. Introduction
The Mesozoic is a long Era in which floras all over the world developed intensively, biodiversity increased, and plants evolved significantly. The first two periods (Triassic and Jurassic) of the Mesozoic were gymnosperm-dominated and during the third (Cretaceous), angiosperms appeared. Land plant cover was rich owing to favorable climatic conditions – warm and humid. Many fossil localities are connected to coal or clay mining, and many plant remains are available owing to drilling for geology or industry.
Here, we introduce select achievements of Polish palaeobotanists. We maintain the chronological order of studies to demonstrate directions and trends in the development of methodology and selection of subjects in line with the advancement of research techniques. Polish Mesozoic paleobotany has traditions dating back to the nineteenth century when Marian Raciborski, considered the first prominent Polish palaeobotanist and outstanding botanist (Kornaś, 1986), described some Triassic and Jurassic floras, and provided a taxonomical basis for paleofloristic studies (Raciborski, 1891a, 1891b, 1892a, 1892b, 1894). This period was powerful for collecting, determining, and describing plant fossils. Besides Raciborski, at the turn of the nineteenth and twentieth centuries, two German palaeobotanists, Goeppert (e.g., Goeppert, 1836, 1846) and Gothan (1906) contributed to the knowledge of Polish floras. The wide use of microscopy allowed the investigation of not only gross morphology but also the cuticular structure of studied leaves following the development of chemical treatment for cuticle preparation suitable for fossil coalified material by Schulze (1855). The study of cuticular features highly facilitated the taxonomical determination of plant remains and their classification. Owing to the technique of that time, a microscope was also used for preparing drawings of plant organs and cuticles. Such investigations characterized European paleobotany at the end of the nineteenth and the beginning of the twentieth century. Mesozoic fossil plant research developed in the most prominent scientific centers like Germany (Goeppert, 1836, 1846; Gothan, 1906; Roemer, 1866, 1867, 1870, 1889; Schenk, 1867), France (Brongniart, 1828–1837, 1849; Saporta, 1872–1873, Schimper, 1869–1874, 1870–1872), Italy (de Zigno, 1856–1885), and Great Britain (e.g., Seward, 1911; Thomas, 1913, 1925).
The milestone in twentieth-century Polish paleobotany was Maria Reymanówna’s style of research. She investigated Jurassic macrofossils at the Institute of Botany of the Polish Academy of Sciences, Kraków, and continued research initiated by Raciborski.
Reymanówna’s great merit was the expansion and maintenance of numerous foreign contacts, particularly with Western European palaeobotanists, which was unusual at this time. She spent 6 months between 1958 and 1959 at Prof. John Walton’s lab in Glasgow and then 5 months in 1962 in Great Britain (close cooperation with professor Thomas M. Harris). She gained considerable experience visiting foreign collections (Denmark, Great Britain, Sweden, Russia, and many others) and took part in regularly organized international paleobotanical conferences in many countries in Europe and beyond. She published in English in international journals; therefore, her results gained a wide audience. Her works were characterized by a more modern approach to taxonomy than that in the nineteenth century, in the times of Raciborski, when taxonomical studies of fossil plants were often based on leaf morphology and taxa were sometimes misclassified. Thus, pinnate leaves belonging to cycadophytes or pteridosperms were incorrectly attributed to ferns. Reymanówna was open to new techniques in paleobotany and wrote short notes about using new methods such as polymers (Reymanówna & Stuchlik, 1959) and scanning electron microscopy (Reymanówna et al., 1986) in cuticle examination.
. Triassic and Jurassic (MB, GP)
. Cuticular and Anatomical Studies
The first anatomical study was conducted by Reymanówna (1960) on Cycadeoidea, a bennettitalean trunk of Cycadeoideaceae, known mostly from Cretaceous deposits. Some trunks were found in the southern part of Poland, and Reymanówna described one of them from the Western Carpathians. The trunk was silicified, which enabled sectioning and the peel-section technique. The technique used by Reymanówna was developed by Walton (Reymanówna, 1960) and consists of etching the mineral residues, leaving a thin layer of protruding plant tissues, which were stripped off with a celluloid film.
The description contains the morphology and anatomy of the trunk and sunken bisexual cones inside the trunk containing ovules and pollen grains (Figure 1A–C). Reymanówna was one of the few palaeobotanists who provided many anatomical details of Cycadeoidea trunk, which contributes to the understanding of the life strategies of these very interesting plants.
She also noted traces of insects in the vascular system, particularly in the xylem and wood (Figure 1D,E). Borings were also observed in the cones, damaged ovules, and microsporophylls. Reymanówna did not identify insects but thoroughly studied the responses of a plant. She noticed the increased secretion of the “gum” ducts and the tendency to turn the parenchyma into a non-structured secretion, as well as a tendency to produce abscission layers and abortion of attacked parts. She dismissed the possibility that insects feeding on the cones might have pollinated the ovules because attacked ovules were abortive in the very early stage of development. This eliminated the theory of insect-pollination symbiosis (at least in this case), which is an interesting issue to date.
The most remarkable of Reymanówna’s studies, and the first of its kind in Polish paleobotany, was the extensive study of leaf cuticles according to the German method, which she introduced to Poland. This method was used, among others, for the first study of the internal structure of cupules (ovule/seed-bearing organ) belonging to Caytonia (a seed fern of the Caytoniaceae family) from the Grojec clays (Reymanówna, 1970, 1973). Her reconstruction of this organ, including its anatomy and mechanism of pollination, is still being quoted today.
Figure 1
Reymanówna (1960). The stem of Cycadeoidea sp. from Przenosza in profile. (A) General view of the trunk fragment. (B) Section of trunk fragment showing periferically arranged cones. Reproduction of Plate 1, Figures 1, 2 (permission No. AP/2022/1). (C) Reymanówna (1960). Cycadeoidea sp. longitudinal section of bud; peel section. Reproduction of Plate 8, Figure 31 (permission No. AP/2022/1). (D) Reymanówna (1960). Cycadeoidea sp. destroyed part of ramenta. Ground section. (E) The same specimen as in (D); Traces of insects (?). Undamaged tissue on the left, debris scattered on the right. Reproduction of Plate 10, Figures 42, 43 (permission No. AP/2022/1).

The studied material was well preserved as compressions of detached cupules isolated from the clay (Figure 2). For the internal study, she used her own method of sectioning and maceration. She developed also her own method for observing tracheids and vascular strands of the seeds (Reymanówna, 1970). Microtome sectioning was performed in three directions of fructification – transverse and longitudinal in two directions. As a result, for the first time, several external and internal tissues were recognized, including the vascularization of the cupule and seeds. Reymanówna (1973) provided anatomical evidence that, as previously observed by Harris, small canals in pollination openings are connected with ovule micropyles. The number of canals corresponded to the number of ovules. Reymanówna also pointed out that the small diameter of the canals corresponded with the very small size of pollen grains (ca. 30 µm) produced by the male organ of Caytonanthus. According to her interpretation, the canals were also a natural obstacle for pollination with pollen grains of other gymnospermous plants, thus qualifying the type of pollination as typical for gymnosperms.
Figure 2
Reymanówna (1973); reproduction of Figure 12 with original explanation (permission No. AP/2022/1).
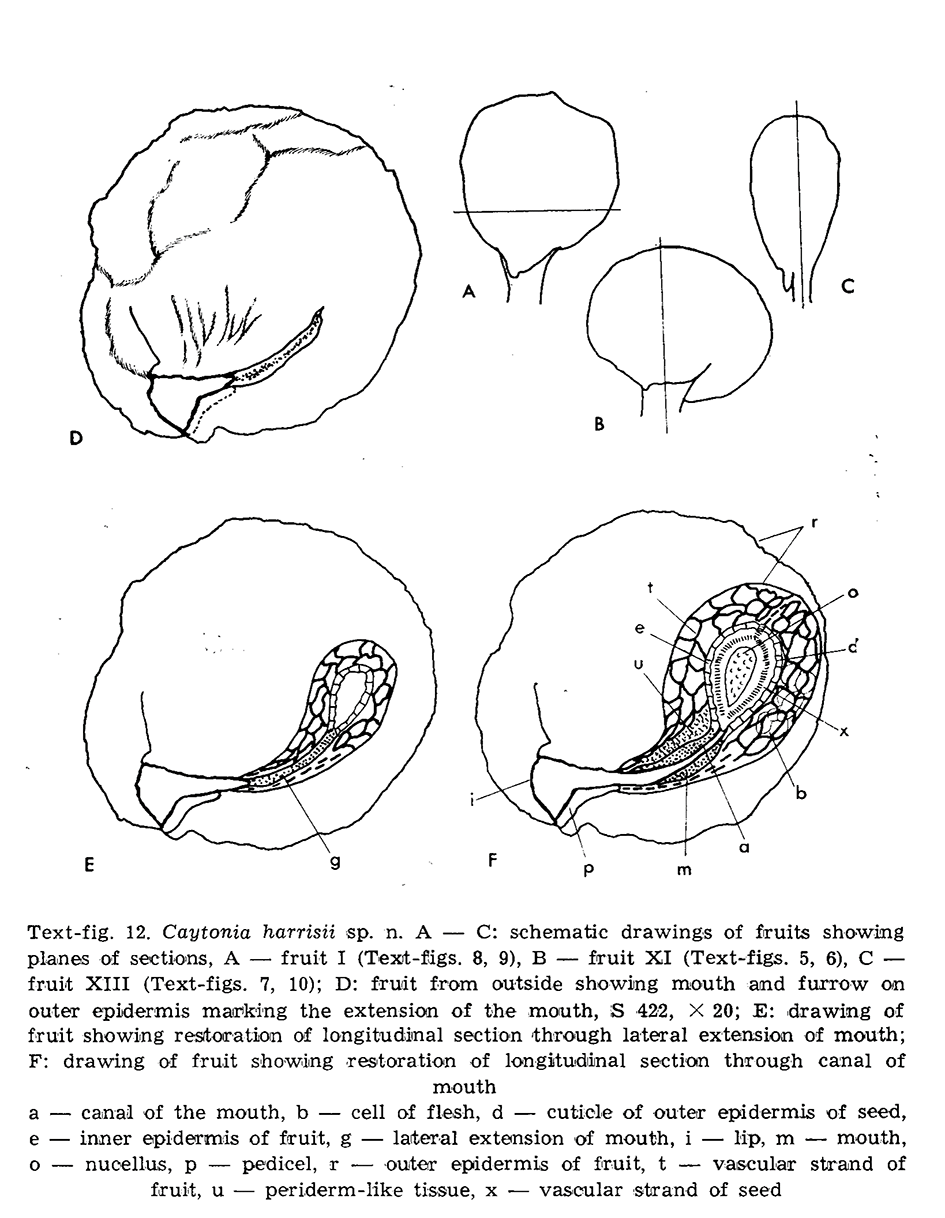
Another result of her study was anatomical proof confirming the hypothesis that the cupule of Caytonia was equivalent to an incurved megasporophyll segment similar to the leaflet of Sagenopteris (leaves attributed to Caytonia) (Reymanówna, 1974, with references).
Similarity concerned the double-cuticle structure of the cupule, net of the vascular system, and structure of the flesh of fructification and leaf mesophyll. This study on Caytonia fructification was, in the early seventies, spectacular both as an experimental methodology and as a source of important information about the reproductive strategy of this common, but still enigmatic Mesozoic seed fern. At the same time, the study was an important contribution to gymnosperm evolution, manifested by an increasing number of protected ovule-bearing structures in this advanced plant group, pollen-conducting structures, and pollination mechanisms. Observations on the possible origin of this organ were later used as a basis for further research and interpretation by other researchers (e.g., Doyle, 2006, 2008; Krassilov, 1977; Shi et al., 2021). Fossil plant anatomy has rarely been investigated but continued during the history of paleobotany. The first papers on this subject appeared at the beginning of the twentieth century (e.g., Scott, 1901; Stopes & Fuji, 1911). Recently, owing to modern techniques such as X-ray or computer tomography, anatomical studies have become more popular (e.g., De Beer et al., 2006; Blanco-Moreno et al., 2021; Friis et al., 2011).
. New Fossil Floras From Upper Triassic
New horizons in Polish Triassic research opened up in the 1990s and early twenty-first century with the discovery of localities with well-preserved plant remains associated with vertebrate bones (Pacyna, 2014). Krasiejów, Patoka, Poręba, and Lipie Śląskie-Lisowice (Figure 3) were the fossil plant sites found during the prospection and excavation of bones of large land vertebrates. Philippe et al. (2015) documented the conifer wood Agathoxylon keuperianum from several Silesian Upper Triassic localities. This species is restricted to the Germanic Basin Keuper (Poland, Germany, and France). Polish material helped clarify the taxonomy of this species. Zatoń et al. (2015) analyzed carnivore coprolites from Lipie Śląskie-Lisowice and Poręba which contained in the composition some plant cuticles. Pacyna (2019) documented rare hydrophilic elements, sphenopsids, and ferns, from the flora of Krasiejów. A sphenopsid species, Neocalamites merianii, was also identified. It was preserved as impressions, flattened casts, molds, and isolated leaves. Only one small fern specimen, Sphenopteris sp., was found. These elements were probably transported to the deposition site and did not grow where they were buried.
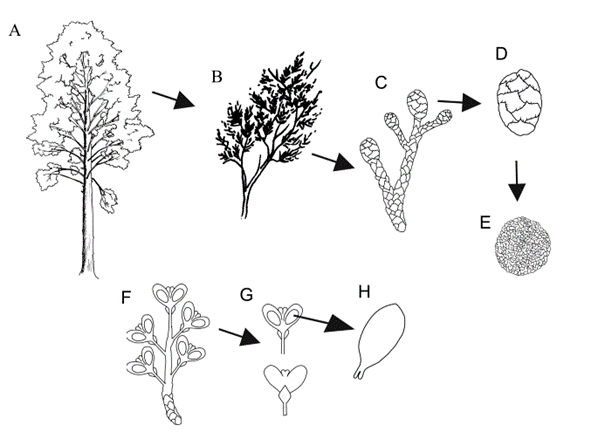
Taxonomic studies, although constitute the basis for all other paleofloristic studies, are, in most cases, underestimated. However, such fortunate records are rarely found in the Triassic clay pit in Patoka (Figure 3). Among macrofossils preserved as compressions in clay and easily removable from the sediment by hydrofluoric acid, the coniferous leafy shoots were collected together with female cones containing ovules and male cones with pollen sacs containing Enzonalasporites pollen grains known earlier from the sediments in a dispersed state (Figure 4). These pollen grains were also found inside the micropyle of the ovules. Determining the affiliation of individual organs to one plant based on organic connections, the same cuticular structure of these organs, with characteristic papillate cells and stomata also protected by papillae, justified the creation of new a genus and species – Patokaea silesiaca (Pacyna et al., 2017). The structure of the reproductive organs was outstanding, and placed Patokaea in a new family, Patokaeaceae, combining the Brachyphyllum–Pagiophyllum type of leaves with seed scale-bract complexes derived from evolutionary advanced Permian–Triassic Voltziales. The shape and size of the pollen cones resemble those of Classostrobus, the male cone of Jurassic–Cretaceous Cheirolepidiaceae, but the arrangement of the pollen sacs on the microsporophyll corresponds more closely with that of Voltziales. It seems that this family did not continue to evolve further within conifers but turned out to be important in evolutionary considerations (Atkinson et al., 2018; Leslie et al., 2018) and in research on pollen preserved in situ (for references see Pacyna et al., 2017; Zhang et al., 2021). This is the first and so far the only detailed documented fossil coniferous plant in Polish paleobotany.
Figure 3
Map of localities and regions studied. (A) The Sudetes; (B) Lower Silesia; (C) The Holy Cross Mountains. Violet – Triassic; blue – Jurassic; green – Cretaceous (drawing by A. Sojka).
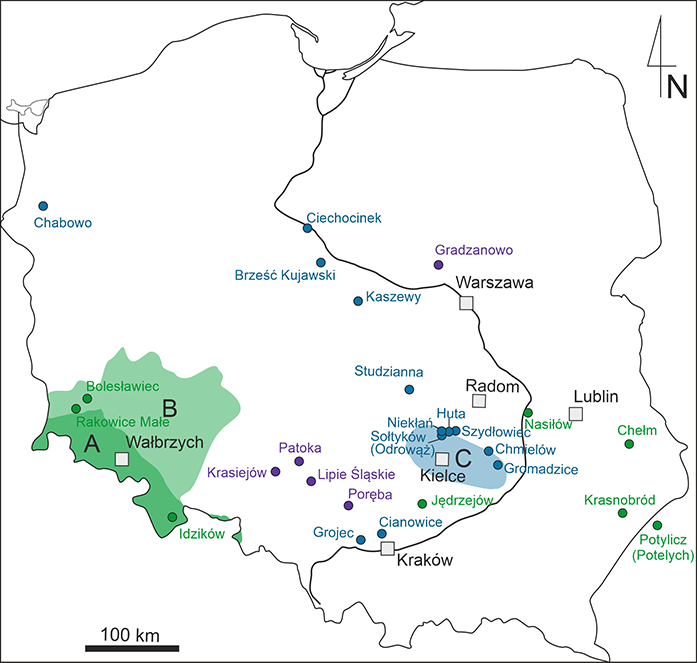
Figure 4
A reconstruction of Patokaea silesiaca Pacyna, Barbacka et Zdebska. (A) Tree. (B) Branch fragment. (C) Leafy twig with a male cone attached. (D) Male cone. (E) Pollen grain found in situ. (F) Ovuliferous cone. (G) Ovuliferous scale-bract complex. (H) Ovule.
Not to scale (drawing by A. Sojka).
. Taxonomy, Revisions, New Data on Old Materials, Investigations, and Reconstructions of Polish Jurassic Floras
Strict taxonomic studies are applied to single records, often originating from drilling cores and not associated with other taxa. Their identification allows for the confirmation of given taxa in given areas, which provides evidence of their distribution. This type of research is the most classic and has been conducted from the beginning of paleobotany to the present day. In the Polish Triassic (Rhaetian) seed fern, Lepidopteris ottonis was found in the drilling core Gradzanowo 2 near Płońsk. Its identification involved the revision of historical specimens of this species from Silesia–Kraków Highland (this species was described for the first time from Poland in the nineteenth century by Goeppert) and the Holy Cross Mountains stored at Wrocław University (Barbacka, 1991). Taxonomic revision of these specimens resulted in the discovery of the female reproductive organs of this seed fern, the cupule (Peltaspermum rotula) containing seeds, so far, the only such record in Poland.
Early Jurassic fern remains of matoniacean affinity were found in the Lower Hettangian strata of lacustrine/backswamp origin from the Niekłań PGI 1 borehole (central Poland, Holy Cross Mountains). The preserved fragments were identified as Matonia braunii (Barbacka et al., 2016). The gross morphology of the remains suggests a low-growth plant with palmately compound fronds. Spores were found in situ in well-preserved sori. This study confirmed that this species and two other similar taxa, Phlebopteris braunii and P. muensteri, were conspecific. The fern was deposited in strata indicating a warm and humid climate approaching transgression, resulting in a high water table and enhanced accumulation of organic matter.
Studies of historical Raciborski’s collection of Middle Jurassic fossil plants from Grojec Clays near Kraków were more extensive. However, it needed careful revision because, since Raciborski studied and described this flora in the nineteenth century, the taxonomic system changed significantly with the requirements of modern taxonomy. Jarzynka and Pacyna (2015) revised the sphenophytes from Grojec clays. Three new species of Equisetum created by Raciborski (Equisetum renaulti, E. remotum, and E. blandum) are attributed to the common Jurassic species Equisetites lateralis and Equisetites cf. columnaris. The occurrence of Neocalamites lehmannianus (originally described by Raciborski as Schizoneura hoerensis) has been confirmed from Grojec clays. Numerous specimens that Raciborski referred to as Neocalamites lehmannianus showed heterogeneity and served to describe the new species Neocalamites grojecensis (Jarzynka & Pacyna, 2015). Jarzynka (2016) revised the osmundaceous ferns from this locality. The number of Cladophlebis species recognized by Raciborski was much reduced. The presence of fertile leaves of Osmundopsis and Todites was confirmed and illustrated for the first time using high-quality photos. Pacyna (2014, 2021a) began a revision of the Early Jurassic flora from the Holy Cross Mountains based on the historical Raciborski and Makarewiczówna collections and new materials from the Gromadzice locality. The presence of Matonia braunii in the Hettangian of the Holy Cross Mountains was confirmed, and Phlebopteris angustiloba was documented based on numerous specimens, including sterile specimens not noted earlier in Poland (Pacyna, 2021a).
Pacyna (2021b) documented the occurrence of the fern Matonia braunii preserved in the growth position in sandstones of dune origin in the late Pliensbachian Szydłowiec Sandstone of the Drzewica Formation in the Holy Cross Mountains (Figure 5A). He concluded that the presence of this common Early Jurassic fern in such an unfavorable environment attests to its huge ecological plasticity.
Although studies on whole flora cannot be considered a strict milestone in Polish Mesozoic paleobotany, they cannot be overlooked when considering the last 100 years of paleobotanical activity. Floristic compositions of different localities and different epochs or stages contributed to understanding general biodiversity and their variability in a spatial-temporal context, depending on the local environment and climate changes. They constitute a database for all later analyses and modelling of the mechanisms of plant response to topographic and climatic factors. Paleofloristic studies are popular and have been conducted since the nineteenth century and have continued throughout the world.
After Raciborski initiated paleofloristic studies in Poland in the nineteenth century with three large localities, namely, Grojec clays, Gromadzice, and Chmielów, several localities were discovered and a large number of samples for microfossils and macrofossil specimens were collected. Nevertheless, these localities are still sites of new records and the subject of new studies.
Flora from Gromadzice, initially investigated by Raciborski (1891a, 1891b, 1892a, 1892b), was later a subject of further research by Makarewiczówna (1928), Samsonowicz (1929), Karaszewski (1960, 1962, 1965), and Grabowska et al. (1970), Wcisło-Luraniec and Barbacka (2000), Ziaja and Krupnik (2010), Pacyna and Zdebska (2011), and Pacyna (2013, 2014, 2021a). Makarewiczówna (1928) determined the age of the flora to be Early Jurassic. Two plant assemblages from different depositional environments have been reported in this locality. One was associated with a crevasse splay and was dominated by ginkgophytes (Ginkgoales and Czekanowskiales) and conifers. The other represented riparian thickets with a significant dominance of ferns, rare cycadophytes, seed ferns, sporadic ginkgophytes, and conifers.
Study of Early Jurassic (early Pliensbachian) flora from Chmielów extended after Raciborski to twentieth century (Grabowska et al., 1970; Karaszewski, 1960, 1962, 1965; Makarewiczówna, 1928; Reymanówna, 1963a, 1963b; Samsonowicz, 1929). The range of the locality and long-term research revealed some assemblages depending on local conditions. Czekanowskiales (Ginkgophyta) and non-diversified conifers (genus Podozamites) associated with cycads occurred along the riverbanks, while the drier upslope assemblage was composed of Ginkgoales, seed ferns, and bennettitaleans.
The classic, mainly taxonomical paleofloristic studies changed over time to more advanced works including palaeoecology, reconstructions of local environments or ecosystems, and investigations of climate influence on plant composition, structure, adaptation mechanisms, and life strategy. The discovery of a new locality in Odrowąż (Sołtyków) (Figure 3) in the 1970s provided new Lower Jurassic (Hettangian) plant fossils and opportunities for modern research (Karaszewski & Kopik, 1970; Reymanówna, 1987). The interest increased with the discovery of numerous dinosaur tracks (nests, eggs, and footprints) (e.g., Niedźwiedzki, 2006; Niedźwiedzki & Niedźwiedzki, 2004; Pieńkowski, 1998).
The diversified flora of this site is composed of lycopsids, sphenopsids, ferns (Figure 5B), seed ferns, cycadaleans, bennettitaleans, gnetaleans, ginkgoaleans, and conifers (e.g., Barbacka et al., 2007, 2010, 2022; Reymanówna, 1987; Wcisło-Luraniec, 1989, 1992a, 1992b; Ziaja, 1992) representing flora not very rich in taxa but fully developed vegetation with forms ranging from herbs to bushes and low trees, typical for well-watered plains with meandering rivers and periodically flooded areas (Figure 5D).
The localities studied and described during the last 20 years were discovered and explored in the twentieth century; they include Jurassic boreholes in Huta and Studzianna, Ciechocinek, Chabowo, Brześć Kujawski, Cianowice, Kaszewy, and Triassic clay-pit in Patoka (Figure 3). The first five localities have been studied together with the mutual correlation of their strata (Barbacka, Pacyna, et al., 2014). Their taxonomic research was followed by a study of floristic changes over time and changing depositional environments.
The Studzianna and Huta OP-1 boreholes (Figure 3) were also interesting in terms of plant responses to climatic changes (Barbacka, Pacyna, et al., 2014). Both localities were situated approximately 70 km from each other. There was a relatively short time difference between them. Huta represented the lowermost part of the Hettangian sequence, and Studzianna, the uppermost, almost at the Lower/Middle Hettangian boundary. There were no common elements in both floras associated with the same lacustrine facies, and the major plant groups in the two were different.
Huta contained fern–gymnosperm plant assemblage with conifers and ginkgophytes (Figure 5E), whereas Studzianna contained, apart from ferns, dominant Czekanowskiales (three genera) typical for the Siberian Region (Figure 5C) and was associated with a rather cold climate (for reference see Barbacka, Pacyna, et al., 2014). The co-occurrence of three Siberian genera in Studzianna confirmed that over a relatively short period (probably less than 1 million years, but possibly a few hundred thousand, in the later interval of the Early Hettangian and the earliest Middle Hettangian), the climate became colder and drier (confirmed by kaolinite/illite ratio; for reference see Barbacka, Pacyna, et al., 2014). Such a change in this time interval was sufficiently long for a floristic change.
Cianowice is another interesting locality (Barbacka et al., 2021) dominated by diverse and abundant bennettitaleans, whereas ferns and other gymnosperms (seed ferns, conifers, and incertae sedis) are sparse (Figure 5F,G).
Figure 5
(A) Palmately dissected frond of Matonia braunii; seen from above. Void after stipe decay and several pinnae visible. Late Pliensbachian, Podkowiński Quarry, Szydłowiec, Drzewica Formation; specimen No. PC IB UJ 109/1 (photo M. Szewczyk). (B) Fern Todites princeps from Odrowąż; specimen No. KRAM P PM 68/122. (C) Czekanowskia rigida, one of the Siberian element from Studzianna; specimen No. KRAM P PM 138/39 (photo M. Szewczyk). (D) Accumulation of coniferalean leaves of Podozamites sp., one of the most characteristic plant for Odrowąż; specimen No. KRAM P PM 68/1079 (photo M. Szewczyk). (E) Fern Cladophlebis nebbensis and incertae sedis Pseudotorellia sp.; specimen No. KRAM P PM 139/159, Huta OP-1 borehole (photo M. Szewczyk). (F) Seed fern Ptilozamites cycadea from Cianowice; specimen No. KRAM P PM, 141/1. (G) Bennettitalean Pterophyllum thomasii from Cianowice; No. KRAM P PM 141/15 (photo M. Szewczyk).

The gross morphology and cuticle structure of all bennettitaleans showed xerothermic character (small leaf size and thick cuticle with protected stomata), which together with other factors suggested a specific local paleoenvironment – a low and dry slope (Figure 6). This small locality provided good evidence for the role of local conditions in plant composition and structure (Barbacka et al., 2021).
Figure 6
Reconstruction of the environment of Cianowice based on the taxonomical composition of the assemblage, plant morphology, cuticular structure, taphonomy, and climatological and sedimentological background (modified from Barbacka et al., 2021; drawing by A. Sojka).

The use of interdisciplinary methods was helpful in detailed studies on climate–plant relationships. In the Kaszewy borehole (Upper Rhaetian–Toarcian), climatic changes were studied (Pieńkowski et al., 2020) based on carbon isotope data, together with sequence stratigraphic correlation and biostratigraphic markers. The changes (fluctuations of average temperature) were followed by alterations of the dominant plant taxa (genera of Ginkgoales and Czekanowskiales) along the sequence. Plant adaptation to temperature changes was confirmed by the stomatal index associated with atmospheric pCO2.
. Reconstructions of Jurassic Ecosystems, Interactions Between Plants and Vertebrates
Plant cover shares close relationships with animals, forming an ecosystem. The discovery of dinosaur tracks in the Holy Cross Mountains, especially in locations where plant remains were described earlier, has boosted new research on the flora there. By adding invertebrate records, full reconstruction of ecosystems is possible for many localities.
Studies on Mesozoic ecosystems require a more developed environmental background and rich fossil records. Such reconstructions, more or less precise, have became popular some decades ago. One of the earliest fully investigated ecosystems was the Middle Jurassic Yorkshire area with fossil plants and ichnological records of invertebrates and vertebrates (Harris, 1961; Harris & Miller, 1974; Romano & Whyte, 2003). An exceptionally rich record of diversified animal and plant microfossils and macrofossils was noted in the USA in the Morrison Formation (e.g., Chure et al., 2006), the Chinese Daohugou Formation (e.g., Pott et al., 2012; Pott & Jiang, 2017), and the North American Hell Creek Formation (Johnson, 1997; White et al., 1998), which have become the subject of multidisciplinary and paleoecological studies.
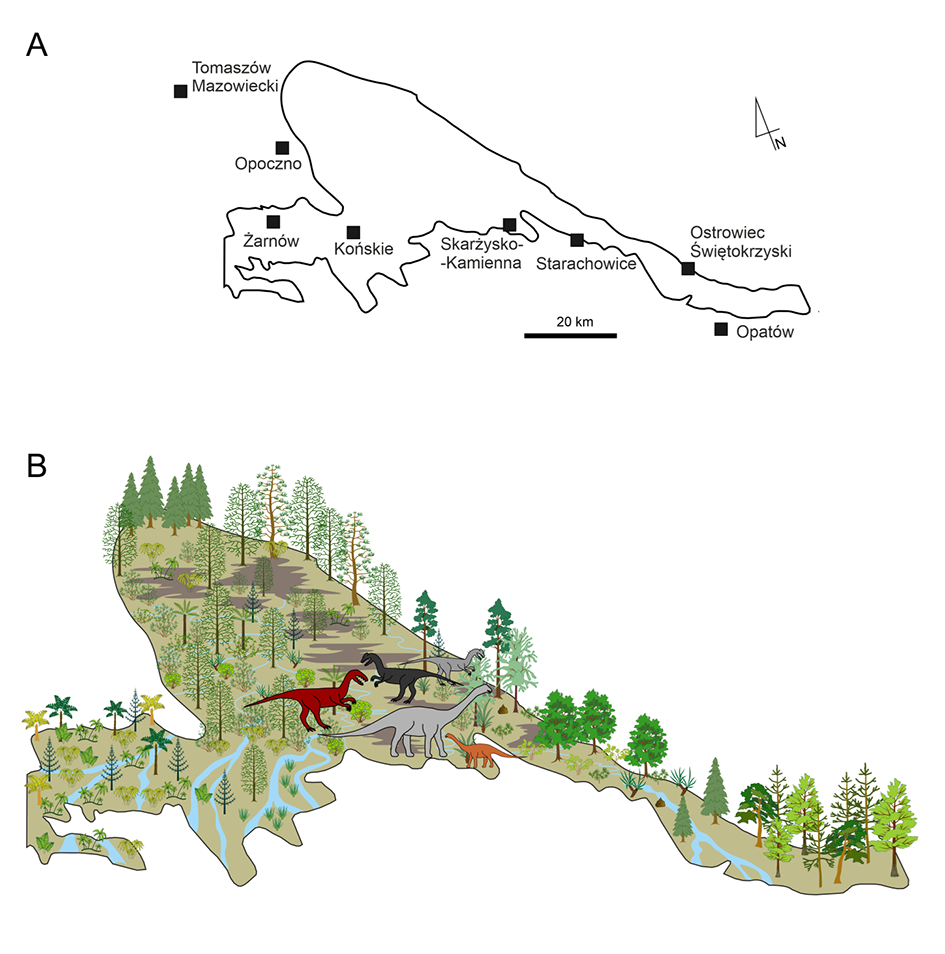
The Jurassic ecosystems in the Holy Cross Mountains are still being studied (Pacyna et al., 2022) using interdisciplinary research and methods. The sequence stratigraphy of boreholes and outcrops forms the basis for topographic maps of the study area in subsequent time horizons. Such maps are completed by plant fossils from corresponding sites and levels, and other data on vertebrate occurrence (tracks of dinosaurs, mammal-like reptiles, early mammals, lepidosaurs, pterosaurs, and early crocodylomorphs). Invertebrate remains found in coprolites (insect cuticles, bivalves, and gastropod shells) and vertebrate bones and teeth provide additional information on the ecosystem. Changes in topography (changes in riverbeds, floods, rise of crevasses, sea level fluctuations, transgressions, and regressions) are sometimes associated with changes in climatic conditions and are closely related to changes in plant composition in a given territory (Figure 7). Changes in plant cover are followed by changes in the animals living and foraging in these areas. The study of ecosystem dynamics is becoming increasingly popular worldwide and is of wide interest. Research on plant-animal interactions at different levels is promising. These include examination of drilling and foraging traces on leaves (Pacyna et al., 2018), the study of vertebrate footprints, and analysis of coprolites containing plant remains (Barbacka et al., 2022).
Figure 7
(A) Study area corresponding to the Lower Jurassic deposits in the northern margin of the Holy Cross Mountains. (B) A reconstruction of topography in the selected area (sedimentary basin of the lower part of Zagaje Formation; earliest Hettangian; parasequence Ia) with plant cover based on plant fossils and most characteristic dinosaur trackmakers based on the climatological and sedimentological background (modified from Pacyna et al., 2022; drawing by A. Sojka).
Modern methods of coprolite examination (non-invasive using computer tomography analysis) and preparation by dissolving samples by a chemical process (invasive method) have revealed, besides other remains of animal origin, plant cuticles belonging to different gymnosperms (they have a thicker cuticle and are more resistant to digestion). Seven coprolites of sauropodomorph herbivores and the same amount of those of large predators found in Sołtyków (Odrowąż, the Holy Cross Mts) were studied (Barbacka et al., 2022). Plant remains in the predator’s excrements were likely ingested together with the stomach and intestines of prey animals. The cuticles from the studied coprolites were well preserved, which allowed the taxonomic determination of most of the plants from which they originated (Figure 8).
Figure 8
Reconstruction of plants from dinosaur coprolites and their hypothetic environment based on known plant environmental requirements and climatological and sedimentological background (according to Barbacka et al., 2022; drawing by A. Sojka).
The plants consumed by dinosaurs belonged to seed ferns (including the new species proposed), cycadophytes, ginkgophytes, and conifers (13 taxa and three indeterminate cuticle types). One of the cuticle types, a fragment of a needle leaf, also represents a new taxon (new genus and species). Its triangular cross-sectional shape is very similar to that of the leaves of Pinus, and this is the oldest example of such a leaf type in the fossil records.
Most ingested plants came from beyond the immediate surroundings of today’s outcrop (Sołtyków) since they were not previously reported in this territory (Barbacka et al., 2010). Interestingly, Pachypteris papillosa was inferred to be halophytic (collected mainly in the coastal zones) in the coprolites of large predators. Increased boron content was noted in the Holy Cross Mountains in Lower Jurassic sediments near Miłków. Presumably, this confirms the local salinity of the soil, and may explain the occurrence of halophytic plant species in inland areas and suggest that large predators roamed over large areas (the distance between Miłków and Sołtyków is approximately 70 km). Palynological analysis of the coprolites yielded 31 taxa of spores and pollen grains from herbivore coprolites and 14 sporomorph taxa from predator coprolites, which provided additional data about plants growing in today’s Holy Cross Mountains.
. Statistical Methods in Paleobotany
Statistical methods have been used in paleobotany for decades since computers came into everyday use. Multivariate analyses were introduced in paleobotany in the 1970s (Spicer & Hill, 1979), and then employed for paleoecological studies (e.g., Jasper et al., 2010; Rees et al., 2000). Polish databases and statistics were used to provide an overview and comparison of all significant European Early, Middle, and Late Jurassic floras based on environmental conditions. The database for this study comprised 770 taxa from 46 localities grouped into 25 units from France, Germany, Greenland, Hungary, Italy, Norway, Poland, Romania, Scotland, Serbia, Sweden, Switzerland, and the United Kingdom (Barbacka, Bodor, et al., 2014).
To examine the extent of floras diversification and its possible causes, multivariate statistical methods were used: alpha diversity, multivariate methods (sample-centered, Q-mode, community analytical methods), principal component analysis (PCA), non-metric multidimensional scaling (NMDS), and cluster analysis. The analyses were applied at different taxonomic levels, namely, at the level of species, genus, and major plant groups, depending on the aim of the analysis. For example, cladograms based on plant genera show affinities between different localities based on environmental factors, while those based on species affinities indicate only taxonomical correlations. Grouping plants in the PCA plot and three-dimensional NMDS plot revealed that the similarity in floras was caused by local environmental and climatic conditions. Floras of the same age did not necessarily have similar composition. Habitat played a primary role in forming plant assemblages and determining their diversity for 56 million years of the Jurassic period. This grouping method was also useful for determining the most probable environments in the case of allochtonic drifted flora, where the environment could not be defined.
Another subject based on statistics was the study of plant extinctions in Poland and Europe (Barbacka et al., 2017) during events at the Triassic–Jurassic boundary (TJB). An extended database was created for 311 macroflora taxa from 106 localities (including 16 Polish localities) in 13 countries grouped into 29 regions, and 571 microflora taxa from 105 localities (29 Polish localities) in 11 countries grouped into 37 regions. The data covered the period from the Anisian to Toarcian stages. The TJB is known as one of the most significant mass extinctions that affected both marine and terrestrial ecosystems and caused the disappearance of many animal taxa, mostly in the marine realm. Within a project supported by the National Science Center, the study aimed to verify the influence of climatic changes during TJB events on terrestrial flora, which was not as evident as that on fauna. For analysis, multivariate methods such as hierarchical clustering, Horn’s coefficient of dissimilarity, and Ward’s method (for references see Barbacka et al., 2017), controlled by standardization (classical rarefaction and simple by-list rarefaction) were used. Based on these analyses, it was determined that there was no mass extinction of European terrestrial plants during the TJB. Furthermore, there was no significant difference in plant composition between the Rhaetian (the last stage of the Triassic) and Hettangian (the first stage of the Jurassic) stages suggestive of a dramatic event. In addition, for European localities, the floral changes (replacement of taxa) were not caused by global events during the TJB but rather depended on local changes in topography. The extinction of certain taxa/families by the end of the Triassic (e.g., Peltaspermales) was due to catastrophic events or a consequence of ordinary evolutionary processes remains to be understood.
. Biomolecular Studies on Fossil Woods
Paleobotany also uses biomolecular methods to determine the relationships between certain fossils. Biomolecules are organic compounds with structures identical to those of living organisms (Briggs & Summons, 2014). Such compounds are uncommon in sedimentary organic matter because they are destroyed or modified during diagenesis. Conditions required for their preservation in sediments are a low degree of maturation (temperatures not above 50–60 °) and an oxygen-depleted sedimentary environment. However, biomolecules can survive in sediments for tens or even hundreds of millions of years under favorable circumstances. For example, some pigments, such as carotenoids, melanins, and porphyrins, can survive virtually unaltered even for approximately 200 million years (Briggs & Summons, 2014). In the plant kingdom, the most common biomolecules are DNA, proteins, carbohydrates, and lipids (Li et al., 2022). Except for DNA and proteins, the remaining biomolecules can survive in sedimentary organic matter (e.g., Marynowski & Simoneit, 2022; Otto & Simoneit, 2001), and lipids appear the most stable.
Marynowski et al. (2007) found lipid biomolecules preserved in a conifer wood Protopodocarpoxylon sp. from the Middle Jurassic of Poland for the first time. Unaltered lipid compounds identified in fossil wood include ferruginol, sugiol, communic acid, lambertianic acid, and 2-ketototarol, among others. The aforementioned compounds are resin constituents, which are well known in extant trees. However, phenolic abietanes, such as ferruginol and its derivatives, are produced only by individual conifer families, including Cupressaceae s. l., Podocarpaceae, and Araucariaceae. Such a molecular composition uniquely classifies Protopodocarpoxylon into one of these groups, which is in line with the anatomical characteristics of this fossil wood (Philippe et al., 2002).
. Cretaceous (ATH)
. Palaeobotanical Research on Cretaceous Floras in Light of the Philosophy of Science
The present synthetic account of research on Mesophytic floras from Poland during the last century allows a few comments pertaining to the general methodology of science, a subdiscipline of the philosophy of science.
As described in more detail in Section 3.2, early investigators of Cretaceous floras were convinced of their significant similarity to present-day floras; the presence of present-day genera in the Cretaceous was thus considered a rule rather than an exception. This perspective was abandoned in the mid-twentieth century. Cretaceous floras were subsequently considered to be composed mostly of groups of angiosperms other than those frequent in present-day floras. As shown above, this change has had methodological consequences and resulted in a radical modification of our understanding of the history of terrestrial vegetation. Within paleobotany, such a change has been a genuine paradigm shift, a term used by the philosopher Thomas Kuhn to denote a fundamental change in the basic concepts of a scientific discipline (Kuhn, 1962).
Moreover, it is worth noting that the project of revising Cretaceous floras from Poland (see Section 3.2 for details) has been feasible only within a precise historical period. Specifically, this would not have been possible without political events having taken place. First, the near-disappearance of the internal borders of the European Union made feasible a research project conducted by an international team on the basis of collections scattered in several museums in Poland, Germany, and the Czech Republic. Similarly, working on the collections assembled by Polish scientists in the Natural History Museum of the National Academy of Sciences of Ukraine in Lviv (formerly Dzieduszycki Museum, Lwów) is no longer a political taboo. This observation emphasizes the importance of the social context of scientific discoveries. This is a counter-example to the more traditional approach of scientific research being free from all cultural biases (e.g., Beard, 2015, and references therein).
. Taxonomic Investigations of Cretaceous Leaf Floras
Three Late Cretaceous megafloral assemblages have been described from the present territory of Poland, starting at the very end of the eigtheenth century (von Buch, 1797). Due to boundary changes, plant fossils from southwestern Poland were dealt with by German and Czech palaeobotanists, whereas Polish scientists worked on the material collected partly in the territories belonging now to Ukraine. The Coniacian flora of the Sudetes was described by Velenovský (1883), Bayer (1896), Langenhan and Grundey (1891), and Frič (1897). Goeppert (1866) and Roemer (1889) investigated the Coniacian–Santonian flora in Lower Silesia. The Campanian–Maastrichtian fossil plants from the locality Potylicz (now Potelych in Ukraine) have been described by Nowak (1907).
These works were of paramount importance for the understanding of the Late Cretaceous floras of Central Europe. First, collections of plant fossils were patiently assembled over long periods (approximately 50 years for Potylicz flora). All Late Cretaceous floras from Poland are known from marine sediments, therefore, plant fossils are rare, and there are no individual beds characterized by higher frequencies of such findings (in contrast to the situation in lakes or fluvial sediments). Assembling collections was thus possible only through long-term monitoring of small quarries. It must be stressed that such collections cannot be established today as quarries are few and large, and their exploitation is conducted using heavy machinery (so fossils are almost entirely lost).
The papers published between 1797 and 1907 were mostly prepared according to the state-of-the-art methods of that period, and several species names used up to now have been validly introduced by the aforementioned authors. The practice of classifying Cretaceous angiosperm species within modern genera, such as Magnolia, Quercus, and Bignonia, widespread in the nineteenth century, was taxonomically erroneous and created a wrong impression of the close similarity between the Cretaceous and Recent floras.
After approximately a century, during which megafloral studies of mostly local significance were published (e.g., Cieśliński & Milaković, 1962; Karczmarz & Popiel, 1971), the project of a comprehensive taxonomic revision of all Late Cretaceous megafloras from Poland was formulated in 2006 by Maria Ziembińska-Tworzydło and Aleksandra Kohlman-Adamska. The general plan was presented by Halamski (2007, 2008). This revision was completed within approximately 13 years, starting from the megaflora of the Campanian–Maastrichtian of eastern Poland and western Ukraine (Halamski, 2013), through the megaflora and microflora of the Coniacian of the Sudetes (Halamski & Kvaček, 2015), and finishing with the Coniacian–Santonian megaflora, mesoflora, and microflora of Lower Silesia (Halamski et al., 2020). Halamski (2020) provided a short general overview of this project. It has been possible to attain such a goal due to a fortunate conjunction of two causes – methodological changes in paleobotany and political changes in Europe.
The methodological change in Cretaceous paleobotany consisted in starting the description of the leaf remains not by determining presumed affinities with modern taxa on the basis of a small number of characters, but rather from a very detailed description of morphology, followed by effectuating a placement within an artificial (morphographic) classification system, and “translating” (whenever possible) the form-taxa into units of the natural system only at the end. This approach was proposed by Hickey (1973, 1974), developed by Krassilov (1979), and standardized by Ellis et al. (2009). Halamski (in Halamski & Kvaček, 2015; Halamski et al., 2018) proposed an adaptation of this method to poorly preserved Cretaceous floras in marine sediments of Central Europe, which consisted in using precisely delimited but wide fossil genera.
The major results of the systematic investigations of Cretaceous megafloras in Poland include the identification of the common species Dryophyllum geinitzianum (Coniacian of the Sudetes) and Dryophyllum westerhausianum (Santonian of Lower Silesia) as representatives of the Fagales. Analogous interpretation has been conducted for the type species of the genus Dryophyllum from Eocene in the Paris Basin (Jones et al., 1988) and was extended to the Cretaceous leaves, which were shown to belong to the same genus (Halamski & Kvaček, 2015, Halamski et al., 2020).
The discovery of the oldest (Santonian) representative of the extinct group of trifoliolate platanoids is also noteworthy. The leaves (preserved solely as dispersed leaflets) from the environs of Bolesławiec in Lower Silesia were described in 2020 as the new species Platanites willigeri. Similar leaves are known from the Upper Cretaceous of western North America (Platanites marginatus; see Johnson, 1996) and the Paleocene of North America and northern Europe. Today, Platanaceae is a relict family with low diversity; all modern representatives thereof have simple leaves. The second-oldest representatives of the family Pandanaceae all over the world have been found in the Coniacian of the Sudetes (Halamski & Kvaček, 2015).
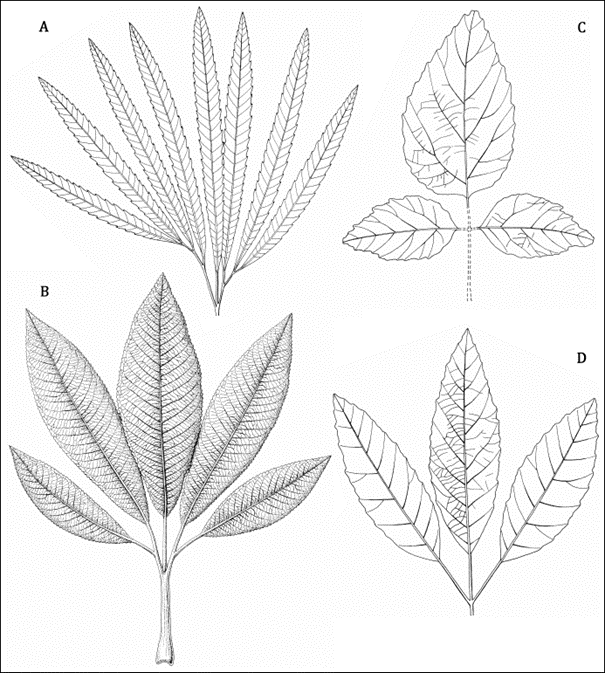
Finally, the study of Cretaceous floras allowed us to propose an important systematic criterion that is valid for both Cretaceous and Cainozoic floras. The presence of compound leaves was shown (Halamski, 2013) among dicots nearly exclusively in a subgroup thereof – the so-called eudicots or tricolpates – defined primarily based on palynological characters. This is an important criterion for interpreting angiosperm fossils (Figure 9, Figure 10).
Figure 9
Dewalquea paulinae (Halamski) Halamski (Eudicotyledoneae; possibly basal eudicots): nearly complete leaf (holotype JS3); Jan Sitek collection, Museum of the Zamość Inspectorate of the Home Army, Bondyrz, Poland. Krasnobród, upper Campanian (compare Halamski, 2013, Figure 2B and Figure 2B herein).

Figure 10
Leaves of selected Late Cretaceous plants from Poland. (A) Dewalquea haldemiana Debey ex Saporta et Marion (Eudicotyledoneae; possibly basal eudicots). (B) Dewalquea paulinae (Halamski) Halamski (Eudicotyledoneae; possibly basal eudicots). (C) Platanites willigeri Halamski et J. Kvaček (form group Platanophylls; probably related to Platanaceae). (D) Dryophyllum westerhausianum (P. B. Richt.) Halamski et J. Kvaček (Fagales). (A,C,D) Semi-schematic reconstructions after Halamski et al. (2020). (B) Reconstruction after Halamski (2013).
Not to scale. Drawings by Bogusław Waksmundzki.
. Reconstructions of Cretaceous Ecosystems
Another methodological innovation in the investigation of Cretaceous flora was the synthetic use of megaflora, mesoflora, and microflora-related data. The mesofossils are fossils that are too small for traditional megafossil-related techniques, but too large for standard micropaleontological techniques. Thus, they have long been overlooked by researchers. The first palaeobotanist to study Cretaceous mesofossils was Else-Marie Friis (e.g., Friis & Skarby, 1981; for the Cretaceous of Silesia first by Knobloch & Mai, 1986).
Using megafossils, mesofossils, and microfossils together allows for a much more detailed reconstruction of plant paleocommunities than if any single data class had been used. It has been shown that different classes of fossils, even if they occur together within the same rock, may come from different plant paleocommunities and/or ecological groups. For example, within the Turonian–Santonian Klikov flora (southern Bohemia), Laurales and Platanaceae are known as megafossils, but no representative of either group has been found among the numerous (65 species) mesofossils (Heřmanová et al., 2021). Similarly, the Late Cretaceous plant megafossil assemblage from Lower Silesia is dominated by angiosperms, and among palynomorphs, bryophyte and pteridophyte spores are dominant (Halamski et al., 2020). This phenomenon reflects the origin of the megafossil record, mostly from the forest-forming trees, whereas the microflora is primarily related to the understory or vines. Late Cretaceous forests with a fern-dominated understory were intermediate in character between the Caenophytic (modern, i.e., angiosperm-dominated) and Mesophytic (pre-angiosperm) plant communities (Halamski et al., 2020). In other words, in the Coniacian–Santonian, the Cretaceous Terrestrial Revolution (Lloyd et al., 2008) was still ongoing.
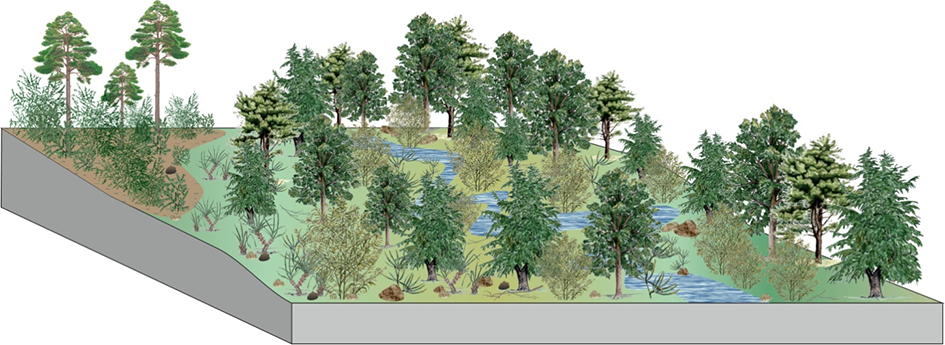
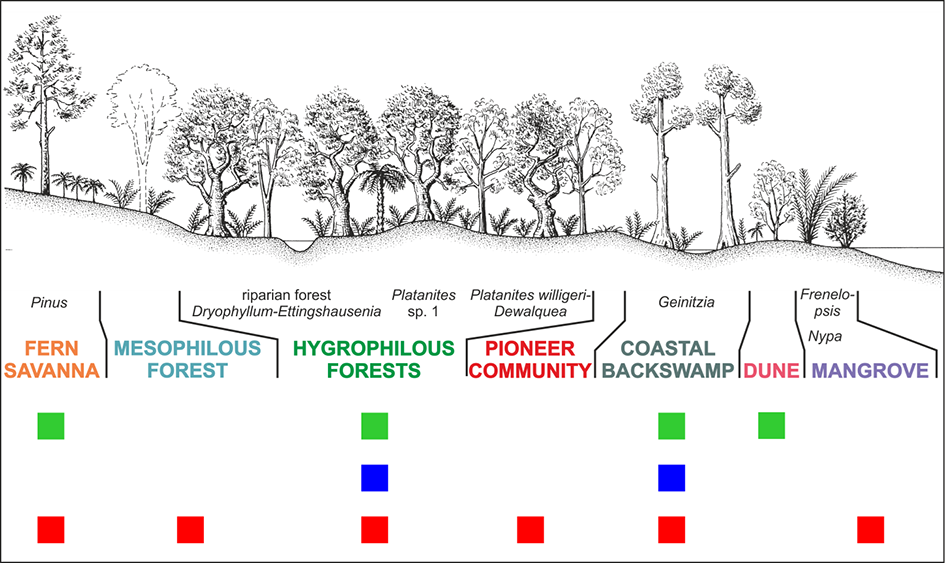
The proposed reconstruction of the Coniacian–Santonian vegetation of the West Sudetic Island (Halamski et al., 2020; Heřmanová et al., 2019, 2020) includes the following seven types of palaeocommunities: mangroves (the palm Nypa) and halophytes (the conifer Frenelopsis); dunes with Dewalquea haldemiana and possibly matoniaceous ferns; coastal backswamp conifer forests; pioneer communities; hygrophilous forests (mostly platanoid-dominated; three variants); mesophilous forests; fern savanna (Figure 11). One might also mention the oldest (Santonian) occurrence of the palm Nypa and the second-to-last (Coniacian) occurrence of caytonias (Mesozoic seed ferns; the last occurrence is from the Santonian of Sakhalin).
Figure 11
Hypothetical reconstruction (palaeocatena sensu Krassilov, 2003) of vegetal paleocommunities in the Coniacian–Santonian flora of Lower Silesia (modified after Halamski et al., 2020). Green squares – paleocommunities preserved in the megafossil record; blue squares – paleocommunities preserved in the mesofossil record; red squares – paleocommunities preserved in the microfossil record. Drawing by Bogusław Waksmundzki.
The field of science has always been in progress. It is difficult to judge the value of recent discoveries, such as those described above, and to deal with the Polish Cretaceous. However, the morphographic method of the study of poorly preserved leaf megafossils and ecosystem reconstructions based on the total evidence continue to be used on European Cretaceous floras (McLoughlin et al., 2021); therefore, at least part of the above-mentioned methodological innovations and systematic as well as paleoecological discoveries may be expected to offer a solid contribution to paleobotany.
. Summary
Mesozoic studies of Polish floras exhibit well the development of paleobotany. Owing to technical progress and numerous new discoveries, the general idea and attitude toward paleobotany are changing through an increasing scope of studies and their accuracy. On the one hand, we strive for a broad view of ecosystems from the taxonomy of individual specimens and on the other hand, we analyze fossils on the biomolecular level, in line with general scientific trends.
