. Introduction
Secondary metabolism was first conceptualized by Kossel in 1891 (Kossel, 1891). Unlike primary metabolites, which are essential for plant growth and development, secondary metabolites protect plants from biotic and abiotic stresses. Most secondary metabolites belong to three major types: phenolics, alkaloids, and terpenoids (Rehab & Amira, 2018). Terpenoids are the most diverse type, with at least 80,000 members, and provide abundant resources for both plant sustainability and human utilization (Christianson, 2017).
Terpenoids play a crucial role in ensuring plant survival in nature. First, they can assist plants in defending themselves against herbivores and pathogens (Gershenzon & Dudareva, 2007). For example, limonene and menthol can prevent herbivores from feeding. Azadirachtin from the seeds of Azadirachta indica could disrupt the release of morphogenic peptide hormones in Colorado potato beetles to impede the growth of pests (Kuhar et al., 2022). Terpenoids facilitate communication between plants and other organisms. They can attract insects for pollination or seed dispersal to ensure plant reproduction (Dobson, 1993). Third, terpenoids can influence population structure and species distribution through allelopathy. For example, camphor can inhibit the germination and growth of other plants. Terpenoids are essential for plant growth and development. Gibberellins, sterols, chlorophyll, and ubiquinone (Coenzyme Q) are well-known bioactive molecules. Specifically, gibberellins are phytohormones; sterols are related to the cell membrane; chlorophyll is involved in photosynthesis; and ubiquinone plays a role in oxidation resistance (Hernández-Camacho et al., 2018).
Terpenoids are also beneficial to human life. Essential oils, volatile aromatic substances extracted from plants, are commonly used as flavors and fragrances in the food and cosmetic industries. Several terpenoids have been successfully developed into clinical drugs owing to their antimicrobial, anti-inflammatory, antioxidant, anticancer, and analgesic effects. For example, artemisinin from sweet wormwood (Artemisia annua) is not only an effective anti-malarial medicine, but also exhibits anticancer and anti-inflammatory activities (Wright, 2005). Paclitaxel from Chinese yew (Taxus chinensis) is used to treat ovarian and breast cancers (Sabzehzari et al., 2020). Ginsenosides from ginseng (Panax ginseng) are applied in relieving fatigue, improving memory and the treatment of cardiovascular and cerebrovascular diseases (Ratan et al., 2021). Terpenoids are used as food colorants and nutritional health components. For example, lycopene is not only a natural red colorant but also exhibits antioxidant activity, which is beneficial to human health (Khan et al., 2021). In addition, saponins as the glycosylated derivatives of steroids are widely used as natural emulsifiers, foaming agents, and stabilizers, and rubber as the polymer of terpenoids is used in industries such as ink, coatings and plastics (Timilsena et al., 2023). Despite the importance of terpenoids in plants and humans, their content is usually extremely low. For example, artemisinin is only 0.01% to 0.6% of the dry weight of A. annua, which is far from meeting the market demand. Therefore, enhancing the terpenoid content is urgently needed.
With the development of synthetic biology, large-scale production of some terpenoids has been achieved in heterologous hosts, with artemisinin as the first landmark. Keasling’s lab spent nearly a decade achieving a high-level semi-synthesis of artemisinin. Ceaseless modifications have been made to produce artemisinin in microbes, including switching the host from E. coli to yeast (Saccharomyces cerevisiae), introducing the mevalonic acid (MVA) pathway to boost the precursor, complementing three other enzymes (cytochrome b5, CYB5; alcohol dehydrogenase 1, ADH1; and aldehyde dehydrogenase 1, ALDH1), employing a two-phase fermentation process, and adding the extractive solvent isopropyl myristate to the culture medium. Finally, the titer of artemisinic acid reached 25 g/L in 2013, and the industrial production of artemisinin was achieved in collaboration with Amyris (Paddon et al., 2013; Zhao et al., 2022). In addition to microbial hosts, tobacco is also a promising host for terpenoid synthesis. Osburn’s lab first sequenced the genome of the Chilean soapbark tree (Quillaja saponaria) and elucidated enzymes in the Quillaja saponin pathway (QS pathway). They successfully purified all the intermediates at the milligram level and synthesized the final product saponin adjuvant Q7 via transient expression in tobacco (Nicotiana benthamiana) (Reed et al., 2023). Recently, they achieved heterologous synthesis of QS-21 in tobacco (Martin et al., 2024). However, the heterologous synthesis of terpenoids relies on the comprehensive elucidation of the entire metabolic pathway. In addition, tedious synthetic steps, aberrant enzymes, incorrect compound structures, and drug resistance of pure terpenoids still hinder the heterologous biosynthesis of terpenoids. Therefore, engineering the synthesis of terpenoids in host plants remains one of the most effective ways to address the limitations of low yields.
Three major approaches to enhance the content of endogenous terpenoids in plants include overexpression of key synthetic enzymes, blocking of competitive pathways, and strengthening the regulation of synthesis. By overexpressing genes encoding key enzymes such as amorpha-4,11-diene synthase (ADS) and cytochrome P450 monooxygenase (CYP71AV1) or by inhibiting the expression of enzymes in its rival pathways, such as squalene synthase and β-caryophyllene synthase, the level of artemisinin has been improved to different extents (Lv et al., 2016). Transcription factors (TFs) regulate terpenoid synthesis by activating or inhibiting the expression of genes involved in terpenoid biosynthetic pathways. Furthermore, they usually regulate multiple enzymes and pathways simultaneously. Recently, many studies on the transcriptional regulation of terpenoids in economic medicinal plants like A. annua, tomato (Lycopersicon esculentum), cotton (Gossypium hirsutum) and danshen (Salvia miltiorrhiza) have been reported. Here, we summarize the progress in the regulation of terpenoid biosynthesis to provide references for boosting the yield of terpenoids in native plants.
. Classification and biosynthesis pathways of terpenoids
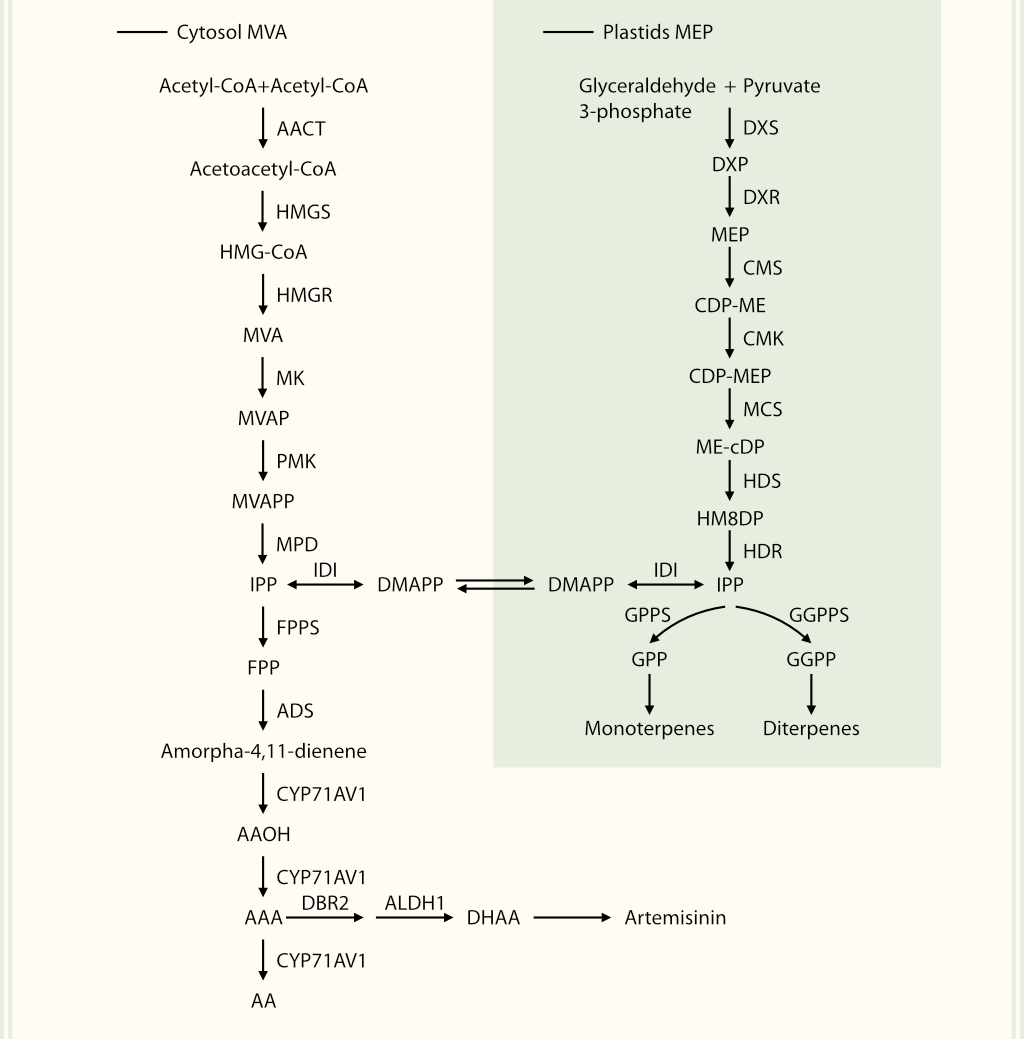
Terpenoids can be classified into monoterpenes, sesquiterpenes, diterpenes, triterpenes, tetraterpenes, and polyterpenes depending on the number of building blocks (Nagegowda & Gupta, 2020). Isopentenyl diphosphate (IPP) and its isomer dimethylallyl diphosphate (DMAPP) are two types of terpenoid building blocks. They are synthesized by the cytoplasmic MVA pathway or the plastidic methylerythritol phosphate (MEP) pathway. The MVA pathway exists mainly in animals, plants, fungi, and some protists, whereas the MEP pathway exists in bacteria, algae, and plants. Thus, plants can utilize both MVA and MEP pathways to synthesize terpenoids (Lipko et al., 2023). The MVA pathway begins with the condensation of two acetyl-CoAs to form acetoacetyl-CoA, which is catalyzed by acetyl-CoA acetyltransferase (AACT) (Miziorko, 2011) and subsequently catalyzed by hydroxymethylglutaryl-CoA synthase (HMGS), hydroxymethylglutaryl-CoA reductase (HMGR), mevalonate kinase (MK), phosphomevalonate kinase (PMK), and mevalonate pyrophosphate decarboxylase (MPD), leading to the production of isopentenyl pyrophosphate (IPP) (Vranová et al., 2013). The MEP pathway is initiated when pyruvate and glyceraldehyde 3-phosphate are catalyzed by 1-deoxyxylulose-5-phosphate synthase (DXS) to form 1-deoxy-D-xylulose-5-phosphate (DXP). DXP is further converted into IPP via catalysis by enzymes such as 1-deoxy-D-xylulose 5-phosphate reductoisomerase (DXR), 4-diphosphocytidyl-2-C-methyl-D-erythritol synthase (CMS), 4-diphosphocytidyl-2-C-methyl-D-erythritol kinase (CMK), 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase (MCS), 4-hydroxy-3-methylbut-2-enyl diphosphate synthase (HDS), and 4-hydroxy-3-methylbut-2-enyl diphosphate reductase (HDR) (Banerjee & Sharkey, 2014). Finally, isopentenyl diphosphate isomerase (IDI) converts IPP and DMAPP to form five-carbon terpenoid building blocks (Vranová et al., 2013).
IPP and DMAPP have been used to synthesize the precursors of various terpenoids. Geranyl diphosphate (GPP), the precursor of monoterpenes, is formed by one IPP and one DMAPP under the catalysis of geranyl diphosphate synthase (GPPS). Farnesyl diphosphate synthase (FPPS) catalyzes the condensation of two IPPs and one DMAPP to form the sesquiterpene precursor, farnesyl diphosphate (FPP). Geranylgeranyl diphosphate (GGPP), a precursor of diterpenes, is synthesized by geranylgeranyl diphosphate synthase (GGPPS) with three IPPs and one DMAPP (McGarvey & Croteau, 1995; Withers & Keasling, 2007). However, the precursor of triterpenes, squalene, is condensed directly from two FPPs, whereas the precursors of tetriterpenes and polyterpenes follow a similar process. These precursors are then converted into numerous terpenes via catalysis by diverse terpene synthases (TPS).
Artemisinin is a typical sesquiterpene lactone with an endoperoxide bridge, which has been the priority anti-malarial component isolated from A. annua in China over 40 years ago (Tu, 1981). FPP is first cyclized to amorpha-4,11-diene by amorpha-4,11-diene synthase (ADS) (Zhao et al., 2022). Amorpha-4,11-diene is sequentially oxidized into artemisinic alcohol (AAOH), artemisinic aldehyde (AAA), and artemisinic acid (AA) under the catalysis of CYP71AV1. AAA is catalyzed by a double-bond reductase (DBR2) and then by ALDH1 to yield the intermediate dihydroartemisinic acid (DHAA) (Judd et al., 2023), which is speculated to yield artemisinin via a photo-oxidative reaction. Here, artemisinin was used as a prime case for the transcriptional regulation of terpenoid biosynthesis.
Figure 1
Terpenoid biosynthesis pathways with artemisinin as an example HMG-CoA, 3-hydroxy-3-methylglutaryl-CoA; MVAP, mevalonate-5-pyrophosphate; MVAPP, mevalonate-5-diphosphate; MEP, 2-C-methyl-D-erythritol 4-phosphate; CDP-ME, 4-(cytidine 5'-diphospho)-2-C-methyl-D-erythritol; CDP-MEP, 2-phospho-4-(cytidine 5'-diphospho)-2-C-methyl-D-erythritol; ME-cDP, 2-C-methyl-D-erythritol-2,4-cyclodiphosphate; HMBDP, hydroxymethylbutenyl diphosphate; abbreviations of enzymes are shown in the main text.
. Transcription factors regulate the expressions of enzyme genes of terpenoid biosynthesis
The synthesis and accumulation of terpenoids are spatiotemporally regulated by a complex network in response to various biotic and abiotic stresses in which TFs play a pivotal role. TFs bind to the cis-elements in the promoter regions of their targets either independently or by forming complexes with other TFs to regulate the transcription of enzymes and ultimately the terpenoid content. They can regulate multiple enzymes in single or different terpenoid biosynthesis pathways, making them invaluable tools for plant genetic engineering. TFs are categorized into two primary types according to their functions: general TFs, which form the pre-initiation complex with RNA polymerase II, and specific transcription factors, which bind to the unique cis-elements in the promoter with their DNA-binding domain (DBD). TFs can also be sorted according to their conserved DBD into v-myb avian myeloblastosis viral oncogene homolog (MYB), WRKY (named because of the presence of WRKYGQK amino acids), basic leucine zipper (bZIP), basic helix-loop-helix (bHLH), and APETALA2/Ethylene Response Factor (AP2/ERF) families, etc. In accordance with their transcriptional regulatory roles, most TFs contain nuclear localization signals (NLS) that enter the nucleus. However, a subset of TFs that lack NLSs must interact with other TFs for nuclear translocation and are known as co-regulators. Here, we summarize the regulatory roles of TFs in terpenoid biosynthesis following the DBD classification criterion.
. MYB transcription factors participate in terpenoid biosynthesis
MYB TFs, which are prevalent in both plant and animal kingdoms, play a pivotal role in plant development, cell morphogenesis, and metabolic regulation (Cao et al., 2020). The N-terminal of MYB TFs harbors a conserved MYB domain comprising 1 to 4 R repeats, while their C-terminal exhibits variability and encompasses the transcription regulatory region. Each R repeat contains 50–53 amino acids in total and is interspersed with a tryptophan residue every 18–19 amino acids to maintain structural stability. R repeats form three α-helices, and the second and third ones coalesce into a helix-turn-helix (HTH) structure to bind to specific DNA sequences in the target genes (Thakur & Vasudev, 2022). Depending on the number of R repeats, MYB transcription factors are classified into four families: 1R, 2R, 3R, and 4R (Lal et al., 2022). The 1R (R1/R2/R3-MYB) family, which possesses a single R1/R2/R3 repeat, regulates gene transcription and chromosome structure (Feller et al., 2011). The most abundant 2R (R2R3-MYB) family, endowed with R2 and R3 repeats, plays a crucial role in the regulation of cell differentiation and secondary metabolism (Cao et al., 2020). 3R (R1R2R3-MYB), which includes R1, R2, and R3 repeats, predominantly influences cell cycle and differentiation to enhance plant resistance to stress. The smallest 4R-MYB containing four R1/R2 repeats remains largely unexplored (Dubos et al., 2010). R2R3-MYB TFs have been widely studied and have been shown to play a central role in terpenoid biosynthesis. For example, overexpression of SmMYB98 in the hairy roots of S. miltiorrhiza increased the content of tanshinones threefold (Hao et al., 2020). Furthermore, AaMYB108 significantly elevated the transcription of CYP71AV1, augmenting artemisinin production by 70–90% (Liu et al., 2023b). R2R3-MYB TFs also regulate the synthesis of paclitaxel, linalool, apocarotenoids, and ocimene in Chinese yew (T. chinensis), sweet osmanthus (Osmanthus fragrans), saffron (Crocus sativus), and snapdragon (Antirrhinum majus) (Yu et al., 2020; Bhat et al., 2021; Han et al., 2022; Lan et al., 2023). Additionally, a limited number of R3-MYB TFs have been reported to be involved in terpenoid biosynthesis. Specifically, TcMYB8 substantially enhanced pyrethrin synthesis in pyrethrum (Tanacetum cinerariifolium) by binding to the promoters of TcCHS and TcGLIP (Zhou et al., 2022).
. WRKY transcription factors modulate terpenoid biosynthesis
WRKY TFs are plant-specific and play indispensable roles in responses to biotic and abiotic stresses (Wang et al., 2023). They comprise a highly conserved WRKY domain in their N-terminal and a zinc-finger motif at the C-terminal. WRKY domain is composed of 41 amino acids and is featured by the distinctive “WRKYGQK” motif responsible for DNA binding (Mirza et al., 2022). WRKY TFs can be classified into three groups based on the number of WRKY domains and the type of zinc finger motif (Wang et al., 2023). Group I possesses two WRKY domains and one C2-H2 zinc finger. One member of this group, LcWRKY17 in laurel (Litsea cubeba), doubled the production of monoterpenes including α-pinene, camphene, and β-myrcene by specifically binding to the W box of the LcTPS42 promoter (Gao et al., 2023). Group II includes one single WRKY domain and one C2-H2 zinc finger and is the most extensively studied group among WRKY TFs. During the maturation of grape (Vitis vinifera), VviWRKY40 is downregulated by abscisic acid, which leads to the release of monoterpenol β-D-glucosyltransferases (VviGT14) and the accumulation of glycosylated monoterpenoids (Li et al., 2020). Group III contains one single WRKY domain and one C2-HC zinc finger and has only been detected in higher plants so far. CmWRKY41 in chrysanthemum (Chrysanthemum morifolium) augmented the content of sesquiterpenes by modulating the expression of CmHMGR2 and CmFPPS2 (Hu et al., 2023). Likewise, AaWRKY9 enhanced artemisinin production 1.6 to 2.2 times by regulating the expression of the AaDBR2 and AaGSW1 (Fu et al., 2021).
. bZIP transcription factors regulate terpenoid biosynthesis
bZIP TFs in eukaryotes play pivotal roles in the regulation of growth and development, signal transduction, stress responses, and secondary metabolism. This family is distinguished by the 60–80 amino acid leucine zipper domain (bZIP) (Han et al., 2023). The N-terminal region of the bZIP domain is crucial for nuclear localization and DNA-binding. The C-terminal is the leucine zipper region, which is responsible for dimerization and is characterized by scattered leucines every seven amino acids (Zhao et al., 2021). bZIP is a common TFs that regulates terpenoid synthesis. For example, SmbZIP1 negatively regulates tanshinone biosynthesis by repressing SmGGPPS expression in S. miltiorrhiza (Deng et al., 2020). Conversely, CsbZIP44 promoted the accumulation of total carotenoids by about one-third by adjusting the expression of carotenoid catabolism-related genes in citrus (Citrus spp.) (Sun et al., 2024). Similarly, TcbZIP60 directly binds to the promoters of chrysanthemol synthase (TcCHS) and allene oxide cyclase (TcAOC) genes to activate their expression and increases the total amount of pyrethrins by 1.24 times in pyrethrum (T. cinerariifolium) (Xu et al., 2023). In A. annua, AabZIP9 significantly elevated the biosynthesis of artemisinin by upregulating the ADS expression as well (Shen et al., 2019). bZIP transcription factors coordinate terpenoid biosynthesis.
. bHLH transcription factors coordinate terpenoid biosynthesis
bHLH TFs represent a substantial category of eukaryotic proteins that are widespread in both animal and plant kingdoms and play pivotal roles in growth and development, light signal transduction, and stress responses. bHLH TFs are characterized by a highly conserved bHLH domain approximately 60 amino acids in length. The bHLH domain is composed of two adjoining conserved motifs: a basic region that engages in binding to target genes, and a helix-loop-helix region that consists of two alpha helices and is responsible for dimerization (Hao et al., 2021). The roles of MYC homology box-containing TFs in the bHLH family have been extensively studied in terpenoid metabolism. Heterologous expression of lavender (Lavandula angustifolia) LaMYC4 in tobacco elevated the biosynthesis of caryophyllenes by 0.5 to 1.0 times through enhancing the expression of HMGR, FPPS, DXS, DXR, and GPPS in MVA and MEP pathways (Dong et al., 2022). TaMYC2 interacts with the promoter region of squalene synthase (TaSS) and activates its expression, leading to the accumulation of taraxasterol and taraxerol in the dandelion (Taraxacum antungense) (Liu et al., 2023c). Other bHLH TFs are involved in the regulation of terpenoids in addition to the MYC type. Overexpression of bHLH iridoid synthesis 1 (BIS1) TF in periwinkle (Catharanthus roseus) suspension cell cultures significantly increased the expression levels of iridoid biosynthetic genes and promoted the accumulation of loganic acid (Van Moerkercke et al., 2015). Overexpression of AabHLH113 boosted the contents of dihydroartemisinic acid and artemisinin by 1.7–2 times and 1.5–2.2 times respectively, by targeting the DBR2 and ALDH1 genes (Yuan et al., 2023). AP2/ERF transcription factors orchestrate terpenoid biosynthesis.
. AP2/ERF transcription factors orchestrate terpenoid biosynthesis
AP2/ERF TFs are plant-specific and are involved in morphogenesis, metabolic regulation, and hormone signal transduction. They possess a conserved AP2 domain consisting of approximately 60 amino acids and form three β-sheets and one α-helix (Meraj et al., 2020). They can be categorized into four subgroups according to the number and structure of the AP2 domains. The AP2 subgroup harbors two AP2 domains, whereas the ERF and DREB subgroups contain only one AP2 domain. The RAV subgroup is classified as a single AP2 domain coupled with a B3 domain (Licausi et al., 2010). AP2/ERF TFs are involved in the regulation of terpenoid synthesis. In Apple (Malus domestica), MdAP2-34 directly targets phytoene synthase 2 (MdPSY2) to activate its transcription, leading to 60% accumulation of total carotenoids in the fruit (Dang et al., 2021). Recent studies have found that in P. ginseng, PgERF120 positively regulates the biosynthesis of ginsenosides and enhances their content by approximately 1.8 times in transgenic hairy roots (Jiang et al., 2024). LcERF19 binds to and activates the transcription of LcTPS42, and the contents of geranial and neral increase by 242% and 266%, respectively, in L. cubeba (Wang et al., 2022a). In A. annua, AaERF1 and AaERF2, two homologous TFs, both promoted the biosynthesis of artemisinin by upregulating the transcription of ADS and CYP71AV1 (Yu et al., 2012).
. Other transcription factors in terpenoid biosynthesis
In addition to the aforementioned TF families, many other types of TFs are also involved in the regulation of terpenoid biosynthesis. For example, in P. ginseng, PgNAC41-2 of the NAC TF family increased the total saponins by 0.4 times in transgenic hairy roots (Liu et al., 2023a). CsMADS5, a member of the MADS TF family, increased the content of carotenoids fivefold in orange (Citrus sinensis) by controlling the transcription of phytoene synthase (PSY), phytoene desaturase (PDS), and lycopene β-cyclase 1 (LCYb1) (Lu et al., 2021). The scarecrow-like family TF, SlSCL3, can enhance the content of volatile terpenoids, such as β-ocimene, β-(E)-caryophyllene, and α-humulene, by two to three times through transient overexpression of SlSCL3 (Yang et al., 2021). In addition, AaSHI1 from the plant-specific SHORT INTERNODES (SHI) family positively regulates artemisinin synthesis by upregulating the transcription of ADS and CYP71AV1 in A. annua (Yang et al., 2024).
. Transcription factor complexes adjust terpenoid biosynthesis
TFs can regulate terpenoid synthesis not only individually, but also by forming complexes with other TFs. TgbHLH95 and TgbZIP44, which belong to the bHLH and bZIP families, respectively, collaboratively promote terpene biosynthesis in Chinese torreya (Torreya grandis) nuts by activating TgGPPS expression (Zhang et al., 2023). Similarly, PpERF5 and PpERF7, both from the AP2/ERF family, form a complex to strengthen the activation of lipoxygenase (PpLOX4) transcription, thus boosting volatile synthesis in peach (Prunus persica) (Wang et al., 2022b). Moreover, TFs can influence plant secondary metabolism through a ‘cascade reaction’. As demonstrated in A. annua, AaTGA6 from the bZIP family interacts with PATHOGENESIS-RELATED GENES 1 (AaNPR1), an important regulator of plant immunity in the salicylic acid signaling pathway, to stimulate the transcription of AaERF1, which then acts on artemisinin biosynthesis (Lv et al., 2016).
Table 1
Representative transcription factors involved in plant terpenoid biosynthesis
| Category | Name | Source | Function | References |
|---|---|---|---|---|
| MYB | AaMYB108 | annua | Activate the transcription of CYP71AV1 to increase the amount of artemisinin. | Liu et al., 2023b |
| SmMYB98 | S. miltiorrhiza | Transactivate the promoters of SmGGPPS1, SmPAL1, and SmRAS1 to enhance the biosynthesis of tanshinones. | Hao et al., 2020 | |
| TcMYB8 | T. cinerariifolium | Bind to the TcCHS and TcGLIP promoter, activate their expression, and promote the synthesis of pyrethrin. | Zhou et al., 2022 | |
| WRKY | AaWRKY9 | annua | Upregulating the expression of AaDBR2 and AaGSW1 positively increases the accumulation of artemisinin. | Fu et al., 2021 |
| CmWRKY41 | C. morifolium | Upregulate the expression of CmHMGR2 and CmFPPS2 to enhance the biosynthesis of sesquiterpenes. | Hu et al., 2023 | |
| LcWRKY17 | L. cubeba | Combine the LcTPS42 promoter and activate its transcription, thereby promoting monoterpene synthesis. | Gao et al., 2023 | |
| VviWRKY40 | V. vinifera | Inhibit the expression of VviGT14 to negatively regulate the accumulation of glycosylated monoterpenoids. | Li et al., 2020 | |
| bZIP | AabZIP9 | annua | Promote ADS gene expression levels, thereby increasing artemisinin biosynthesis. | Shen et al., 2019 |
| AaTGA6 | annua | Interact with AaNPR1, activate the transcription of AaERF1, and increase the synthesis of artemisinin. | Lv et al., 2019 | |
| CsbZIP44 | Citrus spp. | Regulate the expression of catalytic-related genes of carotenoids and promote the accumulation of carotenoids. | Sun et al., 2024 | |
| SmbZIP1 | S. miltiorrhiza | Down-regulate the expression of GGPPS, thereby reducing the accumulation of tanshinone. | Deng et al., 2020 | |
| TcbZIP60 | T. cinerariifolium | Activate the transcription of TcCHS and TcAOC to increase the amount of pyrethrins. | Xu et al., 2023 | |
| bHLH | AabHLH113 | annua | Enhance the expression of DBR2 and ALDH1, thereby increasing artemisinin production. | Yuan et al., 2023 |
| BIS3 | C. roseus | Increase the expression level of iridoid biosynthetic genes and promote the accumulation of loganic acid. | Van Moerkercke et al., 2015 | |
| LaMYC4 | Lavender | Enhance the expression of genes such as HMGR and FPPS to promote the biosynthesis of terpenoids such as caryophyllene. | Dong et al., 2022 | |
| TaMYC2 | T. antungense | Promote the expression of the TaSS gene, increase the accumulation of taraxasterol and taraxerol. | Liu et al., 2023c | |
| AP2/ERF | AaERF1 | annua | Upregulate ADS and CYP71AV1 transcription levels and enhance artemisinin synthesis. | Yu et al., 2012 |
| AaERF2 | annua | Upregulate ADS and CYP71AV1 transcription levels and enhance artemisinin synthesis. | Yu et al., 2012 | |
| LcERF19 | L. cubeba | Activate the promoter of LcTPS42, thereby promoting the production of geranial and neral. | Wang et al., 2022a | |
| MdAP2-34 | M. domestica | Activate the transcription of MdPSY2-1, leading to increased accumulation of phytoene and total carotenoids in fruits. | Dang et al., 2021 | |
| PgERF120 | P. ginseng | Positively regulate the biosynthesis of ginsenosides. | Jiang et al., 2024 | |
| Other | AaSHI1 | annua | Upregulate the transcription level of artemisinin biosynthetic genes and positively regulate artemisinin synthesis. | Yang et al., 2024 |
| CsMADS5 | C. sinensis | Upregulate the expression levels of PSY, PDS, and LCYb1 to promote carotenoids biosynthesis. | Lu et al., 2021 | |
| PgNAC41-2 | P. ginseng | Increase the content of total saponins. | Liu et al., 2023a | |
| SlSCL3 | S. lycopersicum | Promote the accumulation of β-ocimene, β-(E)-caryophyllene and α-humulene. | Yang et al., 2021 | |
| Complex | PpERF5–PpERF7 | P. persica | Activate PpLOX4 transcription to increase the synthesis of aroma compounds in peaches. | Wang et al., 2022b |
| TgbHLH95–TgbZIP44 | T. grandis | Bind to the TgGPPS promoter, activate TgGPPS expression, and promote the biosynthesis of terpenes in nuts. | Zhang et al., 2023 |
. Discussion and prospect
Terpenoids are the largest group of plant secondary metabolites that are not only essential for plant survival, but also beneficial to human society. Terpenoid yields in plants are seriously compromised by rate-limiting enzymes and strict regulation networks. TFs are key elements in intricate regulatory networks. They can regulate the expression of single or multiple enzyme genes alone or in a complex and ultimately affect the production of terpenoids (Huang et al., 2023). Consequently, comprehensively revealing the roles of TFs in the regulatory network of terpenoid biosynthesis will pave the way for boosting terpenoid production.
The regulatory network of terpenoid metabolism is extremely intricate, and investigations into the functions of TFs in many plants are still at a nascent stage. Advances in genomics and research methods have accelerated the discovery of TFs that are involved in terpenoid metabolism (Alami et al., 2022). As a result, the regulatory mechanism of terpenoid metabolism is constantly being elucidated, positioning TF as a vital target for genetic engineering to improve terpenoid content. There are several common strategies for identifying these TFs. Phylogenetic analysis in public databases, using reported TFs as queries, is likely to identify homologous TFs with similar functions (Wang et al., 2016). Other approaches, such as gene co-expression analysis and gene cluster identification, will assist in identifying TFs involved in terpenoid synthesis. Moreover, methods of transient overexpression, virus-induced gene silencing in native plants, or heterologous expression in other hosts, such as yeast, will further help elucidate the functions of TFs in terpenoid synthesis (Zhang, 2003). However, the establishment of transgenic systems remains an insurmountable barrier to applying TFs to harness terpenoid metabolism and boost the terpenoid content in non-model plants.
In conclusion, what strategies can be employed to enhance the production of terpenoids in non-model plant species? Numerous studies have focused on increasing the artemisinin content in A. annua. In addition to employing critical TFs, other strategies, such as overexpressing key enzymes, blocking competitive pathways, increasing the number of storage organs, and applying phytohormones, can also increase the amount of terpenoids in plants either individually or in combination.
