et al., 2010). Due to the large number of species and wide morphological variation, the genus has been divided into subgenera, sections, and groups by Zuloaga & Morrone (2005). Paspalum dilatatum can easily be identified by its spikelets with long (1–2 mm) marginal ciliate fringes of silky hairs, more than two racemes (up. to 10), large spikelets of 3–4 mm, and upper florets equal to 2/3 of the length of the spikelet.
Distribution and habitat
Paspalum dilatatum is a plant of South American origin, mainly found in northeast Argentina, Bolivia, southern Brazil, central Chile, Paraguay, and Uruguay (POWO, 2024). It was introduced in many African countries, including North America, Western Europe, and Southern Asia (POWO, 2024). In Middle Asia, it has been reported only in Uzbekistan (Merkulovich, 1941).
During field research in the southern districts of Dushanbe in July 2024, the species was found in city lawns, park paths, and road verges with the entire old and modern center of the city (Figure S2). The population is huge, comprising tens of thousands of individuals that form dense lawns in irrigated areas. Species populations have been observed on pavements on bridges and along the Varzob River, which may pose a severe threat to the rapid spread of this invasive species southward along the river. Notably, floristic surveys conducted in the city area over the last several years have not confirmed the presence of this species. Paspalum dilatatum has rapidly overgrown its anthropogenic habitats.
In Dushanbe, the population of Paspalum dilatatum was accompanied by: Amaranthus blitum, A. retroflexus, Bromus tectorum, Convolvulus arvensis, Cynodon dactylon, Descurainia sophia, Eleusine indica, Lactuca serriola, Malva neglecta, Oxalis dillenii, Poa annua, Sisymbrium loeselii, Sonchus oleraceus and Tribulus terrestris.
Phelipanche pyramidalis (Reut.) Ó. Sánchez & Piwow. (Orobanchaceae)
Synonyms: Phelypaea pyramidalis Reut., Phelypaea gussoneana Lojac., Phelipanche schultzioides M.J.Y. Foley, Orobanche schultzii var. pyramidalis (Reut.) Beck, Orobanche schultzii f. pyramidalis (Reut.) Beck, Orobanche schultzii auct. pl. (e.g., Gilli, 1982; Strid & Tan, 1991)
Contributors: Óscar Sánchez-Pedraja, Renata Piwowarczyk
New record
AZERBAIJAN: Azerbaijan SSR, Talysh, 3 km along the stream below Zuvand, on the nettle (Urtica sp.) May 19, 1982. leg. et det. E.S. Teryokhin n. 82. (LE01188569, sub O. ramosa, in the authors’ opinion, the specimen previously mentioned by Rätzel et al. (2018, sub P. gussoneana [LE01015390]), does not correspond to this species).
Taxonomic notes
Phelipanche pyramidalis (Reut.) Ó. Sánchez & Piwow. was erroneously included by Beck (1890, 1930) as a form or variety of Orobanche schultzii, based on this, among other characters, in its dense and relatively long inflorescence and that the calix-teeth are longer than the calix-tube. These characters indicated here led him and other later authors (e.g., Gilli, 1982; Strid & Tan, 1991) to include this species within his concept of P. schultzii. The same happened with species that had this characteristic and relatively similar habits, but they had nothing to do with the W Mediterranean species (e.g., P. libanotica (P. orientalis), P. heldreichii) (Piwowarczyk et al., 2019; Piwowarczyk et al., 2023). Beck himself had some doubts about his treatment of this taxon, always putting a question mark “?” to his new combination.
Conversely, Beck himself believed that the distinguished F. gussoneana (Lojacono Pojero, 1882; Rätzel et al., 2018) was very similar to the taxon discussed here (Beck, 1890: 113, “cum O. Gussoneana consentiunt”). The authors’ opinion was similar to that of Beck (1890). We think the same about P. schultzioides from Greece and Crimea (Foley, 2008; Rätzel et al., 2017a, 2018). The two last mentioned names must be brought to the synonymy of Phelipanche pyramidalis by nomenclatural priority. The species apparently closest to this seems to be P. oxyloba, from which it would be distinguished by the characters indicated by Reuter (1847), such as its habit, smaller size, less dense inflorescences with fewer flowers, calyx teeth equal or shorter than the tube. The Geneva sheet (G00330493) on which the lectotypification (Rätzel et al., 2017b: 646, Abb. 2) was made seems to contain two different taxa, based in this we believe that the lectotypification should be done on any of the less robust specimens that fit better with Reuter’s description, the more robust (second by left of the specimens placed above and below of the sheet; Figure 12) seem to correspond to a different taxon.
Figure 12
Phelipanche pyramidalis, lectotype, and isolectotypes (G00330494-Boissier-n. SIB 359358/1 [images 1, 2, https://www.ville-ge.ch/musinfo/bd/cjb/chg/adetail.php?id=407748&base=img&lang=frand]).

New combination
Phelipanche pyramidalis (Reut.) Ó. Sánchez & Piwow., comb. nov.
Basionym:—Phelypaea pyramidalis Reut. in DC., Prodr. 11: 7 (1847).
Homotypic synonyms:—Orobanche schultzii var. pyramidalis (Reut.) Beck in Engl., Pflanzenr: 96: 92 (1930); O. schultzii f. pyramidalis (Reut.) Beck in Biblioth. Bot. 19: 112 (1890).
Heterotypic synonyms:—Phelypaea gussoneana Lojac. in Naturalista Sicil. 1: 199 (1882); Phelipanche schultzioides M.J.Y. Foley in Bot.Chron. (Patras) 19: 7 (2008). Types:—Turkey (lectotype designated here): 1. “Orobanche / (fleurs blues) / Tmolus [Bozdağ Mountains] supra Philadelphiam Sun”. – 2. “Phelipaea pyramidalis Reuter in / DC. Prod. [m. Reuter]”. – 3. “holo-Typus 1/2” (G00330494-Boissier-n. SIB 359358/1 [number G00330494-Boissier-n. SIB 359358/1 is composed of two sheets, image 1 and image 2, we choose the lectotype on image 1 and the lectotype is the second specimen on the right of the six that contains the sheet]; Figure 12); isolectotypes: idem (G00330494-Boissier-n. SIB 359358/1 [image 1, the rest of the specimens that are not the lectotype]), “Tmolus” (G00330494-Boissier-n. SIB 359358/1 [image 2, the four specimens that this sheet contains]; Figure 12); paralectotypes: “Mesogis (Aydos Da), vi 1842, Boiss.” (K000061352 [two equal specimens from same gathering, the left specimen is very representative of this species]; K000061353 [two equal specimens from same gathering, which were reviewed by Beck sub O. schultzii v. pyramidalis Beck (m. Beck)]; P04384654 [four identical specimens from the same gathering, with two identical labels each accompanying two of these specimens]; P02968069 [three equal specimens from the same gathering]); MW0747817 [only one specimen, it is not very representative].
Studies specimens and mentions
CYPRUS: In m. Pentadactylos, May 1880 (WU0028130-Sintenis et Rigo Iter cyprium 1880 no. 632, sub O. ramosa [determined by Beck as “Orob. schultzii (Phel. Pyramidalis Reuter) (Beck)” and by H. Abu-Sbaih as O. schultzii]; P04393737-Sintenis et Rigo Iter cyprium 1880 no. 632, sub O. ramosa; P02968073-Sintenis et Rigo 1880 no 632, sub “Phelipaea. Ramosa?”; P02968074-Sintenis et Rigo 1880 no. 632, sub O. ramosa); pr. Rhizo-Carpass, April 24 (P02982350 – Sintenis et Rigo Iter cyprium 1880 no. 38, sub P. aegyptiaca [only the three specimens near their label]); (Beck, 1930: 92, sub O. schultzii var. pyramidalis).
GREECE: (Boissier, 1879: 498, sub P. pyramidalis; Beck, 1890: 113, sub O. schultzii f. pyramidalis; Beck, 1930: 92, sub O. schultzii var. pyramidalis; Strid & Tan, 1991: 267, sub O. schultzii; Foley, 2008: 7, sub P. schultzioides [E00246315, E00214873]). Crete: Kato Symi, April 16, 2016 (https://www.inaturalist.org/observations/2963532 by Steve Daniels, sub P. mutelii).
ITALY: Sicily: Busambra (Palermo), on Rumex triangularis DC., June 1878, Lojacono (P04384573-Lojacono 1878, sub Phelipaea rufescens); (Lojacono Pojero, 1882: 200, sub P. gussoneana; Rätzel et al., 2018, sub P. gussoneana).
CRIMEA: Eastern Crimea, Karadag, forest along the slope of the Karadag riverbed, June 28, 1928, D. Syreyshchikov n. 568 (MW0626351, on Urtica, sub O. ramosa); Perevalne, July 7, 1980, T. Vylegzhanina (MW0626341, sub O. mutelii); (Rätzel et al., 2017a: 302, sub P. schultzioides).
LEBANON: Horsh Ehden Nature Reserve, North Lebanon, Forêt Ehden 1–5, s. coll., sub Orobanche (photos [“Orobanche Forêt Ehden”]!); Wadi Ramayah bei Nabay, April 17, 2003, Böcker, Nr. LIBA 4 (B 10 0626295, as O. ramosa L., rev. Rätzel, July 24, 2019, P. gussoneana var. gussoneana; Rätzel et al., 2020, sub P. gussoneana).
RUSSIA: Krasnodarskiy Kray, Anapskiy rayon, Abrau Peninsula, Navagirskiy Khrebet, May 25, 2018, Ryabenko & Tokavera (MW1008942, sub O. purpurea) (Rätzel et al., 2018, sub P. gussoneana).
SYRIA: col de Freiket dans les Monts Nosaïris, July 12, 1939, apparently on Lamium?, Frère Louis n. 554 (P04242361, sub Phelipaea ramosa and O. mutelii); (Boissier, 1879: 498, sub P. pyramidalis; Beck, 1890: 113, sub O. schultzii f. pyramidalis; Beck, 1930: 92, sub O. schultzii var. pyramidalis).
TURKEY: Tmolus [Bozdağ Mountains] and Mesogis (Aydos Da), vi 1842, Boissier (Reuter, 1847: 7, sub Phelypaea pyramidalis; Boissier 1879: 498, sub P. pyramidalis; Beck, 1890: 112, sub O. schultzii f. pyramidalis; Beck, 1930: 87, sub O. schultzii var. pyramidalis; Gilli, 1982: 10, sub O. schultzii).
Distribution and habitat
The species was described from the Mount Tmolus [Bozdağ Mountains], above Alaşehir [Philadelphia, Manisa Province, Aegean Region, Turkey] by (Reuter, 1847: 7, sub Phelypaea pyramidalis). It has an Oriental ± Mediterranean distribution: Azerbaijan, Cyprus, and Greece (incl. Crete), Italy (Sicily), Crimea, Lebanon, Russia, Syria, Turkey (Boissier, 1879; Beck, 1890; Beck, 1930; Gilli, 1982; Strid & Tan, 1991; Rätzel et al., 2020). In addition, we believe that the French mentions of this taxon are apparently incorrect.
Phelipanche reuteriana (Rchb. f.) Carlón, G. Gómez, M. Laínz, Moreno Mor., Ó. Sánchez & Schneew. (Orobanchaceae)
Synonyms: Phelypaea reuteriana Rchb. f., Orobanche tunetana Beck, Orobanche aegyptiaca subsp. tunetana (Beck) Maire, Orobanche tunetana var. tacassea Beck
Contributors: Óscar Sánchez Pedraja, Renata Piwowarczyk
New records
EGYPT: Champs, Alexandrie, Egyptia, March 8, 1847 (US147282-03927206-ex herb. L. Kralik [as Phelypaea aegyptiaca Walp.]);
ETHIOPIA: Debr-Eski [Dabra Eski/Däbr Eski, towards Woina, Semien Mountains], Abyssinie, July 10, 1852, sur Cynoglossum [Boraginaceae], M.W. Schimper n. 227 (P02970733 [as Phelypaea aegyptiaca Walp.]); Debr-Eski [Dabra Eski / Däbr Eski, towards Woina, Semien Mountains], Abyssinie, envoi de M.W. Schimper 1853 n. 226 (P02970734 [as Phelypaea aegyptiaca Walp.]);
MALI: Kidal Region: French Sudan: Kidal (Adrar des Ifoghas), February 5, 1933, Fronteau n. 6 (P02970726-hb. Aug. Chevalier [as Orobanche aegyptiaca Pers. var. lasianthera Maire]); Timbuktu Region: Soudan [French Sudan] (Sahara): Tombouctou [Timbuktu / Tumbutu], on Nicotiana rustica, March 7, 1932, A. Chevalier n. 43837 (P04392556 [as O. aegyptiaca Pers. var. lasianthera Maire]); idem (P02970727-hb. Chevalier-1931-32); idem (P02970730-hb. Chevalier-1931-32 [Mali’s specimens are more robust than typical (see var. lanuginosa Maire)]).
Taxonomic notes
Reichenbach f. (1862), when he described Phelypaea reuteriana Rchb.f. from Spain, he thought he was facing a very similar species, especially in terms of its indumentum, to the species that his father had described from Russia (Orobanche caesia), and compared it to the Russian species. Later, Beck (1890), on material collected by Kralik in Tunisia, described what he considered a new species, Orobanche tunetana, that also compared to Russian species (O. lanuginosa (= O. caesia)). Some years later, Maire (1929) subordinated the species discussed here to O. aegyptiaca with the rank of variety, and created a new combination (O. aegyptiaca var. lasianthera). Maire himself (1935) identified its variety with the species of Beck and raised its rank to subspecies, creating a new combination, O. aegyptiaca subsp. tunetana (Emberger & Maire, 1941). Pujadas et al. (1997) rediscovered this species in the European continent (Spain) and considered it conspecific to Beck’s O. tunetana from North Africa. Eight years later, Carlón et al. (2005) identified this Spanish Levante plant with that described by Reichenbach f., and indicated the priority of his name in the specific rank, over the Beck’s name, typifying and combining this one within the Phelipanche genus as P. reuteriana.
Other species closely related to this have been described in Saudi Arabia (P. hypertomentosa) by Foley (1998) and Israel (P. daninii) by Domina & Raimondo (2009), which seem to fit into the variability of the species, and it may be more appropriate to take them as synonyms of the species treated here.
In addition, we believe that Saharian material fits better in what Maire (1935) described as var. lanuginosa, according to the characters indicated by the author. Apparently, Mali’s material collected in 1932–1933 and determined by Maire to be O. aegyptiaca var. lasianthera corresponds to the same variety, but we have not been able to find material before the year of its description (1929) undoubtedly referred to this variety that Maire himself considers synonym of O. tunetana (≡ O. aegyptiaca subsp. tunetana var. eutunetana (Emberger & Maire, 1941)).
Phelipanche reuteriana var. lanuginosa (Maire) Ó. Sánchez & Piwow., comb. nov.
Basionym: Orobanche aegyptiaca [subsp. euaegyptiaca] var. lanuginosa Maire in Bull. Soc. Hist. Nat. Afrique 26: 220 (1935 [July 15, 1935]).
Lectotype (here designated): “Université d’Alger / Herbier de l’Afrique du Nord / Orobanche aegyptiaca Pers. / Hoggar [Ahaggar, N 23°10'00" E 5°50'00"], Tamanghasset [Tamanrasset / Tamenghes, Algeria] dans la steppe de Atriplex halimus L. [Amaranthaceae Juss. (incl. Chenopodiaceae Vent.)], sur? / 20-1-1933 / leg. F. Lauriol / [det.] Dr. R. Maire”. – 2. “O. tunetana G. Beck / Determinavit / Anno 2000 A. Pujadas” (MPU059347). Other representative materials of this variety are P04363439, P04076068, and MPU059346 (Figure 13).
Figure 13
Phelipanche reuteriana var. lanuginosa, representative material (MPU059346) (https://science.mnhn.fr/institution/um/collection/mpu/item/mpu059346).

Distribution and habitat
Phelipanche reuteriana was described from Elche/Elx (Alicante/Alacant, Spain) by Reichenbach f. (1862). However, despite its description, the distribution is mainly North African, occupying only a relatively small area of the Spanish Levante, which is the northern limit of its distribution range, whereas the southern limit is the Sahara desert. Its known distribution spreads through the following countries: Algeria, Egypt, Ethiopia (Abyssinia), Mali, Mauritania, and Morocco (incl. Western Sahara), Spain, and Tunisia (Sánchez Pedraja et al., 2016+). Its habitat, as indicated by its distribution, is warm, dry, and frequently desert areas, on sandy lands or very degraded marls (Pujadas et al., 1997), where it parasites different families, for example, Plantaginaceae in the Iberian Peninsula and Brassicaceae in Africa (Sánchez Pedraja et al., 2016+).
With the discovery in herbaria P and the US, we expanded the distribution of this species to Egypt, Ethiopia, and Mali; these countries are the southern and eastern limits of its range (Pujadas et al., 1997; Dobignard & Chatelain, 2013; Chatelain et al., 2022).
Portulaca granulatostellulata (Poelln.) Ricceri & Arrigoni (Portulacaceae)
Synonyms: Portulaca oleracea L. var. granulatostellulata Poelln., Portulaca oleracea L. subsp. granulatostellulata (Poelln.) Danin & H.G.Baker
Contributors: Piotr Szkudlarz, Zbigniew Celka
New record
MADAGASCAR: Lalana Fredy Rajaofera Street, Antananarivo, 18.903932°S, 47.529154°E, elev. 1280 m, ruderal site, April 9, 2022, P. Szkudlarz [POZ-C-0004760].
Taxonomic notes
Portulaca is a cosmopolitan genus with over 150 taxa that forms the monotypic family Portulacaceae (APG, 2016; Nyffeler & Eggli, 2010; POWO, 2024). In older systems, Portulaca was joined with other genera into a broadly described family composed of ca. 450 species (Carolin, 1993; Takhtajan, 2009). Numerous taxa included in Portulaca oleracea agg. can be distinguished from one another based only on the basis of their seed characteristics. The seeds of P. granulatostellulata are reniform and black, with a shiny surface. Their diameter is below 0.85 mm and their mean length varies from 0.703 mm to 0.849 mm (general mean 0.733 mm). Epidermal cells of the seed testa are stellulate and elongated with relatively short arms. The cell surface is verrucose, with verrucae located mostly in the distal parts of the cell arms (e.g. Danin et al., 2012; Ocampo, 2013; Bulakh et al., 2022) (Figure 14).
Figure 14
Portulaca granulatostellulata (SEM). General view of the seed (A), general view of the testa cells (B).
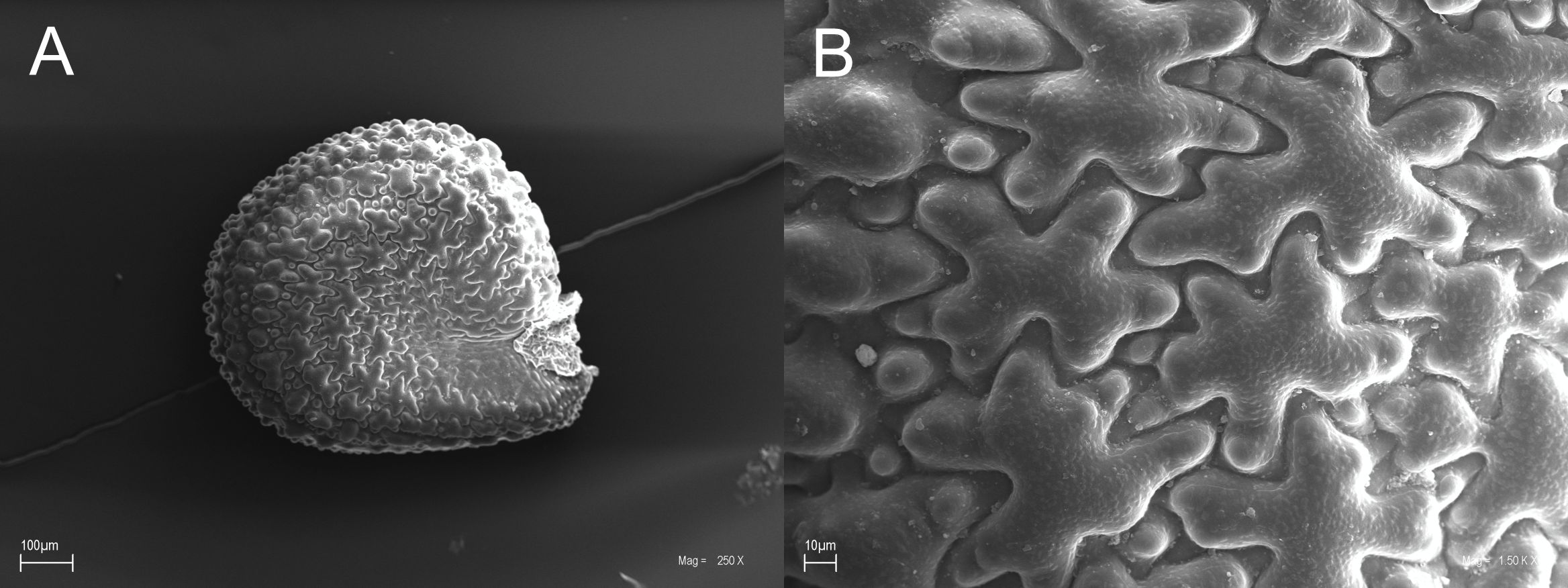
Distribution and habitat
The natural range of Portulaca granulatostellulata includes the Mediterranean region, Central Europe to Caucasus, Kyrgyzstan, and Africa. In Great Britain, North Europe, and East Europe, it is classified as introduced (POWO, 2024; WFO, 2024). It is considered to be the most common species of Portulaca oleracea agg. in Central Europe (Hassler, 1994–2024). However, this annual semi-succulent plant grows primarily in subtropical biomes (POWO, 2024). To date, only P. oleracea has been reported from Madagascar, from approximately a dozen localities at elevations of up to 500 a.s.l. (CPM, 2024), which comprises ruderal sites, cultivated fields, coastal rocks, muddy soil, and salt marshes (CPM, 2024; Hughes & Hughes, 1992). However, P. granulatostellulata has been reported from islands surrounding Madagascar, e.g. Reunion (GBIF, 2023). During floristic research in Madagascar, the last mentioned species was found in the capital city, Antananarivo, on the edge of a road, roadside, and between pavement blocks in the city center, in Lalana Fredy Rajaofera Street (Figure 15). The site is located at an elevation of ca. 1400 m a.s.l. The observed population consisted of about a dozen individuals growing in more or less dense patches. The reported species was accompanied by single individuals of other species, e.g. Alternanthera caracasana, Eleusine indica, Erigeron canadensis, and Oxalis corniculata.

Ptilagrostis chingii (Hitchc.) M. Nobis & Krzempek (Poaceae)
Synonyms: Stipa chingii Hitchc., Achnatherum chingii (Hitchc.) Keng, Achnatherum chingii var. laxum S.L.Lu
Contributors: Marta Krzempek, Marcin Nobis, Ewelina Klichowska
New records
BHUTAN: N 19 (Upper Pho Chu), Gasa. Thangkaphu Chu/Tsharijathang junction, open Abies-Betula forest on NE-facing slope, Takin pasture and rest places, alt. 4070 m, 27°59'N 89°32'E, July 23, 00, G. & S. Miehe (E00180485); N 19 (Upper Pho Chu), Gasa, Tarina Camp, Abies densa forest on S-facing lower slope, alt 4040m, 28°3'N, 89°57'E, September 1, 00, G. & S. Miehe (E00180448).
Taxonomic notes
Ptilagrostis is a genus of high-alpine grasses found in Central and Northeast Asia and North America (Barkworth, 1983; Wu & Phillips, 2006; Tzvelev & Probatova, 2019). Currently, 15 species and one subspecies are distinguished within this genus (Krzempek et al., 2024). Described by Hitchcock (1930) Stipa chingii was firstly transferred to the genus Achnatherum by Yi Li Keng (1976), and recently to the genus Ptilagrostis as Ptilagrostis chingii (Krzempek et al., 2024). Plants representing this species are densely tufted, perennial, erect, 31–116 cm tall stems, filiform and convolute leaves, open or loosely contracted panicles with purple or yellowish green, subequal glumes, and 1-geniculate awns (Wu & Phillips, 2006; Zhang et al., 2016; Krzempek et al., 2024). Specimens with contracted panicles are distinguished as typical P. chingii var. chingii, whereas those with open panicles can be identified as P. chingii var. laxum (Krzempek et al., 2024). Ptilagrostis chingii is described as being highly similar to P. duthiei, varying only in anther size (Wu & Phillips, 2006). However, our recent research has shown that the best features to distinguish the two species are the number of vascular bundles in the vegetative leaf and the width of the leaf. Ptilagrostis chingii has very narrow leaves with three or five conductive bundles, whereas P. duthiei has noticeably wider leaves, with 7–11 vascular bundles. These two species, together with the recently described P. contracta (Zhang et al., 2017), have been included in the new section Chenella, within the genus Ptilagrostis. Currently in Bhutan, the genus Ptilagrostis is represented by the following taxa: P. dichotoma, P. bhutanica, P. yadongensis (syn. Stipa milleri, P. milleri), P. chingii, and P. duthiei (names are given after Krzempek et al., 2024).
Distribution and habitat
Originally, P. chingii was described by Labrang in the Gansu Province of China (Hitchcock, 1930). To date, this species was believed to have occurred solely in China. It has been reported in the provinces of Gansu, Qinghai, Shaanxi, Shanxi, Sichuan, Xizang, and Yunnan. P. chingii occurs in Abies forests in the alpine zone at an elevation of 2000–4100 m (Wu & Phillips, 2006; Zhang et al., 2016; Krzempek et al., 2024). Recently, during taxonomic revision of herbarium materials preserved at the Royal Botanic Garden Edinburgh Herbarium (E), we found two specimens of P. chingii collected in Bhutan that were misidentified as P. bhutanica. In both cases, plant material was collected in the Gasa district on mountain slopes over 4000 m above sea level in Abies forests. P. chingii is a new native species in the flora of Bhutan.
Reynoutria japonica Houtt. (Polygonaceae)
Synonyms: Polygonum cuspidatum Siebold & Zucc.; Fallopia japonica (Houtt.) Ronse Decr.
Contributors: Arkadiusz Nowak, Sebastian Świerszcz
New records
TAJIKISTAN: Kante, home garden, N 39.23181, E 68.50187, elev. 2180 m a.s.l., July 11, 2024, A. Nowak s.n.; Dar-dar, along riverside in the village, N 39.43805, E 68.44697, elev. 1396 m a.s.l., July 11, 2024, A. Nowak s.n. (OPUN – Herbarium of the University of Opole).
Taxonomic notes
Reynoutria japonica is a perennial, dioecious herb. It has thickened rhizomes and erected stems. Leaves simple, alternate, petiolate; leaf blade ovate or ovate-elliptic up to 15 cm long, margin entire; ocrea oblique. Sparse glandular hairs were present on the underside of the leaf or the leaves are glabrous. The hairs are barely visible, reduced to short unicellular papillae with a distinct base. The inflorescence axillary, paniculate, with two to four flowers in one bunch.
Distribution and habitat
Reynoutria japonica belongs to the most widespread and most invasive weed species in the world. It grows naturally in Japan, Taiwan, and northern China (Mandák & Pyšek, 1997). It has been cultivated in Europe since 1825 (Mandák & Pyšek, 1997) and is now an established species in many countries, especially in Central and Western Europe (Jalas & Suominen, 1988). This species has never been recorded in Middle Asia.
During fieldwork in western Tajikistan, we found two populations of this species on the southern slopes of the Zeravshan (Kante village) and Turkestan (Dar-dar village) mountains. In Kante (Figure 16), a few individuals grew in private gardens with ornamental plants, mainly Cosmos spp. The homeowners said that the species had not been cut, but it was already known in the region as a weed. In Dar-dar, approximately 20 specimens grew along the stream in the village, mainly on gravel bars with ruderal and riparian species, such as Salix spp., Myricaria spp., Datisca cannabina, and Geranium divaricatum.

Spinulum annotinum (L.) A. Haines (Lycopodiaceae)
Synonyms: Lepidotis annotina (L.) P.Beauv.; Lycopodium annotinum L.; Lycopodium juniperifolium Lam.
Contributors: Daniyar Alibekov, Serik Kubentayev
New record
KAZAKHSTAN: Kokchetav: Akmola Region, Burabay District, SNNP “Burabay”. Near lake Svetloe, Nord-West shore, 53°02'35.1"N / 70°11'08.7"E, July 11, 2019, S.A. Kubentayev (NUR).
Taxonomic notes
Spinulum annotinum was previously assigned to the genus Lycopodium L., and in 2003 was transfered to the new genus Spinulum A. Haines (Haines & Vining, 2003). However, some authors still refer to it as the genus Lycopodium (Gola et al., 2015; Rimgailė-Voicik et al., 2024). This species is morphologically close to Lycopodium clavatum L., from which it can be distinguished by its pointed, horizontally spaced, rather stiff leaves (phyllodes), with sparsely serrate margins. L. clavatum has linear or linear-lanceolate, entire margins, passing into long, upwardly bent white hairs. Moreover, S. annotinum forms single sessile strobiloids, up to 3 cm long, 4-5 mm wide, whereas in L. clavatum, spore-bearing spikelets are mostly 2, less often 3-4 or solitary, linear-cylindrical, on long thin stalks (Ilyin, 1934; Kharkevich, 1985). In addition, S. annotinum can be determined to some extent by annual growth compression of the rhizomes (Primack, 1973).
Distribution and habitat
Spinulum annotinum is a circumboreal Holarctic species. There is information that the statement about the occurrence of S. annotinum in East Asia, including Korea, Japan, and the Himalayas, is incorrect, because only Spinulum lioui Li Bing Zhang & H.He grows there. In contrast, S. annotinum in East Asia grows only in the Russian Far East (Wan et al., 2023). The first information on the occurrence of S. annotinum in Kazakhstan is given in ‘Flora of Western Siberia’ (Krylov, 1927), where three localities of this species are Eastern Kazakhstan, Southern Altai, near Lake Markakol, Chebambai, and Rakhmanovskie kluchi. However, in ‘Flora of Kazakhstan’ (Pavlov, 1956), only one locality is given near Lake Markakol, with reference to the materials of geographer-researcher A. N. Sedelnikov. Spinulum annotinum was first listed in Western Altai (Eastern Kazakhstan) in 2011 (Baitulin & Kotukhov, 2011). However, in Northern Kazakhstan (Arakaragai forestry, Altynsarinsky district, Kostanay region), S. annotinum was first listed in 2008 (Perezhogin et al., 2009), although the herbarium material of that species was collected over 30 years ealier (in 1976). This was the only known locality of S. annotinum in Northern Kazakhstan until we found this species in the Akmola Oblast. Information on the occurrence and status of S. annotinum populations in Arakaragai forestry is unknown; no one has observed this species after 1976. It should be noted that, in September 2022, a large fire occurred in the Arakaragai pine forest, and its consequences on the population of this relict species have not been studied. Although our finding of S. annotinum is the second confirmed locality of this species in Northern Kazakhstan, the location of S. annotinum is new for the flora of the Akmola region, Kokchetav floristic district, and in general for the Kazakh Uplands (Pavlov, 1956; Karamysheva & Rachkovskaya, 1973; Kupriyanov, 2020). The new locality is more than 400 km from the known location of S. annotinum in the Kostanay region (Perezhogin et al., 2009). In the locality, we found S. annotinum growing on wet mossy bumps in a mixed pine-birch forest near Lake Svetloye (Figure 17) in the protected area of the State National Nature Park ‘Burabai’. In our opinion, S. annotinum has not been included in the list of the flora of the Kazakh Uplands because of its rarity and erroneous identification as L. clavatum, which also occurs in Northern Kazakhstan (Gorchakovskiy, 1987; Kupriyanov, 2020). Thus, it has been established that both species, S. annotinum and L. clavatum, grow in the Kazakh Uplands, but it is necessary to further study the distribution of these species in Northern Kazakhstan.

Stipa borysthenica Klokov ex Prokudin (Poaceae)
Synonyms: Stipa pennata f. sabulosa Paczoski; Stipa joannis subsp. sabulosa (Paczoski) Lavrenko; Stipa sabulosa (Paczoski) Sljussarenko; Stipa pennata subsp. sabulosa (Paczoski) Tzvelev; Stipa baktashevae Tzvelev; Stipa joannis var. marchicae Endtman; Stipa borysthenica var. marchicae (Endtman) Rauschert
Contributors: Ewelina Klichowska, Marcin Nobis
New record
ARMENIA: Zangezur Mountains Nothern parts of Bargushat Mts, between rocks, E slopes, elev. 2400 m, August 3, 1957, Akhverdov & Mirzoeva (KRA 634315).
Taxonomic notes
Described by Linnaeus (1753) genus Stipa is one of the largest genera in the family Poaceae, subfamily Pooideae, comprising over 150 species, all native to the Old World (Nobis et al., 2020a). Stipa borysthenica belongs to the nominal section and is characterized by a bigeniculate awn with a plumose upper segment (Tzvelev, 1974). The taxon was described as a form of S. pennata f. sabulosa (Paczoski, 1914) and later underwent multiple nomenclature combinations (Prokudin, 1951; Sljussarenko, 1963; Gonzalo et al., 2013).
Stipa borysthenica differs from that in Armenia, the most closely related species, S. pennata, by having a longer callus (3.4–)3.8–4.3(–5.0) mm vs. (2.4–)2.8–3.3(–3.7) mm, which is straight or slightly curved with a narrow, cuneate-shaped peripheral ring vs. curved callus with a wider, pear-shaped peripheral ring (Gonzalo et al., 2013; Klichowska & Nobis, 2018). Stipa borysthenica also differs in the last before panicle cauline leaves, which are slightly longer (2.2–)3.6–6.2(–12.5) cm than those of S. pennata (0.4–)1.0–2.2(–4.0) cm (Klichowska & Nobis, 2018). Less likely, confusion may occur with other species found in Armenia, such as S. tirsa (= S. stenophylla) or S. pontica (the nomenclature is given after Nobis et al., 2020a). From the first of them, S. borysthenica differs by the glabrous or slightly scabrous abaxial surface of basal leaves vs. basal leaves with setaceous apex and sparsely stiff hairs on the abaxial surface, as well as longer ligules of basal leaves (0.9–3.4 mm vs. 0.1–0.4 mm). However, S. pontica, despite having a similarly long anthecium reaching 17 mm, unlike S. borysthenica, has more or less appressed hairs on the abaxial surface of leaves and admixture of long (up to 0.5 mm) hairs on the adaxial surface of the leaves as well as lemma with a ventral line of hairs reaching the base of the awn (Martinovský, 1980).
Here, we combined morphological and molecular methods (DArTseq) for the proper identification of Armenian specimens that were found during herbarium (ERE) revision in 2023 (Figure 18; Supplementary matrials). Based on the morphological characteristics, we identified one specimen as S. borysthenica. In the case of the second specimen, which showed morphological ambiguity, by combining morphological and molecular data, we confirmed for the first time the possibility of gene flow between closely related S. borysthenica and S. pennata. The hybrid (S. borysthenica x S. pennata) has only been identified in one individual from the herbarium collection [ARMENIA: Martuninskii region, SW parts of Airidzha Yanykh, July 18, 1954, Sh. Aelanyan, Ts. M. (ERE 150425); however, this has not been observed in the field. As we have previously demonstrated, DArTseq is a powerful method for investigating hybridization and introgression in the genus Stipa (Baiakhmetov et al., 2020; Sinaga et al., 2024); however, in this case, we believe that further research is needed to confirm that this hybridization is not a single event.
Figure 18
Genetic relationship between S. borysthenica and S. pennata. (A) UPGMA tree for 30 individuals of S. borysthenica and S. pennata based on 8435 DArTseq-derived SNPs (with S. capillata as an outgroup). The Armenia specimen is marked in red. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches; (B) A FastSTRUCTURE bar graphs for K2 based on 7862 DArTseq-derived SNPs (without an outgroup). The blue bars correspond to S. pennata individuals, and orange bars correspond to S. borysthenica.
Distribution and habitat
Stipa borysthenica is widely distributed in the steppe belt, extending from Central Asia (Mongolia, Russia, and Kazakhstan) to Eastern Europe (Ukraine). The northwesternmost limit of its range reaches the territories of Germany, the Czech Republic, and Poland. In the south, it reaches the southwest regions of Russia on the Caspian Sea Coast (Republic of Kalmykia and Dagestan), the southeast regions of Ukraine (including Donetsk and Zaporizhzhia regions), as well as the territory of Crimea, the eastern areas of Bulgaria on the Black Sea Coast, and geographically isolated localities in Iran separated from the main range (Gonzalo et al., 2013). The species has never been previously reported in Armenia since herbarium specimens were misidentified as S. pennata, S. tirsa, or S. pontica. However, the species could also be expected here, and its localities in Armenia fill the gap in the geographical range between the continuous range and the isolated locality in Iran.
Stipa borysthenica occurs exclusively on sandy substrates in open habitats such as steppes and grasslands on slopes, rivers, and roadsides (Gonzalo et al., 2013). This distinguishes it from the most similar Stipa pennata, which grows mainly in shallow soils with limestone, gypsum, or chalk bedrock.
Stipa drobovii (Tzvelev) Czerep. (Poaceae)
Synonyms: Stipa caucasica subsp. drobovii Tzvelev
Contributors: Marcin Nobis, Ewelina Klichowska
New records
ARMENIA: Arazdanyanskii steppe, on a small hill of Danga Mt., June 2, 1948, Akhverdov & Mirzoeva s.n. (ERE150416-150424, KRA 634209).
Taxonomic notes
Stipa drobovii belongs to the Stipa section of Smirnovia and is characterized by the presence of unigeniculate and pilose awns (Nobis, 2012; Nobis et al., 2020a; Sinaga et al., 2024). Tzvelev (1974) described Stipa drobovii in the rank of subspecies as Stipa caucasica subsp. drobovii Tzvelev, but later raised it to the rank of a species by Czerepanov (1981). It can be distinguished from other members of the C. caucasica group, including S. caucasica subsp. caucasica, which also occurs in Armenia, by having a shorter callus 0.8–1.2(–1.5) mm, as well as hairs in the dorsal part of the callus that are strongly falcate and longer than the ventral ones, versus callus (1.6–)1.8–3.5(–4) mm long, and hairs in the dorsal part of the callus that are straight or flexuous and shorter than the ventral ones (Gonzalo et al., 2011; Nobis et al., 2020a). Within S. drobovii, the four, following lower rank taxa are recognized: S. drobovii var. drobovii, S. drobovii var. jarmica, S. drobovii var. persicorum, and S. drobovii var. iskanderculica (nomenclature given by Nobis et al., 2020a). Armenian specimens are the most similar to the typical variety; however, they differ in having upper culm leafe sheaths pilose along vains; thus, they can be distinguished as a new variety:
Stipa drobovii var. hajastanica M. Nobis & Klichowska, var. nov.
Description: Culms 18–34 cm high. Sheaths of upper cauline leaves in the upper part pilose along ribs (veins) with up directed (0.3–)0.7–1.5 mm long hairs, between ribs scabrous, at margins white and glabrous, the lower sheaths glabrous or glabrous in the lower part of sheath and pilose in the upper. Vegetative leaves abaxially glabrous and smooth, adaxially densely pilose with 0.25–0.4 mm long hairs. Anthecium 9.0–11.5 mm long. Awn 60–90 mm long, unigeniculate; column 15–22 mm long; seta 45–70 mm long.
Type: ARMENIA: Arazdanyanskii steppe, on a small hill on Danga Mountain, [900 m elv.], June 2, 1948, Akhverdov & Mirzoeva s.n. (holotype: ERE150416!; isotypes: ERE150417!, ERE150418!, ERE150419!, ERE150420!, ERE150421!, ERE150422!, ERE150423!, ERE150424!, KRA 634209!); Figure 19. Paratypes: Dzhul’finskii raion, 4-yi km ot Dzhul’fy k Yaidzhy, sukhie kholmy Dary-gaga, June 9, 1985, N.S. Khandzhyan & V.A. Manakyan (ERE150075!); District nakhichevan, inter Dzhalfa, et Darosh[ah?], June 4, 1923, Grossheim 11306 (ERE!); District nakhichevan, inter Dzhalfa, et Aza, May 24, 1923, Grossheim 11308 (ERE!); Nakhkrai, peski u SW podnozhya g. Danga, 1000 m elev., May 30, 1948, Akhverdov s.n. (ERE150073, ERE150074!).

Distribution and habitat
Representatives of the genus Stipa s str. are widely distributed in the warm temperate regions of the Old World (Nobis et al., 2020a). Stipa drobovii occurs in Central and West Asia, from Kazakhstan, Kyrgyzstan, and Tajikistan through Turkmenistan and Azerbaijan to northern Iran. Although this taxon was previously noted in the Caucasus region, the newly discovered site in Armenia is the westernmost (Tzvelev, 2006). The taxon was also mentioned in the Flora of Armenia (Gabrielyan, 2009) but has been placed within the synonyms of S. caucasica. Stipa drobovii var. hajastanica grows on arid steppes and semideserts in lower mountain elevations, usually on calcareous clay soils.
Stipa zuvantica Tzvelev (Poaceae)
Contributors: Marcin Nobis, Ewelina Klichowska
New record
Armenia: Armenia, between Jeghegandzor and Malishka, steppes on the hill, 39°44'38.62"N / 45°21'42.87"E, elev. 1150 m, exp. SW, incl. 20°, June 17, 2023, wp. 1650, M. Nobis, E. Klichowska s.n. (KRA 634215-634221).
Taxonomic notes
Stipa zuvantica belongs to section Barbatae A. Junge. This was described based on the collection of C.A. Mayar from the Zuvant region in Azerbaijan (specimen collected in 1830 and preserved at the LE!). This species was described by Tzvelev (1966) from Azerbaijan (Zuvant). Later, it was found only in Turkey in Konya (Obruk Yaylasi between Konya and Sultanhani, alt. 1000 m, Central Anatolia; Scholz in Greuter & Raus, 1998). During field studies in the Tstanscaucasian steppes, we found this species in Armenia. From the most similar species representing the section Barbatae growing in the country, S. zuvantica can be distinguished as follows:
1. Lower segment of the awn (column) scabrous, covered with 0.07–0.1(–0.2) mm long prickles ………………………… 2
– Lower segment of the awn (column) pilose, covered with 0.4–2.0 mm long hairs ………………………… 3
2. Awn stiff, terminal segment of the awn (seta) wavy, shortly pilose, covered with 0.4–0.8(–0.9) mm long hairs, sheaths of the vegetative shoots shortly pubescent, ligules of the vegetative shoots (1.5–)3–6(–9) mm long ………………………… S. zuvantica
– Awn flexuous, terminal segment of the awn (seta) wavy, pilose, covered with 1.5–2.5 mm long hairs, sheaths of the vegetative shoots glabrous, scabrous to shortly pubescent, ligules of the vegetative shoots (2–)3–8(–10) mm long ………………………… S. hohenackeriana
3. Awn stiff, terminal segment of the awn (seta) wavy, distinctly scabrous, covered with 0.1–0.2(–0.3) mm long prickles, sheaths of the vegetative shoots shortly pubescent or glabrous, ligules of the vegetative shoots (0.5–)1–2(–3) mm long ………………………… S. holosericea s. l.
– Awn stiff or flexuous, terminal segment of the awn (seta) wavy, pilose, covered with 1.5–2.5(–3) mm long hairs ………………………… 4
4. Lemma with two distinct rows of hairs, dorsat endings in 1/3–1/2 of lemma length, ventral termilals in 1/2–2/3 of lemma length; awn stiff, terminal segment of awn (seta) wavy; glabrous and smooth leaves, ligules of vegetative shoots 1–2 mm long ………………………… S. ehrenbergiana
– Lemma minutely pilose throughout or with seven indistinct rows of hairs terminating in the upper half of the lemma with two distinct rows of hairs; awn flexuose, terminal segment of the awn (seta) wavy; leaves scabrouos (rarely pubescent), and ligules of the vegetative shoots (1.5–)3–8(–15) mm long ………………………… S. arabica
Distribution and habitat
Stipa zuvantica occurs in southwestern Asia, Azerbaijan, Armenia, and Turkey. However, its occurrence is also possible in Iran. This newly discovered site in Armenia fills the distribution gap between Azerbaijan and Turkey (Tzvelev, 2006). It grows on a stony steppe on the western slope of the hill in the region of Vayots Dzor (Figure 20). Although the species was found accidentally, it is likely that it is more common in this stepic region of Armenia. At the locality, S. zuvantica was found together with the following feather grasses: S. anisotricha, S. araxensis, S. arabica, S. hohenackeriana, S. capillata, S. pontica, S. holosericea var. holosericea and, S. holosericea var. transcaucasica (nomenclature is given after Nobis et al., 2020a).

Taraxacum arrhenii Palmgr. (section Taraxacum, Asteraceae)
Synonyms: Taraxacum amoenum Puol.
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
Taraxacum is one of the largest and most taxonomically complex apomictic genera. Its great diversity is evidenced by over 2800 species described thus far, grouped into about 60 sections (Kirschner & Štěpánek, 2008; Kirschner et al., 2015; Rewicz et al., 2020). According to Kirschner & Štěpánek (1996), the main evolutionary mechanisms include multiple hybridization, polyploidization, and apomixis. In the high latitudes of the Northern Hemisphere, where there are few sexually reproducing species, the appearance of new forms (microevolution) is caused by autopolyploidization processes and somatic mutations fixed by apomixis (Kirschner & Štěpánek, 1996; Mes et al., 2002; Majeský et al., 2012). Genetic variability is compounded by very high phenotypic plasticity, which significantly increases the already serious problems in species identification (Kirschner et al., 2016). These problems, combined with the small number of specialists, have contributed to the poor and uneven level of research on the taxonomic diversity and distribution of Taraxacum. This genus is relatively well-recognized in the British Isles, Scandinavia, the Netherlands, Germany, Poland, the Czech Republic, and Slovakia (Dudman & Richards, 1997; Kirschner & Štěpánek, 1998; Lundevall & Øllgaard, 1999; Uhlemann, 2003, 2015; Trávníček et al., 2010; Danihelka et al., 2012; Marciniuk, 2012; Marciniuk et al., 2012, 2016, 2018, 2019; Trávníček et al., 2018; Mirek et al., 2020; Uhlemann et al., 2020; Wolanin et al., 2023). However, the distribution of Taraxacum in Eastern Europe is poorly understood. In a previous study (Nobis et al., 2020b), we provided new data on the distribution of 17 Taraxacum species that were new to the flora of Ukraine, and below, we provided new records of the next 10 species.
New record
UKRAINE: Lviv, Lychakiv Cemetery, May 8, 2017, J. & P. Marciniuk (WSRP)
Taxonomic notes
Taraxacum arrhenii belongs to the T. arrhenii group. The main diagnostic characteristics of this taxon are as follows: large plant, leaves mid-green, obviously hairy, side lobes recurved, deltoid, medium acute, undivided; the upper edge is straight, usually entire, sometimes with uniform dentation; the lower edge is straight, usually entire, interlobes acute angled without tarry discoloration; the terminal lobe is blunt to acute, approximately triangular, the petioles are intensely red, unwinged or narrowly winged, midrib red to purple with striatulate pattern; outer bracts 3–4 mm wide and 14–15 mm long, horizontally positioned, unbordered; capitulum with a diameter of ca. 50 mm, convex; stigmas discolored; pollen present (Figure 21A).
Figure 21
Herbarium specimen of Taraxacum arrhenii (A) Lviv Lychakiv Cemetery 8.05.2017; T. diastematicum (B), Lviv Lychakiv Cemetery 8.05.2017; T. crassum (C) Lviv prospekt Chervonoi Kalyny 10.05.2017; T. baeckiiforme (D) Wola (District Mykolaiv) 10.05.2017; T. gibberum (E) Wola (District Mykolaiv) 10.05.2017. Scale bar 5 cm (photo. P. Marciniuk and M. Wolanin, 2024).

Distribution and habitat
The species is known only from the Scandinavian Peninsula in Denmark, Finland, Norway, and Sweden (Kirschner et al., 2007). Taraxacum arrhenii was found in Ukraine at one anthropogenic site. Several dozens of individuals of this species grew in the immediate vicinity of the main avenue of the Lychakiv Cemetery in Lviv. This facility is visited by numerous tourists worldwide, and it is possible that T. arrhenii seeds were brought by Scandinavian tourists. T. arrhenii is a new, probably alien, species in the flora of Ukraine.
Taraxacum baeckiiforme Sahlin (section Taraxacum, Asteraceae)
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
New record
UKRAINE: Wola, dry grassland with Orchis morio, N 49º32'13"/ E 24º01'14'’, May 10, 2017, J. Marciniuk (WSRP).
Taxonomic notes
Taraxacum baeckiiforme is a medium-sized plant distinguished by the following characteristics: leaves dark green, side lobes recurved, deltoid, medium acute, undivided, their upper edge is straight, usually entire, sometimes with robust teeth, the lower edge is straight and usually entire, interlobes acute angled with a tar-like discoloration, the terminal lobe is blunt to acute, spear-shaped, the petioles are intensely red, unwinged or very narrowly winged, midrib green not striatulate; outer bracts quite narrow, 2–3 mm wide and 12–13 mm long, horizontally positioned, unbordered; capitulum with a diameter of 35–40 mm, convex; stigmas discolored; pollen present (Figure 21D).
Distribution and habitat
This species is known to occur in the Czech Republic, Denmark, and Poland (Kirschner et al., 2007). In western Ukraine, we found one locality of Taraxacum baeckiiforme, where the species grew in a semi-natural meadow with Orchis morio. Taraxacum baeckiiforme is a new native species of Ukrainian flora.
Taraxacum crassum H. Øllg. & Trávn. (section Taraxacum, Asteraceae)
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
New records
UKRAINE: Lviv, prospekt Chervonoi Kalyny, lawn, N 49º46'53" / E 24º03'33", May 11, 2017, M. Wolanin (herb. M. Wolanin); Wola, District Mykolaiv, hill with Orchis morio, May 10, 2017, J. Marciniuk (WSRP); Ivano-Frankove (Janiw), meadow, Roztochia Biosphere Reserve, May 11, 2017, J. & P. Marciniuk (WSRP).
Taxonomic notes
Taraxacum crassum is a medium-to large-sized plant distinguished by the following characteristics: leaves light green, side lobes recurved, deltoid, medium acute, undivided; upper edge straight or irregular with entire teeth; lower edge straight or irregular, entire or subulate dentate, interlobes broadly rounded to angular without tar-like discoloration, terminal lobe not larger than other lobes, obtuse to acute, without tip, petioles on outer leaves greenish, inner leaves reddish or pink, narrowly to broadly winged, midrib green to faintly brownish; outer bracts 4–5 mm wide and 15–18 mm long, regularly arranged, usually unbordered; capitulum with a diameter 45–60 mm, convex; stigmas discolored; pollen present (Figure 21C).
Distribution and habitat
This species is known from Austria, Bulgaria, the Czech Republic, Denmark, Germany, Poland, Slovakia, and Sweden (Kirschner et al., 2007). In western Ukraine, we found this species in Lviv, where we observed several dozen individuals in lawns, as well as in Wola and Janiw, where T. crassum was observed in large numbers in dry meadows. T. crassum is a native species of Ukrainian flora.
Taraxacum diastematicum Markl. (section Taraxacum, Asteraceae)
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
New records
UKRAINE: Lviv, Lychakiv Cemetery, May 8, 2017, J. & P. Marciniuk (WSRP); Krechiv, wet meadow near the monastery, May 11, 2017, J. & P. Marciniuk (WSRP).
Taxonomic notes
Taraxacum diastematicum belongs to the T. tanyphyllum group. The main diagnostic characters of this taxon are: large plants with long, medium-green leaves, side lobes bent deltoid with a tendency to divide, their upper edge is sickle-shaped, usually entire, the lower edge is straight or more or less concave, usually entire, interlobes long serrated, sometimes with a tar-like discoloration, the terminal lobe is blunt, hastate, petioles pale on the outside, red on the inside, narrowly winged, red midrib with a striatulate pattern; outer bracts large, 4.5–5.5 mm wide and 14–15 mm long, horizontally positioned, unbordered; capitulum with a diameter of ca. 50 mm, convex; stigmas discolored; pollen present (Figure 21B).
Distribution and habitat
This species is found in the Czech Republic, Denmark, Finland, Germany, Great Britain, Poland, Slovakia, Sweden, and Switzerland (Kirschner et al., 2007). We found two localities for this species in western Ukraine. Taraxacum diastematicum occurs in semi-natural and anthropogenic grass communities: meadows, lawns, parks, and roadsides. To our knowledge, T. diastematicum is a new native species of the flora of Ukraine.
Taraxacum gibberum Markl. (section Taraxacum, Asteraceae)
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
New records
UKRAINE: Wola, dry grassland with Orchis morio, N 49º32'13" / E 24º01'14", May 10, 2017, J. Marciniuk (WSRP); Lviv, Navariyska Street – the southern suburbs of Lviv, May 10, 2017, J. Marciniuk (WSRP).
Taxonomic notes
Taraxacum gibberum is a medium-sized plant distinguished by the following characteristics: leaves mid-green, side lobes recurved, deltoid, medium acute, often divided, upper edge straight or falcate, usually entire, sometimes with robust teeth; lower edge straight, usually entire, interlobes broadly rounded, non-tar-like discoloration, terminal lobe triangular, sometimes elongated, petioles green to white, narrowly winged, midrib pallid, colored towards apex, usually not striatulate; outer bracts 3–4 mm wide and 12–13 mm long, curved reflexed or horizontally positioned, unbordered; capitulum with a diameter ca. 45 mm, convex; stigmas discolored; pollen present (Figure 21E).
Distribution and habitat
This species is found in the Czech Republic, Denmark, Finland, Germany, Poland, and Sweden (Kirschner et al., 2007). In western Ukraine, we found this species in semi-natural meadows in town Wola, and on lawns in Lviv. Taraxacum gibberum is a new native species of Ukrainian flora.
Taraxacum glossodon Sonck & H. Øllg. (section Taraxacum, Asteraceae)
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
New records
UKRAINE: Lviv, prospekt Chervonoi Kalyny, lawn, N 49º46'53" / E 24º03'33", May 10, 2017, M. Wolanin (herb. M. Wolanin); Lelechiwka, Bila Skelya, May 11, 2017, J. & P. Marciniuk (WSRP); Between Lozina and Dubrovica, wet meadow, May 11, 2017, J. & P. Marciniuk (WSRP); Fraga, Ukraine, calcareous roadside, N 49º28'03" / E 24º26'48", May 12, 2017, K. Oklejewicz, M. Wolanin (herb. M. Wolanin).
Taxonomic notes
Taraxacum glossodon is a medium-sized plant distinguished by the following characteristics: leaves olive-greenish green, side lobes patent, narrowly deltoid, medium acute, usually long, and lingulate at the apex; the upper edge is usually falcate and irregularly dentate; the lower edge is usually entire or subulate dentate, interlobes regularly plicate, tar-like discoloration absent, terminal lobe not larger than other lobes, trilobate, often with lingulate tip, petioles reddish or pink, unwinged, midrib green or faintly colored; outer bracts 4–5 mm wide and 15–17 mm long, regularly arranged, usually unbordered; capitulum with diameter 40–50 mm, convex; stigmas discolored; pollen present (Figure 22A).
Figure 22
Herbarium specimen of Taraxacum glossodon (A), between Lozina and Dubrovica 11.05.2017; T. glowackii (B), Lviv Lychakiv Cemetery 8.05.2017; T. guttigestans (C), Lviv Lychakiv Cemetery 8.05.2017; T. homoschistum (D), Lviv Lychakiv Cemetery 8.05.2017; T. hemicyclum (E), Lviv Lychakiv Cemetery 8.05.2017. Scale bar 5 cm (photo. P. Marciniuk and M. Wolanin, 2024).

Distribution and habitat
This species is found in the Czech Republic, Denmark, Finland, Germany, Norway, Poland, Slovakia, and Sweden (Kirschner et al., 2007). In western Ukraine, we found Taraxacum glossodon in Lviv, Lelechiwka, and Fraga, where we observed this species in grassy anthropogenic habitats, and between Lozina and Dubrovica, where we observed several dozen individuals in wet meadows. T. glossodon is a new native species for the flora of Ukraine.
Taraxacum glowackii H. Øllg. (section Taraxacum, Asteraceae)
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
New records
UKRAINE: Lviv, Lychakiv Cemetery, lawns at cemetery avenues N 49º49'56" / E 24º03'11", May 8, 2017, J. & P. Marciniuk (WSRP); Lviv, Piekarska Street, Lawn, May 8, 2017, J. & P. Marciniuk (WSRP).
Taxonomic notes
Taraxacum glowackii is a quite large plant distinguished by the following characteristics: leaves light green, side lobes patent, triangular to recurved, medium acute, usually undivided, upper edge usually convex, often strongly irregularly serrated, lower edge straight or concave, usually entire, interlobes clearly angular without tar-like discoloration; terminal lobe quite variable: obtuse, acute, sometimes with distinct tip, petioles outer pallid inside pink, narrowly winged, sometimes broadly winged, midrib green not striatulate; outer bracts 3–4 mm wide and 12–13 mm long, curved reflexed, unbordered; capitulum with a diameter ca. 40 mm, convex; stigmas discolored; pollen present (Figure 22B).
Distribution and habitat
This species is known only in Poland (Kirschner et al., 2007); in the eastern part of the country, it is a common species in various types of anthropogenic grassland habitats. Taraxacum glowackii was found in Lviv in western Ukraine at two sites: the Lychakiv Cemetery and a lawn on Piotrowska Street. T. glowackii is a new, probably native species for the flora of Ukraine.
Taraxacum guttigestans Kirschner & Štĕpánek (section Taraxacum, Asteraceae)
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
New records
UKRAINE: Lviv, Lychakiv Cemetery, May 11, 2017, J. & P. Marciniuk (WSRP); Wola, District Mykolaiv, hill with Orchis morio, May 10, 2017, J. Marciniuk (WSRP).
Taxonomic notes
Taraxacum guttigestans is a medium-sized plant distinguished by the following characteristics: leaves yellowish green, side lobes recurved, deltoid, medium acute, undivided, upper edge usually entire or with a large tooth at the base; lower edge usually entire or rarely subulates dentate, interlobes long, plicate, denticulate, tar-like discoloration absent or present, terminal lobe not larger than other lobes, without or with lingulate tip and usually with symmetric incisions, petioles reddish or pink, usually unwinged, midrib reddish/brownish; outer bracts 3.5–3.9 mm wide and 12–13 mm long, irregularly twisted, purplish, usually unbordered; capitulum with a diameter 45–55 mm, convex; stigmas greenish; pollen present (Figure 22C).
Distribution and habitat
Taraxacum guttigestans is a species with a Central European range, known from the Czech Republic, Denmark, Germany, Poland, and Slovakia (Kirschner et al., 2007). In western Ukraine, we found T. guttigestans in the Lychakiv Cemetery in Lviv, where we observed several dozen individuals in grassy anthropogenic habitats, and in Wola, where we observed several individuals in the dry meadow. T. guttigestans is a new native species for the flora of Ukraine.
Taraxacum hemicyclum G. E. Haglund (section Taraxacum, Asteraceae)
Synonyms: Taraxacum adiantifrons Wisskirchen & Haeupler
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
New record
UKRAINE: Lviv, Lychakiv Cemetery, lawns at cemetery avenues, N 49º49'56" / E 24º03'11", May 8, 2017, J. & P. Marciniuk (WSRP).
Taxonomic notes
Taraxacum hemicyclum is a medium-sized plant distinguished by the following characteristics: leaves mid-green, side lobes recurved, deltoid, quite acute, usually undivided, upper edge straight, entire or with robust teeth, lower edge straight or convex, usually entire, interlobes acute angled without tar-like discoloration, terminal lobe large, usually divided, with distinct tip, petioles red, narrowly winged, midrib green not striatulate; outer bracts 3.5–4.5 mm wide and 15–17 mm long, curved reflexed or straight vertical hanging, unbordered; capitulum with a diameter ca. 50 mm, convex; stigmas discolored; pollen present (Figure 22E).
Distribution and habitat
The species has a broad European range, and it is found in Austria, the Czech Republic, Denmark, Finland, Germany, the Netherlands, Poland, Russia (N European Russia, NW European Russia), and Sweden (Kirschner et al., 2007). In western Ukraine, we found this species in the Lychakiv Cemetery in Lviv, where we observed several dozen individuals along cemetery alleys in grassy anthropogenic habitats. Taraxacum hemicyclum is a new native species of Ukrainian flora.
Taraxacum homoschistum H. Øllg. (section Taraxacum, Asteraceae)
Synonyms: Taraxacum hastatum Markl.
Contributors: Jolanta Marciniuk, Paweł Marciniuk, Mateusz Wolanin
New records
UKRAINE: Lviv, High Castle, urban lawn, N 49º50'44" / E 24º02'06", May 9, 2017, J. & P. Marciniuk (WSRP); Lviv, Lychakiv Cemetery, lawns at cemetery avenues, N 49º49'56" / E 24º03'11", May 8, 2017, J. & P. Marciniuk (WSRP); Fraga, roadside (on limestone soil), N 49º28'04" / E 24º26'48", May 12, 2017, K. Oklejewicz, M. Wolanin (herb. M. Wolanin).
Taxonomic notes
Taraxacum homoschistum is a quite large plant distinguished by the following characters: leaves light green, side lobes patent, triangular, medium acute, usually undivided, their upper edge is usually falcate, entire or with robust teeth, the lower edge is straight, usually entire, interlobes broadly rounded without a tar-like discoloration, the terminal lobe is triangular, obtuse to acute, the petioles are green to white, narrowly winged, midrib green not striatulate; outer bracts, 3.5–4.5 mm wide and 14–15 mm long, curved reflexed, unbordered; capitulum with a diameter of 50–55 mm, convex; stigmas discolored; pollen present (Figure 22D).
Distribution and habitat
The species has a wide European range, and it is found in Belgium, Luxembourg, Czech Republic, Denmark, Finland, France, Monaco, Germany, Great Britain, the Netherlands, Norway, Poland, Russia (NW European part), Sweden, and Switzerland (Kirschner et al., 2007). In western Ukraine, this species was found in grassy anthropogenic habitats such as lawns, squares, wastelands, and roadsides. Taraxacum homoschistum is a new native species of Ukrainian flora.
Tragus racemosus (L.) All. (Poaceae)
Contributors: Marcin Nobis, Agnieszka Nobis, Artur Pliszko
New records
Poland: Jasionka near Rzeszów, along the road S19, (very abundantly on the roadside), 50°7'31.81"N / 22°4'51.60"E, ATPOL grid FF55, August 8, 2023, M. Nobis, A. Nobis s.n. (KRA 634279, 634280-634286), September 12, 2024, M. Nobis obs.; Sokołów Małopolski near Rzeszów, along road S19 (very abundantly on the roadside), ATPOL grid FF45, August 8, 2023, M. Nobis, A. Nobis (KRA 634287, 634288, 634291), September 12, 2024, M. Nobis obs.; Podgórze/Pikuły near Rzeszów, along the road S19 (roadside – few individuals), ATPOL grid FF25, September 12, 2024, M. Nobis obs.; Kraków, between Katowicka Street and Wizjonerów Street, 50°05.409'N, 19°53.020'E – roadside verges and pavements along the roads 79 and E77, over 1000 individuals, elev. 213 m, ATPOL grid DF69, October 15, 2023, A. Pliszko s.n. (KRA 627429-31).
Taxonomic notes
The genus Tragus Haller belongs to the family Poaceae and is represented by seven annual or perennial cespitose species (Anton, 1981; POWO, 2024). Tragus racemosus is an annual plant with 10–30 cm tall culms. The inflorescence is 2.5–6 cm long, compactly cylindrical, congested with racemelets consisting of (1)2–4 fertile and (1)2 sterile, 3.2–4 mm long spikelets (Sulekic & Zapater, 2001). It can be distinguished from other species of the genus by its 7-nerved upper glume and clustered arrangement of spikes (Anton, 1981).
Distribution and habitats
The native range of T. racemosus includes the Mediterranean area, tropical and southern Africa, and southwestern Asia, while as an anthropophyte, its occurrence was confirmed in North America, South America, Central Europe, and Central and Eastern Asia. It is usually found as a weed in pastures and along roadsides (Anton, 1981; Lazkov & Sennikov, 2014; Dudáš et al., 2018).
The first locality of T. racemosus in Poland was recorded in 2005 in Sosnowiec, where it grew on a heap of sand near one of the hypermarkets (Urbisz & Węgrzynek, 2007). The second locality was found in 2022 in Kraków on Zakopianka Street (Pliszko and Górecki in Wójcik et al., 2023), and was confirmed in 2024 (A. Pliszko, obs.). Here, we list the next localities of T. racemosus in Poland, at which it was observed in great abundance in 2023 and 2024 (from dozens to hundreds of thousands of individuals). The presence of such abundant populations indicates that this species is established in Polish flora; therefore, its status should be changed from ephemerophyte to epecophyte (neophyte occurring in anthropogenic habitats). At the site near Sokołów Małopolski (Figure 23), T. racemosus grew with plant species that are currently spreading rapidly along expressways in Poland, e.g. Digitaria ischaemum, Dittrichia graveolens, Eragrostis albensis, Leontodon saxatilis, Plantago coronopus, Spergularia salina, and Symphyotrichum ciliatum. Finding further populations of T. racemosus along roadsides in Poland is highly desirable.

BRYOPHYTES
Orthotrichum pamiricum Plášek & Sawicki
Contributors: Vítězslav Plášek, Marcin Nobis
New records
UZBEKISTAN: SE part of the country, Sariosiyo District, 30 km NW of Sariosiyo town, valley of Sangardak River, between Sangardak village and Sangardak waterfall Monument, bark of Salix sp., 38°32'24"N & 67°33'40"E, April 24, 2024, V. Plášek (OSTR #8310); E part of the country, Qashqadaryo District, Shahrisabz town, city centre, bark of Morus alba, 39°02'59"N / 66°49'39"E, April 25, 2024, M. Nobis (KRA).
Taxonomic notes
Orthotrichum Hedw. is one of the most distinctive moss genera and is easily recognized by its immersion in short-exserted capsules, which are typically ribbed, though occasionally smooth, and covered by large, campanulate-mitrate calyptraes that are usually hairy. The entire capsule is enclosed by these calyptraes. Even in the absence of sporophytes, plants can be identified by their orthotropic, sympodially branched stems along with small, isodiametric, and mostly papillose laminal cells with thickened walls. These xerophytic adaptations are closely linked to the ecological preferences of Orthotrichum species, which generally thrive in dry and sunny environments. They are often found growing as epiphytes on the bark of deciduous trees and shrubs, or epipetrically on rocks, boulders, and stone walls.
Recent taxonomic and molecular studies have confirmed the polyphyly of the traditionally conceived genus Orthotrichum, leading to the segregation of three distinct genera: Nyholmiella Holmen & E. Warncke, Pulvigera Plášek, Sawicki & Ochyra, and Lewinskya F. Lara, Garilleti & Goffinet (Plášek et al., 2015, 2016; Lara et al., 2016; Sawicki et al., 2009, 2010, 2017). The epiphytic moss Orthotrichum pamiricum was recently described as a new species in the Pamir region of Tajikistan and Afghanistan (Plášek et al., 2014). Orthotrichum pamiricum shares similarities with O. crenulatum in gametophyte characteristics, including an obtuse and crenulate leaf apex. However, when fertile, O. pamiricum can be readily distinguished by its 16 long endostome segments. Additionally, a prostome is present in O. pamiricum but absent in O. crenulatum (Figure 24).
Figure 24
Macro- and micro-photographs of Orthotrichum pamiricum. (A) plants with a young capsule covered by a naked calyptra; (B) mature capsules with a peristome; (C) stem leaves with gemmae; (D) gemmae in detail; (E) leaf sections. Scale bars: 1 mm (A–C); 100 µm (D); 250 µm (E) (photo. were taken from the specimen OSTR #8310 by V. Plášek, 2024).

Superficially, Orthotrichum pamiricum could be mistaken for Asian populations of Nyholmiella obtusifolia because of their shared ovate-obtuse leaves. However, these two species differ primarily in their sporophyte characteristics, particularly in the position of the stomata and the structure of the peristome.
Distribution and habitats
Nine species of the broadly interpreted genus Orthotrichum have been recorded in Uzbekistan: Lewinskya affinis (Schrad. ex Brid.) F.Lara, Garilleti & Goffinet, L. speciosa (Nees) F.Lara, Garilleti & Goffinet, Nyholmiella obtusifolia (Brid.) Holmen & E. Warncke, Orthotrichum anomalum Hedw., O. crenulatum Mitt., O. cupulatum Brid., O. hallii Sull. & Lesq., O. pallens Bruch ex Brid., and O. tenellum Bruch ex Brid. (Lazarenko, 1938; Lewinsky, 1992; Higuchi, 2005; Ignatov et al., 2006; Jalov, 2016; Ellis et al., 2017; Zhalov, 2018; Czernyadjeva et al., 2019; Zhalov & Abdirasulov, 2022). However, the extent to which these records are supported by herbarium specimens remains unclear. Therefore, validating this list will require critical revision of the collections in the future.
In addition to Tajikistan and Afghanistan (Plášek et al., 2014), Orthotrichum pamiricum has been recorded in Kazakhstan (Ellis et al., 2018) and Xinjiang Province in China (Ellis et al., 2016; Plášek et al., 2021). Phytogeographically, it belongs to the Asian Temperate and Warm Temperate elements, specifically within the Sino-Central Asian subelement (Plášek et al., 2021). This unit comprises species that have been exclusively documented in the mountains of Central Asia and China. Orthotrichum pamiricum is closely related to O. crenulatum, which is a species commonly found in Central Asia. In addition to Uzbekistan, O. crenulatum has been reported in Afghanistan, Kazakhstan (Lewinsky, 1992), Tajikistan (Ellis et al., 2011), Kyrgyzstan (Ellis et al., 2014), Pakistan (Schäfer-Verwimp & Gruber, 2002), and China (Skoupá et al., 2017; Plášek et al., 2021). Both species are epiphytic mosses and can co-occur, as observed in locality No. 1 (Sangardak River Valley), where both were found to grow on the same tree.
Supplementary material
The following supplementary material is available for this article:
Figure S1. Herbarium specimens of Erigeron canadensis in Mongolia.
Figure S2. Paspalum dilatatum in Dushanbe, city center, Tajikistan (photo. A. Nowak, 2024).
Acknowledgements
Research of Beata Paszko were supported by the W. Szafer Institute of Botany of the Polish Academy of Sciences whereas her visits to herbaria in Edinburgh (E), London (K), and Paris (P) were supported by the SYNTHESYS Project financed by the European Community Research Infrastructure Action under the FP7 “Capacities” Program; Zbigniew Szeląg is grateful to Dr. Richard Jürgen Koch (Osnabrück University) for discussion and confirming the plants identification and to Dr. Tim Rich (Cardiff) for valuable suggestions and improving the English text; research of Julian Chmiel on Dysphania pumilio was part of a project of Adam Mickiewicz University in Poznań, Excellence Initiative – Research University (grant no. 141/04/UAM/0005); research conducted by Marcin Nobis were supported by the National Science Center, Poland (project no. 2023/51/B/NZ8/01179); Flora of Uzbekistan Project (FZ-20200929321) was supported by Innovative Development Agency of Uzbekistan; research conducted by Serik Kubentayev has been funded by the Science Committee of the Ministry of Science and Higher Education of the Republic of Kazakhstan (grant no. AP19680161); research conducted by Ewelina Klichowska were supported by the National Science Center, Poland (project no. 2020/39/D/NZ8/02307); research conducted by Marta Krzempek were supported by project U1U/W18/NO/28.76; the research conducted by Hyeok Jae Choi was supported by the National Research Foundation of Korea (Grant No. RS-2022-NR068406); special thanks go to Mrs. Cordula Bernert, who found and posted photographs of Orobanche clausonis on iNaturalist, the taxon that was noted for the first time in France. We are also grateful to anonymous reviewer and M. Dudáš for the substential improvements to the previous version of the manuscript.
