. Triassic (AFM)
. Palynostratigraphy and Taxonomy
The development of palynological studies of Triassic deposits in Poland took place in the second half of the twentieth century, when palynomorphs began to be widely used as a stratigraphic tool, particularly in continental facies devoid of other fossil groups. The miospores (all fossil plant spores smaller than 200 µm, regardless of whether they are isospores, microspores, small megaspores, prepollen or pollen grains) and megaspores (large spores of heterosporous vascular plants, usually >200 µm) research covered the area of Poland, except for its south-eastern part, and thousands of samples taken from around 200 drill cores and several outcrops have been examined. This resulted in the development of a detailed palynozonation scheme, the identification of many new megaspores and some new miospore species, and reconstruction of the Polish Triassic floras.
. Miospore Zones
The palynostratigraphy of the Triassic deposits in Poland outside the Carpathians, based on miospores, was proposed and developed by Orłowska-Zwolińska (1966, 1967, 1971, 1972a, 1972b, 1977, 1982, 1983a, 1983b, 1984, 1985, 1986, 1988), who distinguished eight assemblage zones and 10 subzones, which are well correlated with other European palynological schemes (Kürschner & Herngreen, 2010). The Lundbladispora obsoleta-Protohaploxypiuns pantii comprises Induan and earliest Olenekian, Densoisporites nejburgii – Olenekian, Voltziaceaesporites heteromorphus – latest Olenekian-early Anisian, Perotrilites minor – middle Anisian, Tsugaepollenites oriens – late Anisian, Heliosaccus dimorphus – Ladinian, Porcellispora longdonensis – late Ladinian-early Carnian, Aulisporites astigmosus – middle Carnian, Corollina meyeriana – Norian, and Riccisporites tuberculaus – Rhaetian.
. Megaspore Zones
In parallel, Marcinkiewicz (1962, 1969, 1971, 1977, 1978, 1992) and Fuglewicz (1973, 1977a, 1979a, 1979b, 1980) conducted megaspore studies. Their results not only determined many new species, but also established a palynostratigraphic scheme, which included eight assemblage zones: Otynisporites eotriassicus (Eo) comprising Induan, Trileites polonicus (Po) – early Olenekian, Talchirella daciae (Da), middle Olenekian, Trileites validus (Va) – late Olenekian-early Anisian, Capillisporites germanicus (Ge) – middle Ladinian, Dijkstraisporites beutleri (Be) – late Ladinian, Narkisporites harrisi (Ha) – middle Carnian, and Trileites pinguis (Pi) – late Norian-Rhaetian, which were correlated to miospore zones (Marcinkiewicz et al., 2014).
The determination of the Early Triassic species Otynisporites eotriassicus by Fuglewicz (1973) turned out to be particularly important. This megaspore occurs in many sections across the Permian–Triassic boundary worldwide and determines the correlation horizon (Foster & Afonin, 2005).
Miospore studies of the Keuper from the Carpathian Foreland by Pautsch (1958, 1971, 1973), the Triassic of the Holy Cross Mountains by Dybová-Jachowicz and Laszko (1980), Fijałkowska (1992, 1994a, 1994b, 2006a, 2006b), and Fijałkowska-Mader (1999, 2018, 2021), the Triassic Polish Tatra Mountains by Fijałkowska and Uchman (1993), Buntsandstein from the Sudetic foreland (Fijałkowska, 1995), the Triassic of the Nida Basin by Fijałkowska-Mader (2013), the Keuper of the Upper Silesia by Fijałkowska-Mader, Heunisch, and Szulc (2015), and megaspore studies of Buntsandstein from the Holy Cross Mountains by Rdzanek (1982) should also be mentioned.
. Taxonomy
Based on palynostratigraphical studies, more than 50 megaspores (Fuglewicz, 1973, 1977b; Marcinkiewicz, 1962, 1971, 1978) and 10 new miospore species have been described, and 12 new combinations have been developed (Fijałkowska, 1994b; Orłowska-Zwolińska, 1966, 1971, 1972a, 1972b, 1983a; Pautsch, 1958).
. Reconstruction of the Polish Triassic Floras Based on Spores and Pollen Grains
After establishing the temporal sequence of changes in microflora assemblages, the next stage in the development of palynological research was the reconstruction of Polish Triassic flora. Miospores were the only tool, as the Triassic plant macrofossils of Poland are very poorly known (Pacyna, 2014). A study on the development of vegetation during the Triassic period in relation to climatic and environmental changes was conducted by A. Fijałkowska-Mader (Fijałkowska-Mader, 2015; Fijałkowska-Mader et al., 2020; Jewuła et al., 2020). An extensive analysis of the literature concerning the botanical affinity of miospores allowed us to link many Triassic dispersed spores and pollen to parent plants (Fijałkowska-Mader, 2015, Table 1). The addition of the modified method of Visscher and Van der Zwan (1981) and the Sporomorph EcoGroup (SEG) model developed by Abbink (1998) enabled the determination of the palaeoenvironmental preferences of the parent plants. The method of Visscher and Van der Zwan was originally developed for the later Triassic spore-pollen assemblages. It is based on quantitative spore-pollen analysis and classifies miospores into xerophytic and hygrophytic elements. Its modification consisted detailing and adapting it to the older Triassic spectra. In the modified method, there are three groups of elements: xerophytic (pollen grains of pteridosperms and/or conifers, excluding Illinites and Parillinites, and cricumpollen), hygrophytic (spores of lycopsids, excluding Densoisporites, horsetails, and ferns), and intermediate (Densoispoites spores, Illinites, and Parillinites, as well and cycadalean and bennettitalean pollen grains). The SEG model connects spores and pollen grains with floristic palaeocommunities, which occupy certain ecological niches: upland/hinterland (composed of pollen grains of arborescent pteridosperms), as well as arborescent and herbaceous conifers, lowland and river (consisting of spores of mosses, horsetails, ferns, pollen grains of herbaceous conifers of chierolepidiaceous affinity), as well as pollen of cycadales and bennettites, and coastal (dominated by spores of halophyte lycopsids). The analyses mentioned above were applied to miospore assemblages stated by Becker et al. (2020), Fijałkowska (1992, 1994a, 1994b, 2006a, 2006b), Fijałkowska-Mader (1999, 2013, 2018, 2021), Fijałkowska-Mader, Heunisch, and Szulc (2015), Fijałkowska and Trzepierczyńska (1990), and Orłowska-Zwolińska (1972a, 1972b, 1977, 1982, 1983a, 1983b, 1984, 1986, 1988; unpublished archival materials) in almost hundred boreholes located in different parts of Poland, with the exception of the eastern part, Carpathians, and Sudetes.
The reconstruction of the earliest Triassic flora of Poland was based on the analysis of the spore-pollen spectra of L. obsoleta-P. pantii zone: coniferous shrubs (Kürschner & Herngreen, 2010), seed ferns [documented by pollen grains Lunatisporites novaulensis, L. pellucidus, Protohaploxypinus pantii (Figure 1B), P. samilovichii, and Striatoabietites balmei], and caytonies (represented by pollen grains Vitreisporites pallidus) covered dry upland areas on the land, which existed in the early Triassic in southern Poland and was a combination of Bohemian, Silesian, and Małopolska massifs with the East European Craton (Dadlez et al., 1998; Ziegler, 1990), while lycopsids [producers of Densoisporites playfordii, Endosporites papillatus, Kraeuselisporites cuspidus, Lundbladispora obsoleta (Figure 1A), and L. willmotti spores], ferns of the Osmundaceae and Marattiaceae families (documented by Cyclotriletes microgranifer, C. triassicus, and Punctatisporites triassicus spores), and horsetails (represented by Calamospora tener spores) dominated in wet areas along the river banks and on delta plains, as well as on the coastal zone along south-eastern part of the marine Central European Basin, also called the German Basin (Abbink, 1998; Abbink et al., 2004a, 2004b). Bennettites (documented by Cycadopites coxii and C. follicularis pollen grains) are characteristic elements of the coastal ecosystems that correspond to modern mangroves (Visscher et al., 1993). The pioneer lycopsids of Pleuromeia sternbergii (producer of Densoisporites playfordii spores) grew in both dry and humid places with normal and increased salinity (Grauvogel-Stamm, 1999; Mader, 1990a).
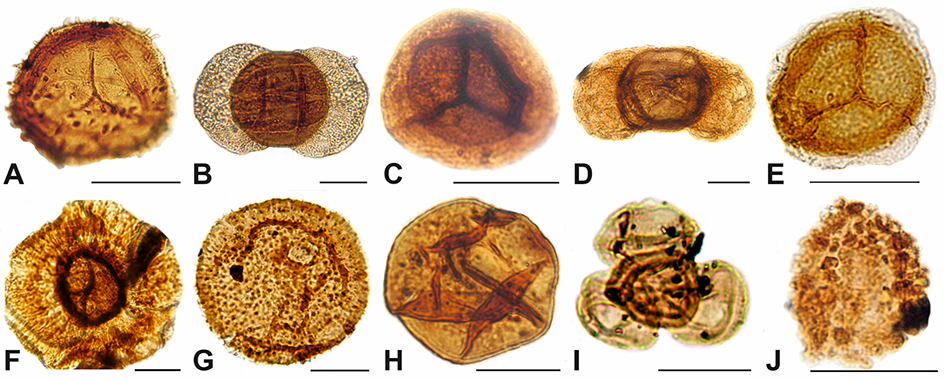
Figure 1
The index miospores of the Polish Triassic: (A) Lundbladispora obsoleta, Tumlin-Podgrodzie IG 1 borehole, depth 204.5 m, Lower Buntsandstein; (B) Protohaploxypinus pantii, Tumlin-Podgrodzie IG 1 borehole, depth 204.5 m, Lower Buntsandstein; (C) Densoisporites nejburgii, Podzamcze IG 1 borehole, depth 182.5 m, Middle Buntsandstein; (D) Voltziaceaesporites heteromorpha, Oblęgor IG 1 borehole, depth 90.9 m, Upper Buntsandstein (Röt); (E) Perotrilites minor, Opoczno PIG 2 borehole, depth 1,230.3 m, Lower Muschelkalk; (F) Heliosacuus domorphus, Opoczno PIG 2 borehole, 1,030.3 m, Lower Keuper; (G) Porcellispora longdonesis, Boża Wola IG 1 borehole, depth 1,316.0 m, Middle Keuper; (H) Aulisporites astigmosus, Mniszków IG borehole, depth 2,320.0 m, Middle Keuper; (I) Corollina/Classopllis meyeriana (tetrad), Włoszczowa IG 1 borehole, depth 1,770.0 m, Middle Keuper, (J) Riccisporites tuberculatus, Włoszczowa IG 1 borehole, depth 1,800.0 m, Upper Keuper. Scale bar: 30 µm.
In the younger miospore assemblage of the D. nejburgii zone, the domination of lycopsids is observed. Pleuromeia rossica Neuburg (represented by Densoisporites nejburgii spores (Figure 1C), P. sternbergi and others (documented by Endosporites papillatus, Kraeuselisporites cuspidus, and Lundbladispora brevicula spores), ferns of the family Osmundaceae (producers of Cyclotriletes microgranifer, C. oligogranifer, C. triassicus, Cycloverrutriletes presselensis, and Punctatisporites triassicus spores), and sphenophytes (represented by Calamospora sp. and Equisetumsporites sp. spores) inhabited wet areas of alluvial and delta plains (Abbink, 1998; Abbink et al., 2004a, 2004b; Yaroshenko, 1975, 1977).
At the Early–Middle Triassic boundary, which is correlated to the Voltziaceaesporites heteromorpha zone, a gradual decrease in the dominance of lycophytes in favor of arboresccent gymnosperms, that is, seed ferns, cycads/bennettitales, and caytoniales, can be observed. This phenomenon is global and reflects the revival of diverse plant ecosystems after the Late Permian crisis. Later, the flora was strongly dominated by xerophytic conifers (Kürschner & Herngreen, 2010). Simultaneously, the paleogeography of Poland was changed to separate the Bohemian and Małopolska massifs in the southern and East European Craton with a wide coastal zone-flood plain stretching from the vicinity of today’s Przemyśl to Koszalin (Dadlez et al., 1998). From these areas the material was transported to the German Basin, where most of the studied borehole sections were located. The land areas were overgrown by conifers, mainly Voltziaceae [documented by Voltziaceaesporites heteromorpha (Figure 1D), Angustisulcites gorpii, A. grandis, A. klausi, Brachysaccus ovalis, Klausipollenites sp., Triadispora crassa, T. plicata, and T. sp. pollen grains] and Podocarpaceae families (represented by Microcahryidites doubingeri, M. fastidiosus, M. sittleri, Platysaccus sp. div., and Alisporites sp. div. pollen grains). Taxonomically diverse conifer forests and shrubs covered not only dry land areas, but also deltas, floodplain areas, and marine coasts, where the herbaceous conifer Aethophyllum stipulare Brongniart, a producer of Illinites chitonoides pollen grains, which forms reed-like belts (Grauvogel-Stamm, 1978; Mader, 1990a; Visscher et al., 1993), coexisted with lycopsids. Drier lowland areas and river valleys were inhabited by ferns (producers of Cyclotrilets sp. and Punctatisporites triassicus spores), horsetails (with Calamospora sp. spores), and bennettitaleans (documented by Cycadopites sp. pollen grains), whereas wetter areas were inhabited by lycophytes (represented by Aratrisporites sp. div., Kreuselisporites apiculatus, and K. ullirichi spores), sphenophytes, and ferns (Mader, 1990a).
Little can be said about terrestrial flora in the early Middle Triassic (P. minor and T. oriens zones), as this area of Poland was flooded by a sea, which only withdrew at the end of the Middle Triassic, with land sedimentation coming back in the time corresponding to the upper part of the H. dimorphus zone. A warm, humid climate favors the development of taxonomically diverse plants (Ziegler et al., 1994). The whole area of Poland is comprised of land with local lakes and swamps (Dadlez et al., 1998). Voltziacean conifers producing Minutosaccus gracilis and Podosporites amicus pollen grains were found not only in the hinterland areas surrounding the south, east, and north of the alluvial plain basin, but were also pioneers of alluvial plain-coastal palaeobiocenoses (Kürschner & Herngreen, 2010; Visscher et al., 1993). A new component of these biocenoses was shrubby conifers, which produced monosaccate pollen grains of Heliosaccus dimorphus (Figure 1F). It also includes the voltzialean Aetophyllum stipulare. Lycophytes are producers of Aratrisporites sp. div. microspores, which are characterized by high environmental tolerance (Brugman et al., 1994; Grauvogel-Stamm, 1978; Orłowska-Zwolińska, 1979; Rothwell et al., 2000; Visscher et al., 1993). Numerous taxonomically diverse lycophytes (Aratrisporites sp. div. and Lycopodiacidites sp. spores), ferns (documented by Leschikisporis aduncus, Todisporites cinctus, T. minor, Deltoidospora sp. div. Acanthotriletes sp. div., and Apiculatisporites sp. div. spores), and horsetails (producers of Equisetumsporites sp. and Calamospora tener spores) inhabited wetter areas of the delta and floodplain. Sphenophytes also formed rushes along river and lake banks (Mader, 1990b, 1997).
At the Middle–Late Triassic boundary, which can be correlated to the P. longdonensis zone, xerophytic plants dominated, and an intensive development of the voltziacean conifers producing Ovalipollis and Triadispora pollen grains took place. This period was followed by the short-time transgression of a sea, rather than its gradual disappearance and transformation into sebha/playa and even saline basin in the area of today’s Greater Poland. The sedimentary basin occupies most of Poland, except for its NE, SE, and SW edges (Dadlez et al., 1998). Voltziales (represented by Labiisporites triassicus, Minutosaccus sp. div., and Parillinites sp. div. pollen grains), Podocarpaceae (producing Podosporites amicus pollen grains), and cheirolepidiacean conifers (represented by Duplicisporites granulatus and Partitisporites maljavkinae pollen grains) grew on land surrounding the sedimentary basin. The wetter areas were inhabited by ferns (producers of Echinitosporites iliacoides spores), erdmanithecaleans (documented by Eucommidites microgranulatus pollen grains), and liverwort moss producing the spores Porcellispora longdonensis (Figure 1G) (Mader, 1990b, 1997; Reinhardt & Richer, 2000). Areas with increased salinity around the playa and/or sebkha basins were colonized by the halophyte lycophyte Lepacyclotes (Annnalepis) zeilleri (producers of Aratrisporites sp. div. microspores) (Mader, 1997; Orłowska-Zwolińska, 1979).
The Carnian Pluvial Episode, correlated with the A. astigmosus zone (Fijałkowska-Mader et al., 2020), which took place later, resulted in the short-term development of hygrophylous flora. Sedimentation in Poland occurred in the deltaic and continental milieu (Dadlez et al., 1998). Ferns, mainly from the family Marattiaceae (documented by Leschikisporis aduncus and Apiculatisporites sp. div. spores), Osmundaceae (Todisporites sp. div. spores), and Cyathaeaceae (Deltoidospora sp. div. spores), horsetails (Calamospora tener spores), sphenophytes (Aratrisporites sp. div. and Kraeuselisporites sp. spores), bennettites producing pollen grains. Aulisporites astigmosus (Figure 1H) grew on the wet and marshy areas of floodplains, deltas, and lake shores. Conifers (represented by Ovalipollis sp. div. pollen grains) prefer drier areas (Roghi, 2004; Roghi et al., 2010).
In the Late Triassic, corresponding to the C. meyeriana zone, the climate became drier. The dominant component of the vegetation at that time was cheirolepidiacean conifers (producers of Corollina/Classopllis meyeriana (Figure 1J), Classopollis torosus, C. simplex, Geopollis zwolinskae, and Granuloperculatipollis rudis pollen grains), which are drought-resistant trees and shrubs that occur in a wide range of environments, including lakes and river banks, where they form dense thickets on floodplains, and on dry, elevated areas (Abbink, 1998; Alvin, 1982). Voltziales (documented by Brachysaccus neomundanus, Labiisporites triassicus, and Enzonalasporites sp. div. pollen grains) and plants similar to cedars (represented by Cedripites microreticulatus pollen grains) grew in dry zones of uplands and lowlands, whereas the wetter areas were colonized by gingkos-producing Monosulcites minimus pollen grains.
The Mid-Norian Pluvial Event caused a short-term increase in humidity (Berra et al., 2010; Preto et al., 2010; Szulc et al., 2015). The damp and wetland areas of the floodplains were inhabited by sphenophytes (represented by Calamospora tener and Equisetumsporites sp. spores), lycophytes (Anapiculatisporites sp. div. and Nevesisporites sp. div. spores), and ferns (Carnisporites granulatus and Deltoidospora sp. div. spores) (Mader, 1990b, 1997). Drier inland areas are covered by seed ferns, voltzialaeans (Ovalipollis sp. pollen grains), araucariceans (Callialasporites sp. pollen grains), and plants similar to cedars (Cedripites microreticulatus pollen grains). Cheirolepidiaceae was dominant along the banks of rivers and lakes. During this period, the remains of the voltzialean Patokaea silesica with polliniferous cones containing pollen grains of the genus Enzonalasporites were found in the fluvial-lacustrine deposits of Patoka (Upper Silesia) (Pacyna et al., 2017).
Triassic Rhaetian flora, which is correlated with the R. tuberculatus zone, is characterized by a strong domination of ferns and lycopsids. The sedimentation of the Rhaetian deposits of Poland occurred in a continental, mainly wet fluvial and lacustrine milieu (Dadlez et al., 1998). Ferns belonging to the Cyathaeaceae family (Deltoidospora sp. div. and Concavisporites sp. div. spores), Dipteridaceae (Dictyophyllidites mortoni and Baculatisporites wellamanii), Marattiaceae (Marattisporites scabratus spores), Matoniaceae (Concavisporites sp. div. and Zebrasporites sp. div. spores), and Osmundaceae (Todisporites sp. div. and Trachysporites fuscus spores), as well as horsetails (Equisetumsporites sp. spores), lycophytes (Lycopodiumsporites sp. div., Lycopodiacidites rugulatus, Densosporites sp. div., Camarozonosporites sp. div., and Acanthotriletes varius spores), and mosses (Sphagnumsporites sp. spores), overgrew the wet areas of floodplains, rivers, and lake banks (Abbink, 1998; Abbink et al., 2004a, 2004b; Van Konijnenburg-Van Cittert, 2002). The drier lowlands were inhabited by cycads/bennettitales (Cycadopites sp. and Chasmatosporites sp. div. pollen grains), erdmanithecaleans (Eucommiidites sp. pollen grains), gingkos (Monosulcites minimus pollen grains), gnetales (Gnetacaepollenites sp. pollen grains), ferns from the family Gleicheniaceae (Gleicheniidites senonicus spores) and Cyatheaceae (Petersen et al., 2013), as well as scarce pines (Pinuspollenites minimus pollen grains), Voltziales (Ovalipollis sp. div. pollen grains), Araucariaceae (Araucariacites sp. pollen grains), and Cheirolepidiaceae (Classopollis sp. div. and Rhaetipollis germanicus pollen grains), which grew in arid inland areas (Bonis & Kürschner, 2012; Kürschner & Herngreen, 2010). The latter also inhabited the coastal zones of the lakes. A characteristic element of this flora is the gymnosperm plant (Mander et al., 2012), which may be a conifer (Vajda et al., 2013) producing Riccisporites tuberculatus pollen grains.
The history of the development of the Triassic flora presented above is confirmed by the scarcity of plant macroremains (Pacyna, 2014).
. Palynofacial and Paleoenvironmetal Investigations
Palynofacial analysis has developed in different directions during several years of study, and different approaches have been used to define palynofacies (Tyson, 1995). A. Fijałkowska (1994a, 1995), who initiated the palynofacial investigations in the Lower Triassic deposits of Poland, defined palynofacies as “a distinctive assemblage of palynoclasts whose composition reflects a particular sedimentary environment,” taken from Powell et al. (1990), and used this analysis to determine the depositional milieu. Later, the studies were extended to the entire Triassic (Fijałkowska-Mader, Heunisch, & Szulc, 2015; Fijałkowska-Mader, Kuleta, & Zbroja, 2015).
. In Situ Pollen Grains in Pollen Cones and Inside Ovules (AF-M and JZ)
Due to the small number of macroremains of Triassic plants in Poland (Pacyna, 2014), finding the different reproductive organs of the Volzialean conifers Patokaea silesiaca, which contain pollen grains of the genus Enzonalasporites in the Upper Triassic, Norian deposits from Upper Silesia, is of exceptional importance. The pollen grains previously known to be dispersed only in sediment were, for the first time, combined with a parent plant. They were found inside the micropylar canal of the ovule (Figure 2A), at the micropylar end of the seed, and in the pollen sacs of polliniferous cones (Figure 2B,C). This is not only the first evidence of a relationship between Enzonalasporites pollen and the parent plant (Pacyna et al., 2017) but also a rare and valuable record of pollen grains found inside the micropylar canal of the ovule, fossilized at the precise moment of pollination.
. Jurassic and Cretaceous (JZ)
. Palynostratigraphy and Systematics
. Miospore Zones
The palynological (spore-pollen grain) zonation of the Jurassic strata of the Polish Lowland was first proposed by Rogalska (1976), who identified two spore-pollen assemblages: Lower and Middle Jurassic (from Aalenian to Callovian) and five microfloristic phases within the Lower Jurassic (I for Hettangian and Sinemurian, II for Lower Pliensbachian, III for Upper Pliensbachian, IV for Lower Toarcian, and V for Upper Toarcian). Later, the Conbaculatisporites mesozoicus-Dictyophyllidites mortoni-Cerebropollenites thiergarti zone was proposed for the Hettangian of NW Poland and the Cingulizonates rheticus-Limbosporites lundbladi zone for the Rhaetian (Pieńkowski et al., 2012).
A palynological zonation for the Lower Cretaceous (Berriasian, Valanginian, Hauterivian, Barremian, and Aptian) strata in Poland was established by Mamczar (1986), followed by the systematic revision of selected spores and pollen grains from Mesozoic sediments by M. Waksmundzka (1992, 2014).
. Megaspores Zones
The megaspore zonation for Jurassic strata was proposed by T. Marcinkiewicz (Marcinkiewicz et al., 2014, and literature cited therein), who proposed assemblages in the rank of assemblage zones, correlated with the standard stratigraphic division of the Early Jurassic: Nathorstisporites hopliticus (Ho) – Hettangian and Lower Sinemurian, Horstisporites planatus (Pl) – Upper Sinemurian – Pliensbachian, and Paxillitriletes phyllicus (Ph) – Toarcian. For the Middle Jurassic, only one assemblage with Horstisporites harrisi–Aalenian–Early Bathonian was established.
Mamczar (1984, 1986) proposed megaspore zonation for the Cretaceous for the Valanginian, Hauterivian, Barremian, and Aptian stages. Similarly, Marcinkiewicz (1962) and Mamczar (1986) conducted studies on Triassic/Jurassic and Jurassic/Cretaceous boundaries, respectively.
. Reconstruction of Polish Jurassic and Cretaceous Floras Based on Spores and Pollen Grains
Since the occurrence of macroremains is much more limited than that of microremains (spores and pollen grains), there are several localities from which the floral composition of the plant cover would remain unknown if there were no palynomorphs. Palynological studies on flora reconstruction are limited because of the nature of the material but can provide a general picture of the environment and its conditions. During the Jurassic period, the number of localities with macroflora decreased. While leaf fossils are common in the Lower Jurassic, they are less frequent in the Middle and Upper Jurassic. In this case, information obtained from palynological research is often the floristic data from given time periods and areas.
In Polish palynology, in addition to determining age, pioneering palynological investigations were aimed at wider floristic characteristics, including the reconstruction of the Early Jurassic palynofloras, as performed by M. Rogalska on the material from the Blanowice brown coal from the region of Zawiercie. This was also the first attempt to use palynological analysis for stratigraphy of the coal-bearing Mesozoic sediments of Poland. The coal examined was found to contain spores of bryophytes, pteridophytes (mainly ferns), and pollen grains of gymnosperms (mostly conifers) (Rogalska, 1954). Other Early Jurassic palynofloras were described from the Polish Lowland, from the Mroczków–Rozwady area, near the Opoczno (Rogalska, 1956) and Gorzów Wielkopolski, Międzychód, Aleksandrów Kujawski, and Gutwin boreholes (Rogalska, 1976). In these localities, fern spores were the most diverse (dominated by the number of species), whereas coniferous pollen grains were the most abundant. Spores of bryophytes, lycopods, and pollen grains of bennettitaleans, cycads, ginkgos, and seed ferns were also present (Rogalska, 1956, 1976, 1980a, 1980b).
In the Lower Jurassic palynoassemblage from Odrowąż (Mesozoic margin of the Holy Cross Mountains), spores of bryophytes, lycophytes, sphenophytes, ferns, and pollen grains of seed ferns, bennettitaleans, cycads, ginkgos, gnetales, and conifers were described, illustrated, and compared with megaflora from the same locality. The major plant groups of the microflora corresponded to the plant groups represented in the macroflora. Only the sporomorphs of bryophytes, lycophytes, and some conifers had no counterparts in the megafiora. This difference may be due to the differences in transport (sporomorphs originate mostly from long-distance transport, while macroscopic plant remains represent first of all the vegetation of the sedimentation basin and its nearby surroundings) and state of preservation (e.g., delicate bryophytes and lycophytes very rare in the sediment). The Hettangian age of the sediments from Odrowąż was confirmed based on miospores. It is worth noting that Aratrisporites minimus – the microspores of isoetalean lycophytes, fern spores (e.g., Todisporites minor, Osmundacidites sp. from Osmundaceae, Matonisporites sp. from Matoniaceae, Cyathidites minor, cf. Cyathidites australis from Cyatheaceae or Dicksoniaceae, and Plicifera delicata from Gleicheniaceae) and Classopollis torosus (Figure 2F) – pollen grains of Cheirolepidiaceae (extinct group of conifers) – are dominant (Ziaja, 2006).
The palynological data compiled by M. Hodbod from the Brody-Lubienia, Mechowo, and Parkoszowice drill cores were used to document changes in the floral composition among bryophytes, Equisetaceae, lycopsids, ferns, seed ferns, cycads, bennettites, ginkgophytes, and conifers, especially in the early Toarcian. An increase in the frequency of fern spores, mainly from the Cyatheaceae and Dicksoniaceae families, and a decrease in the frequency of conifers and other pollen-producing plants confirmed that the climate was relatively warmer. The increasing frequency of spores of bryophytes, horsetails, and lycopsids also indicates an increase in humidity at this time, which is related to geological and geochemical data, suggesting the presence of a warm and humid climate in the early Toarcian (Pieńkowski et al., 2016).
Middle Jurassic sporomorphs from the Międzychód borehole (Polish Lowland) and the NE Mesozoic margin of the Holy Cross Mountains indicate rich and diverse palynoflora (Rogalska, 1976, 1980a, 1980b). In addition to spores of bryophytes, lycophytes, and sphenophytes, numerous ferns were present, especially from the families Cyatheaceae, Gleicheniaceae, and Osmundaceae. Pollen grains of conifers indicate the dominance of Podocarpaceae, Araucariaceae, Pinaceae, and Taxodiaceae.
Spores of lycopods, ferns (Schizaeaceae, Cyatheaceae, Dipteridaceae, and Marattiaceae) and pollen grains of seed ferns, bennettitaleans, cycads, or ginkgos [e.g., Monosulcites minimus (Figure 2E)], conifers (Cheirolepidiaceae), and extinct gymnosperms family (Erdtmanithecaceae), represented by Eucommiidites troedsonii pollen grains (Figure 2H), were found in the Grojec Clays near Kraków (Ichas, 1986).
Another Middle Jurassic (Bathonian) locality in Gnaszyn (Kraków-Silesia Homocline) yielded spores of bryophytes, lycophytes, sphenophytes, and ferns, as well as pollen grains of seed ferns, bennettitaleans, cycads, ginkgos, and conifers. The most frequent were pollen grains of conifers, especially from the families Araucariaceae and Taxodiaceae, and fern spores from Osmundaceae, Cyatheaceae, Dicksoniaceae, Schizaeaceae, Gleicheniaceae, and Matoniaceae. The presence of these ferns and Araucariaceae indicates a warm climate (Gedl & Ziaja, 2012).
Upper Jurassic sediments were palynologically examined by J. Mamczar (1980, 1986) in central Poland (e.g., Żychlin near Kutno), where a predominance of Cheirolepidiaceae pollen grains was noted. Fern spores and sporadic bryophytes, lycophyte spores, pollen grains of other conifers, bennettitaleans, and cycads were less frequent. The Upper Jurassic (Oxfordian-Kimmeridgian) palynoflora showed a dominance of coniferous pollen grains, mainly from the family Araucariaceae – Araucariacites australis and Callialasporites trilobatus (Figure 2G) – from Wólka Bałtowska (NE margin of Holy Cross Mountains), indicative of a dry and warm climate in this area (Gedl & Ziaja, 2004).
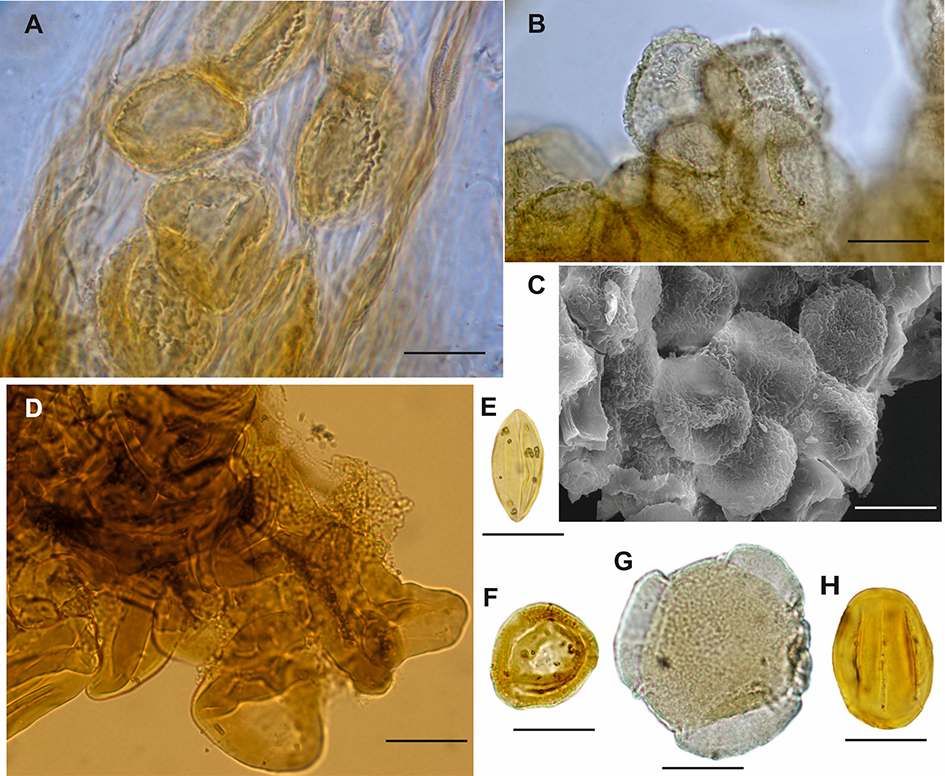
Figure 2
In situ miospores of Triassic and Jurassic and characteristic miospores of Jurassic: (A) Enzonalasporites sp. in situ pollen grains inside micropylar canal of the ovule of the voltzialean conifers Patokaea silesiaca, LM (phase contrast), KRA-PALEO 104/36, Patoka, Upper Triassic; (B) Enzonalasporites sp. in situ pollen grains from the pollen sac of the voltzialean conifers Patokaea silesiaca, LM, KRA-PALEO 104/100, slide KRA-PALEO 104/100/2, Patoka, Upper Triassic; (C) Enzonalasporites sp. in situ pollen grains from the marginal part of the pollen sac of the voltzialean conifers Patokaea silesiaca, SEM, KRA-PALEO 104/100, Patoka, Upper Triassic – stored at the Institute of Botany, Jagiellonian University, under the palaeobotanical collection number KRA-PALEO 104; (D) group of in situ spores from the matoniaceous fern Matonia braunii, LM, Niekłań, Lower Jurassic; (E) Monosulcites minimus pollen grains of ginkgos, LM, KRAM P PM Grojec II 53/6/07, Grojec, Middle Jurassic; (F) Classopollis torosus pollen grain of conifer (Cheirolepidiaceae), LM, KRAM P PM 68/6/1/6, Odrowąż (Sołtyków), Lower Jurassic; (G) Callialasporites trilobatus – araucariaceous pollen grain, LM, KRAM P PM 56/57II/1, Wólka Bałtowska, Upper Jurassic; (H) Eucommiidites troedsonii pollen grain from gymnosperms (Erdtmanithecaleans), LM, KRAM P PM Grojec II 53/6/07, Grojec, Middle Jurassic – stored in paleobotanical collection (Palaeozoic and Mesozoic) of the National Biodiversity Collection – Herbarium KRAM at W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków (KRAM P PM). Scale bar: 20 µm. Photos by J. Ziaja (A,B,D,H) and K. Bóka (C).
Early Cretaceous palynofloras of the Polish Lowland indicate a clear of change in plant composition towards a dominance of Lygodiaceae and Gleicheniaceae fern families, as well as conifers of Taxodiaceae, Araucariaceae, Podocarpaceae, and Pinaceae; while lycophytes, sphenophytes, seed ferns, bennettitaleans, cycads, and ginkgos became less common (Mamczar, 1984, 1986). A similar Early Cretaceous assemblage was found in the W-20 borehole drilled in the Piechcin-Barcin area in the Kujawy region of the Polish Lowlands, including bryophytes, lycophytes, ferns (mainly from Gleicheniaceae), seed ferns, conifers, and the first Clavatipollenites hughesi and Tricolpites sp. pollen grains of angiosperms of unknown parent plants (Waksmundzka, 1992).
The Late Cretaceous (Cenomanian-Turonian) palynoflora of N Poland (Chłapowo I and Chłapowo III boreholes) (Grabowska, 1987) was dominated by fern spores and angiosperm pollen grains, mainly of unknown affinity (30% with 3% of the Normapolles group). The Late Cretaceous (Upper Turonian) palynoflora from the Folwark Quarry (Opole, SW Poland) contained a rich sporomorph assemblage of bryophytes [e.g., Cingutriletes sp. (Figure 3A)], lycopods [e.g. Camarozonosporites sp. (Figure 3B)], ferns (Figure 3C), conifers [e.g., Pinuspollenites sp. (Figure 3D)], and angiosperms, mainly from the Normapolles group (Fagales) (Płachno et al., 2018), indicating a much more diverse vegetation than could be reconstructed based on macrofossils found in this area (Płachno et al., 2018). Santonian palynofloras from nine boreholes situated south of Bolesławiec, located in the North Sudetic Basin, were investigated by Raniecka-Bobrowska (1984, 1989) and Halamski et al. (2020). The dominant groups in Santonian palynoflora Assemblage A (from Lower Santonian?) were ferns (especially Gleicheniaceae) and representatives of angiosperms known only from their unusual pollen grains, namely the Normapolles group. Assemblage B (from Lower–Middle Santonian) was similar to Assemblage A but was dominated by the fern family Schizaeaceae (Halamski et al., 2020, and literature cited therein). Lush vegetation with numerous pteridophytes, mainly ferns from Schizaeaceae [e.g., Ruffordiaspora sp. (Figure 3G)] and Gleicheniaceae [e.g., Gleicheniidites senonicus (Figure 3E) and Clavifera triplex (Figure 3F)] in coastal zones and angiosperms, mainly from Normapolles group – Fagales [e.g., Trudopollis sp. (Figure 3H) Normapolles (Figure 3I)] in the inner parts of islands of the Łysogóry-Dobrogea Archipelago existing during the Lower Maastrichtian in SE Poland, based on data from the Pełczyska borehole (Jurkowska et al., 2019).
Figure 3
Characteristic spores and pollen grains from the Cretaceous of Poland. (A) Cingutriletes sp. spore of bryophyta (Sphagnaceae), Sample 2, Folwark Quarry, Upper Turonian; (B) Camarozonosporites sp., spore of Lycopodiaceae, Sample 2, Folwark Quarry, Upper Turonian; (C) Deltoidospora sp. fern spore of Cyatheaceae ? or Dicksoniaceae ?, Sample 1, Folwark Quarry, Upper Turonian; (D) Pinuspollenites sp. pollen grain of conifers Pinaceae, Sample 3, Folwark Quarry, Upper Turonian; (E) Gleicheniidites senonicus Ross, fern spore of Gleicheniaceae, Sample 22, Pełczyska, Lower Maastrichtian; (F) Clavifera triplex (Bolkhovitina) Bolkhovitina, fern spore of Gleicheniaceae, sample: Pełcz 23, Pełczyska, Lower Maastrichtian; (G) Ruffordiaspora sp., fern spore of Schizaeaceae, Sample: Pełcz 17, Pełczyska, Lower Maastrichtian; (H) Normapolles, pollen grain of Fagales, sample: Pełcz 22, Pełczyska, Lower Maastrichtian; (I) Trudopollis sp., pollen grain of Fagales, Sample 19, Pełczyska, Lower Maastrichtian. Scale bar: 20 µm. Photos by E. Worobiec (A–H).

. Palaeoenvironmental and Palynofacies Investigations
The reconstruction of the paleoenvironment is a valuable part of palynological investigations. The SEG model proposed by Abbink (1998), which assigns specific sporomorphs to plant groups, helps determine distinct habitats and has been used for palynological investigations in the Lower Jurassic (Krupnik et al., 2014) and Middle Jurassic (Gedl & Ziaja, 2012) of Poland.
Palynofacies analysis determines the relative abundance and type of sedimentary organic matter (Tyson, 1995). Combined with pollen analyses, a picture of some components of the environment and sedimentation processes has been obtained. The elements of organic particles in sediments are characteristic of certain types of paleoenvironments. This type of analysis has been used for Jurassic (Gedl & Ziaja, 2004, 2012; Krupnik et al., 2014; Pieńkowski et al., 2012, 2016; Pieńkowski & Waksmundzka, 2009) and Cretaceous sediments (Jurkowska et al., 2019; Leszczyński & Waksmundzka, 2013; Płachno et al., 2018).
. In Situ Spores and Pollen Grains in Sporangia and Pollen Sacs and Inside Ovules
Spores and pollen grains can sometimes be found in situ inside the reproductive organs of parent plants (sporangia or male cones). Such preserved microfossils are useful for the identification of parent plants, as well as for the reconstruction of vegetation. The first findings of Polish Jurassic spores in situ were prepared and described by Reymanówna (1963). Sporangia were found on the frond of the matoniaceous fern Phlebopteris angustiloba from Grojec near Kraków (southern Poland, Middle Jurassic). Other matoniaceous spores in situ were isolated and described from the sporangia of the fern Matonia braunii (Figure 2D) from Niekłań PGI 1 borehole (central Poland, Holy Cross Mts, Lower Jurassic). These smooth spores, triangular in outline, possess a triradiate mark and interradial and apical thickenings, and are similar to Dictyophyllidites mortoni, known as dispersed spores from sediment (Barbacka et al., 2016).
The pollen grains of Classopollis torosus were isolated from polliniferous cones of Hirmeriella muensteri (Cheirolepidiaceae) from the Odrowąż (= Sołtyków) outcrop section (central Poland, northern Mesozoic margin of the Holy Cross Mountains, Lower Jurassic). These unique pollen grains are oval to circular in outline with a circular groove or thinning, parallel to the equator of the pollen grain, with a circular cryptopore on the distal side and triangular scar with concave sides on the proximal side found only in the fossil record (Barbacka et al., 2007; Ziaja, 2006).
Rarely, pollen grains are preserved inside the dispersed seeds. Jurassic Eucommiidites pollen grains, which were initially believed to come from an angiosperm, were found in the micropyle and pollen chamber of the Middle Jurassic in ovules Allicospermum retemirum from Grojec (Reymanówna, 1968). Its affinity was confirmed by the presence of a pollen chamber lacking in the ovules of angiosperms, conifers, and bennettitaleans. This record excluded the possibility of angiospermous affinity of the parent plant. Allicospermum retemirum, initially considered to be a seed of seed fern fructification, is now considered to belong to the gymnosperm order Erdtmanithecales, as the pollen grains of Eucommiidites are diagnostic for this order.
Other pollen grains (Alisporites sp.) were also found in the pollen chamber and were described from the other ovules, Allicospermum szaferi from Borzynowo near Elbląg (northern Poland, Lower Jurassic). The ovules were confirmed to be seed ferns (Wcisło-Luraniec & Ichas-Ziaja, 1990).
. Summary
The Mesozoic is an interesting era for palynological investigations. Triassic palynological studies have been conducted at numerous localities in Poland. Within this field, the most important achievements of Polish Triassic palynology include the proposal and development of the palynostratigraphic zonation of the Triassic deposits based on miospores and megaspores, as well as the reconstruction of the Triassic floras based on miospore assemblages. An equally significant finding was the association of Enzonalasporites pollen grains with the Patokea silesiaca conifer parent plant, which is the first in situ finding of Enzonalasporites pollen grains, which were previously known only from sediment. Jurassic palynostratigraphy is less precise due to the difficulties associated with establishing the corresponding zones. On the other hand, the reconstructions of Jurassic flora in Poland has significantly supplemented the picture of vegetation during this period, known from macroflora studies. Assigning Jurassic spores or pollen dispersed in the sediment to parent plants, for example Dictyophyllidites mortoni spores found in Matonia brauni sporangia or Classopollis torosus pollen grains isolated from polliniferous cones of Hirmeriella muensteri, is a valuable contribution for plant systematics. Although Cretaceous palynological studies are scarce because of the small number of localities, palynostratygraphy and vegetation reconstruction are very interesting. Data obtained from palynological studies of the Triassic, Jurassic, and Cretaceous strata have significantly expanded our knowledge of the vegetation, environment, and climate of these periods.
