. Introduction
The Palaeozoic Era witnessed terrestrialization processes which changed the appearance of our planet on a global scale (Gensel & Andrews, 1984; Servais et al., 2019; Taylor et al., 2009). Lands were covered bit by bit by dense vegetation from coastal lowlands to highlands. Initially, in the Ordovician and Silurian, very small bryophyte-like plants covered coastal lowlands (Steemans et al., 2012). In the Devonian the first shrubs and trees rapidly appeared (Gensel & Andrews, 1984). In the Carboniferous, vast coal forests dominated by huge lycopsids and sphenopsids developed (Bateman et al., 1992). They provided the coal that fueled the Industrial Revolution. In the Permian, aridification caused the collapse of the giant lycopsids, sphenopsids and ferns. At the same time, gymnosperms diversified and filled many new niches in dry areas.
The oldest plant remains are rare and usually poorly preserved. Our knowledge of them is still insufficient, although they are very important from an evolutionary point of view (Edwards & Kenrick, 2015). In Poland there are only a few localities that have revealed fossils of the earliest land plants (Figure 1, Table 1). Historically the first recognized and best-documented plants are Devonian (Czarnocki, 1919; Zdebska, 1972, 1978, 1982, 1986a, 1986b, 1993a). Research in recent years provided some Silurian and Ordovician remains that have been only preliminarily announced so far (Bodzioch et al., 2003; Salamon et al., 2018).
Figure 1
Plant macrofossil localities from the Polish Devonian: 1 – Lubiechów; 2 – Mokrzeszów; 3 – Wilcza; 4 – Andrychów 2, 4 boreholes; 5 – Kostomłoty; 6 – Dyminy IG2 borehole; 7 – Zbrza; 8 – Modrzewie 2A borehole; 9 – Winnica near Nowa Słupia; 10 – Pionki borehole; 11 – Suskowola borehole; 12 – Łagów; 13 – Ciepielów borehole; 14 – Opole Lubelskie borehole; 15 – Dorohucza borehole.
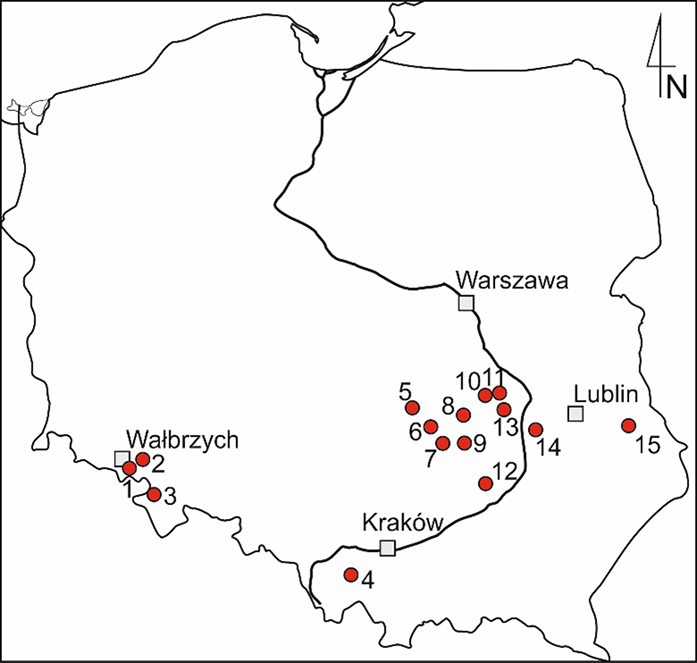
Table 1
Stratigraphic position of the floras and localities discussed in the text.
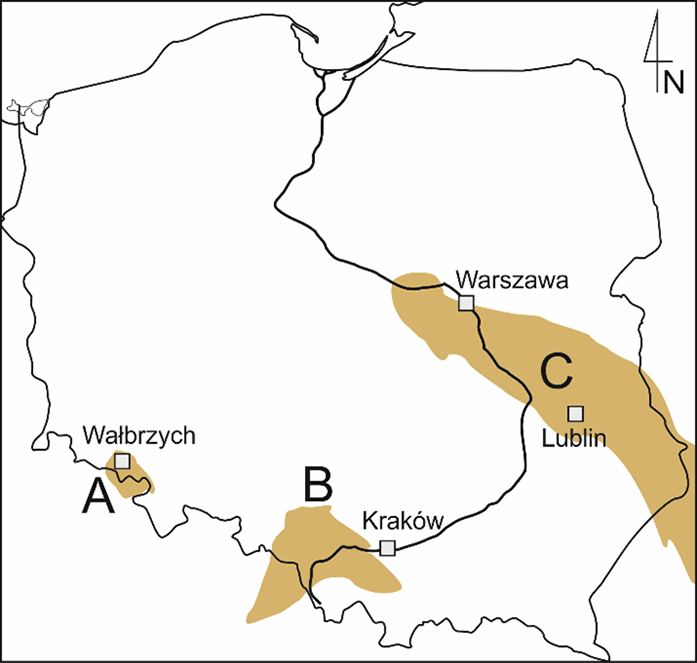
In contrast to this, the palaeobotany of Carboniferous (358–298 Ma) macrofossils has a long tradition in Poland and has yielded great achievements (Pacyna, 2012). Poland possesses three large Carboniferous coal basins (the Upper Silesian, Lower Silesian, and Lublin coal basins; Figure 2), intensively exploited over several centuries, which provided a large number of plant fossils, some very important and iconic in the annals of palaeobotany (Zdanowski & Żakowa, 1995). The best example is a seed fern Lyginopteris hoeninghausii leaf specimen from Czernica in Upper Silesia, illustrated in many palaeobotanical textbooks in Poland and worldwide (e.g., Taylor et al., 2009, Figure 14.29), and still housed in a Berlin museum (Krings & Schultka, 2000, Plate 3).
. Discovery of the Oldest Land Plants From Poland (Ordovician – Devonian) – Description and Phylogenetic Interpretation
. Ordovician and Silurian Plant Remains
The discovery of Ordovician land plants from Poland was announced by Kozłowski and Greguss as early as 1959 (Kozłowski & Greguss, 1959). The presumed land plants were thoroughly documented by Greguss (1961, 1962), recognized as members of the hepaticophytes and bryophytes. However, it is now believed that these alleged fossils are degraded roots of extant plants which struck erratic boulders.
Recently, Salamon et al. (2018) documented from the Uppermost Ordovician (Hirnantian) of the Zbrza locality in the southern Holy Cross Mountains minute shoot fragments, some of them dichotomously branching, others with probable sporangia at the top. Morphologically, the plants are comparable to the younger plant macrofossils, including Hostinella, but rather poorly preserved. These remains are among the oldest land plants documented by macrofossils worldwide (cf. Edwards & Kenrick, 2015; Steemans et al., 2012; Tomescu, 2022). Probable stomatal complexes from Zbrza are similar to those described by Snigirevskaya et al. (1992) in the putative land plant Akdalaphyton caradocki from the early Late Ordovician (Sandbian) of Kazakhstan. This find is consistent with current opinions about the time of emergence of the first land plants (cf. Servais et al., 2019). However, further research is needed to confirm this discovery and to better document the finds.
The high potential of the Holy Cross Mountains to provide fossils of the oldest land plants was noted already by Czarnocki (1919), but the remains he discovered were never properly documented. Only rather recently, Bodzioch et al. (2003) described the first Silurian land plants from the Holy Cross Mountains. The fossil assemblage occurs in a near-shore marine, terrigenous deposit of Late Silurian (Ludlowian/Pridolian) age in Winnica near Nowa Słupia. It contains Cooksonia sp. – one of the oldest land plants – some similar species, and some problematic forms. This flora is among the world’s oldest well-dated Cooksonia floras from the Silurian (cf. Servais et al., 2019, Figure 2).
Szrek et al. (2015) documented horizons with plant root impressions and layers interpreted as palaeosols showing different stages of pedogenesis from the upper Pragian–lower Emsian of Kostomłoty in the north-eastern part of the Kielce region in the Holy Cross Mountains. These plant root impressions are among the oldest evidence of the existence of large roots in the Early Devonian and provide new data on terrestrialization processes at the beginning of the Devonian.
. Anatomy, Morphology, and Evolutionary Significance of Early Devonian Plants
Remains of Devonian plants from Poland have been noted by geologists since the end of the nineteenth century, and horizons rich in plant remains were reported long ago from the Holy Cross Mountains (e.g., from Łagów) by Czarnocki (1919). As a rule, however, these remains were poorly preserved and not numerous, and outcrops of Devonian rocks are rare in Poland. The situation changed in the 1960s and 1970s, when an intensive program of drilling in search of geological resources provided material from deep cores (Figure 1). It turned out that Devonian rocks underlie a large part of Polish territory but most often at depths of more than 1,000 m. Careful inspection of cores by geologists has produced very rare, interesting and well-preserved remains of Early Devonian plants (Zdebska, 2003).
The first description of Early Devonian plants from deep boreholes was prepared by Jakubowska (1968). From two cores from Ciepielów and Dorohucza she described several taxa of psilophytes and primitive lycophytes, but based only on the external morphology of the remains.
Based on material from deep boreholes, Zdebska (1972, 1978, 1982, 1986a, 1986b, 1993a) and Brzyski (1976) documented many Early Devonian taxa including a new genus and two new species. Their research was very detailed and included analyses of all available morphological and anatomical details. Many remains were pyritized with the internal structure preserved. Pyrite petrifactions are very difficult to process, destructive, and requiring special, very time-consuming preparation methods. Despite the difficulties, they obtained spectacular results which are significant achievements of Polish palaeobotany. That work concentrated on the biological and evolutionary interpretation of the described fossils.
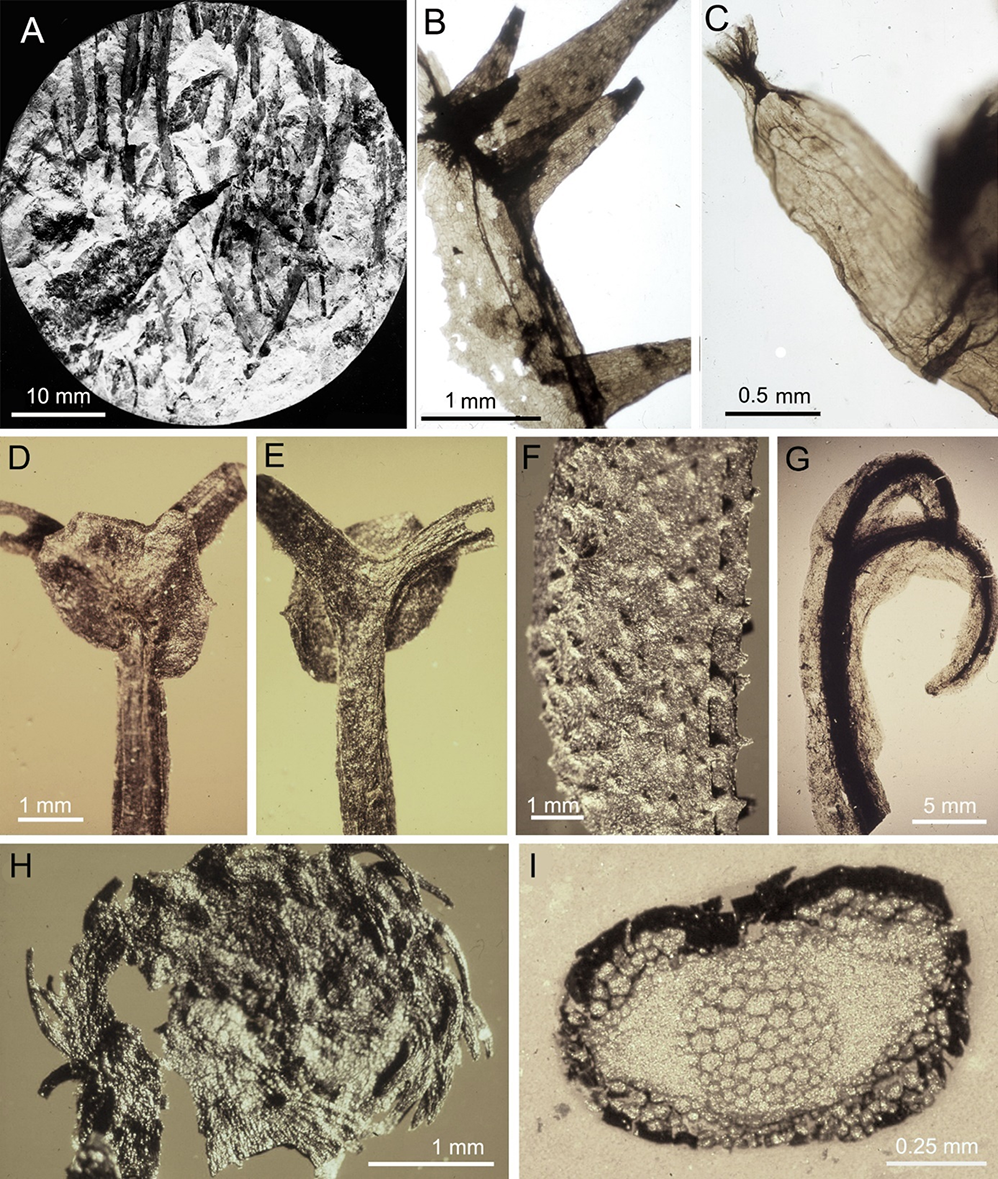
A member of the zosterophyllophytes Sawdonia ornata was described by Zdebska (1972) from the Lower Devonian (Emsian) from a borehole at Opole Lubelskie (Figure 3A–C). The plant possessed dichotomously branched shoots covered with numerous spines that were very diverse in shape and size (Figure 3A,B). Some spines possessed widened vesicular ends and broken off apices, and were filled with remains of secretion (Figure 3C). This type of secretory spine might protect a plant against herbivores. This finding is an important contribution to the evolution of plant–animal interactions. It was not documented for zosterophyllophytes before and is still one of the few plant–animal interactions documented for the Early Devonian (cf. Labandeira, 1998). The Sawdonia specimens showed well-preserved cuticle structure but no internal anatomy. Zdebska (1972) described well-preserved stomata showing new details previously unknown in the genus Sawdonia (see, e.g., Rayner, 1983). She also discussed and clarified the taxonomic status of Sawdonia ornata on the basis of numerous carefully documented, well-preserved Polish specimens.
A new genus and species of zosterophyllophytes Konioria andrychoviensis was proposed by Zdebska (1982) based on well-preserved pyritized axis fragments from the Lower Devonian (Emsian) of two boreholes in the Bielsko–Andrychów area of the Polish Western Carpathians (Figure 3D–I). The axes dichotomize isotomously and anisotomously, form hooks at the apex, and are covered on their lower part with long subulate spines and on their upper part with short triangular spines (Figure 3F). In addition to spines, along the axes one–four longitudinal wings were also found. Very young shoots possessed circinately coiled apices (Figure 3H). Single reniform to rounded sporangia were borne at the points where the shoot dichotomises (Figure 3D,E). The position of sporangia at the final or penultimate dichotomy of the shoot, and their attachment, directly or with a very short stalk, are especially distinctive for this plant among the zosterophyllophytes. The xylem is elliptical in transverse section and exarch (Figure 3I). Tracheids are reported as having scalariform pitting. A peripheral hypodermal layer of thick-walled cells is present. Cuticular features are poorly preserved but include possible stomata (Figure 3G). Thanks to this study, Konioria can be included among the best-documented zosterophyllophytes worldwide and is widely discussed in many textbooks, research and review papers (e.g., Cascales-Miñana & Meyer-Berthaud, 2014; Gensel & Andrews, 1984; Taylor et al., 2009). Detailed documentation by Zdebska (1978) enabled this species to be used in cladistic analysis of early land plants, as one of the few analyzed zosterophyllophytes (Kenrick & Crane, 1997b). This plant is important from an evolutionary point of view. Its terminal branches with a single sporangium may have evolved into the divided leaves of Protolepidodendrales (Zdebska, 1982, 1993b). From the same Andrychów 2 and Andrychów 4 boreholes Zdebska (1982) isolated well-preserved cuticles of the early lycophyte Drepanophycus spinaeformis (Figure 4A,B).
Figure 3
Early Devonian (Emsian) zosterophyllophytes from Poland. Sawdonia ornata, Opole Lubelskie 3 borehole: (A) Core fragment with numerous axes covered with spines, hand specimen 16; (B) Fragment of shoot epidermis with spines, slide 17/10; (C) Spine with widened vesicular end which is open, slide 17/2. Konioria andrychoviensis, Andrychów 2 and Andrychów 4 boreholes: (D,E) Fragment of axis with sporangium situated at the level of a dichotomy (both sides), S/98/31; (F) External surface of a pyritized axis with numerous short spines, S/98/23; (G) Shoot apex showing dark core and stomata as dark spots, S/98/29; (H) Circinately coiled young axis densely covered with long spines, S/98/25; (I) Transverse section of axis showing xylem strand, specimen unnumbered. All specimens stored in Palaeobotanical Collection, Institute of Botany, Jagiellonian University (abbreviated as PC IB UJ) (photos by D. Zdebska).
A new species of the trimerophytes (basal euphyllophytes), Psilophyton szaferi, was proposed by Zdebska (1986b) for Lower Devonian (Emsian) specimens from the Modrzewie borehole near Bostów (Holy Cross Mountains). The plant branching is dichotomous, and unequally dichotomous where the wider branch tends to form an apparently main axis. On their surface the shoots show irregularly distributed enations in the form of rounded swellings (Figure 4D). The apices of sterile axes are forking. Fertile axes end in clusters of fusiform upright sporangia arranged in pairs. The structure of Psilophyton szaferi shoots is preserved in compression (shoot morphology and epidermis structure observable) and pyritized fossils. Pyritized axes show the anatomical structure of the vascular strand (endarch xylem) and accompanying secretory cells, which were described by Zdebska (1993a) for the first time in the genus Psilophyton (Figure 4C). The secretory cells in P. szaferi surround the vascular strand and are located in the place where there is endoderm in some ferns. Similar secretory cells are present in some primitive ferns (e.g., in Botryopteris). This feature may indicate that P. szaferi belongs to the line of basal euphyllophytes from which monilophytes (ferns and sphenopsids) originated. Based on transverse sections of pyritized axes, Zdebska (1993a) documented divisions in the xylem strand and secretory cell ring at shoot dichotomies, and a mode of shoot branching characteristic for trimerophytes, called trifurcation. Such detailed analyses of shoot divisions are very rare in Early Devonian plants worldwide. P. szaferi represents an intermediate but rather early form in the evolutionary sequence of megaphyllous leaves.
Zdebska also described some foreign material of the oldest land plants. In a paper from 1980 (Lemoigne & Zdebska, 1980), the structure of water-conducting cells in the protracheophyte Aglaophyton from Early Devonian Rhynie chert was documented. This was pioneering research and one of the first such papers in the palaeobotanical literature (see, e.g., Edwards, 2003); in recent decades this topic has been of great interest (e.g., Cascales-Miñana et al., 2019; Friedman & Cook, 2000; Kenrick & Crane, 1997a). In the last few years, the structure of the wall of water-conducting cells has been found to differ in major lineages of land plants, providing important phylogenetic information.
An interesting and enigmatic group from the Polish Early Devonian, the nematophytes, was first documented by Brzyski (1976). The nematophytes are a diverse and curious assemblage of early Palaeozoic fossils which according to different researchers may belong to the bryophytes, algae, fungi or lichens (Figure 4E,F). Brzyski (1976) recognized small plectenchyma fragments of the fungus-like or lichen-like genus Prototaxites composed of elongated tubes with embedded filaments. Together with Prototaxites were remains of some small (3–4 mm in diameter) spherical fossils called Pachytheca. Only the outer layer (cortex) consisting of tubes was preserved; the inner zone (medulla) was degraded. Brzyski (1976) confirmed that the coexistence of these two genera in Polish material is consistent with the previously formulated hypothesis that Pachytheca is the reproductive structure of Prototaxites. In material from the same Suskowola borehole, Brzyski (1976) found very interesting sporophytes terminated with a sporangium, determined by him as Sporogonites chapmanii. This fossil probably is an early bryophyte. S. chapmanii was documented from Poland for the second time in the world after its first description from the Early Devonian of Australia.
From the same Suskawola borehole, Zdebska (1978) documented thallus fragments determined as Spongiophyton sp. 2, showing cellular structure and pores (Figure 4F). Later she analyzed in detail the pores in the cuticle and compared them with stomata of living liverworts, pointing out that this plant may indeed represent some extinct type of hepatic (Zdebska, 1986a). According to research by Edwards (1982) these remains should be better referred to the genus Nematothallus. Zdebska (1978) referred to Spongiophyton sp. 1 a fragment of dichotomously branching thallus with circular pores from the Early Devonian from the Andrychów borehole (Figure 4E).
Figure 4
Early Devonian (Emsian) plants from Poland. Lycophyte Drepanophycus spinaeformis, Andrychów 2 and Andrychów 4 boreholes: (A) fragment of shoot cuticle with two leaves; (B) stoma; specimens unnumbered. Trimerophyte Psilophyton szaferi, Modrzewie 2A borehole: (C) Transverse section of axis showing a dividing vascular strand below branching (black arrow) and secretory cells (white arrow), S/99/8; (D) Fragment of axis impression showing irregularly distributed enations in the form of rounded swellings. Nematophyte Spongiophyton. (E) Spongiophyton sp. 1 – fragment of dichotomously branching thallus, outer side of upper surface, slide 10/1, Andrychów 2 borehole; (F) Spongiophyton sp. 2 – fragment of thallus showing outer side with pores, Suskowola borehole. All specimens stored in PC IB UJ (photos by D. Zdebska).
Filipiak and Zatoń (2011) documented plant and animal cuticle remains from the Lower Devonian of southern Poland from boreholes in the Upper Silesian Block and Holy Cross Mountains. From the latter region the most interesting fossils came from the Dyminy IG2 borehole, yielding small cuticle fragments with stomata belonging to the lycophyte Drepanophycus and zosterophyllophyte Sawdonia, enigmatic nematophyte cuticle fragments classified as Nematothallus and Cosmochlaina, and isolated banded tubes probably also of nematophyte origin. This is the third report of nematophytes from Poland, with two new taxa of them recognized for Poland (Kondas, 2018).
. Middle and Late Devonian Floras
Middle and Late Devonian floras are rare in Poland (Table 1). Kuchciński (1964) documented a Middle Devonian flora from the Wilcza Beds in the Middle Sudetes. Lycophytes, progymnosperms and early ferns were present in the plant assemblage but were preserved as highly fragmented remains. This paper was important biostratigraphically and clarified some problems in the very difficult stratigraphy of the Sudetes.
The best-known Upper Devonian floras were described from Mokrzeszów and Lubiechów in Lower Silesia by the German researchers Gothan and Zimmermann (1932, 1937). These floras consist of diversified lycophytes, some sphenophytes and progymnosperms, including beautiful specimens of Archaeopteris leaves.
Upper Devonian plant remains from the Holy Cross Mountains have been noted by many geologists since the end of the nineteenth century but as rule they were poorly preserved and not analyzed by palaeobotanists. One of the localities that provided the most numerous but mostly unidentifiable remains of Famennian age is Kowala Quarry. Only fairly recently, Dąbrowska and Filipiak (2006) documented some determinable plant remains from this locality, including branches and sporangia probably belonging to the progymnosperm Archaeopteris.
. Carboniferous Coal Forest – Taxonomic Composition, Changes Over Time, and the Plant Macrofossils-Based Biostratigraphy of Carboniferous Strata
The first scientific descriptions of Carboniferous plants originating from the present-day territory of Poland (Lower Silesia) were included in Sternberg’s fundamental palaeobotanic work (1820–1838). The end of the nineteenth and early twentieth centuries was a period of intense activity in Silesia by outstanding German-speaking palaeobotanists and geologists from Prussia and the Austro-Hungarian Empire, in particular H. R. Goeppert, D. Štúr, H. Potonié, F. Roemer, W. Gothan, and F. Zimmermann. They used Silesia as a source of valuable and interesting fossil materials, but their studies were largely random and not systematic. The material they collected has disappeared or has been taken out of Poland and housed in many German museums, where they are still used for taxonomic reviews and descriptions of new taxa. The Polish borders changed substantially after World War II; since 1945, almost the entire territory of Silesia belongs to Poland.
Studies of Carboniferous floras in Poland have focused mainly on biostratigraphy; biologically oriented palaeobotanical papers are rarer. Many of the published papers are descriptions or enumerations of floras from separate horizons, beds, or boreholes. Only some of these accounts are adequately documented with good illustrations.
. Plant Macrofossil-Based Biostratigraphy of Carboniferous Strata
The first Polish palaeobotanical papers on the Carboniferous were published by Tondera (1888, 1889, 1890). The next papers, written by Rydzewski (1913, 1915) were devoted to biostratigraphy based on plant macrofossils. In the interwar and post-war period, Bocheński, Czarnocki, and Stopa published many short biostratigraphic reports of Upper Silesian Coal Basin floras but without descriptions and illustrations of the taxa (e.g., Stopa, 1954, 1957a).
After World War II, the coal industry developed very rapidly, providing a huge amount of material for research. However, studies of Carboniferous floras were almost solely biostratigraphic in nature (e.g., Kuchciński, 1986; Lipiarski, 1981). During examinations of drilling cores and open pits, attention was focused mainly on the indicator species, the most commonly found and stratigraphically important ones. The species determinations were rather cursory; the same species names are repeated from many different ages and remote areas. Taxa that were relatively seldom found, especially plant reproductive structures, were ignored as insignificant biostratigraphically and difficult to classify, or were assigned only to genus rank (see comments in Pacyna, 2012). As a result, the documentation of different taxonomic groups is unequal. Best known are arborescent lycophytes (Lepidodendron, Sigillaria), calamitaleans (Calamites shoots, Annularia and Asterophyllites leafy branches), and fern and seed fern leaves (called pteridophylla); less well known are cordaitaleans, ferns, and the reproductive structures of all groups. Biostratigraphers used organ taxa but did not correlate them with each other. The fossil plant specimens from cores were small and fragmentary (depending on core diameter). Large specimens in which different organs are preserved in organic connection were unfortunately not collected from spoil tips and underground mines. The studies created a huge database but used the data only for biostratigraphy, summarized in several thematic volumes (e.g., Kotasowa & Migier, 1983, 1995; Migier, 1966, 1972a, 1972b, 1980, 1988; Zdanowski & Żakowa, 1995). Polish Carboniferous floras have been well documented to a degree comparable with work from Western Europe (cf. Bek, 2017; Opluštil et al., 2022), and huge specimen collections have been made. Changes in floras over time have been noted but were not considered in their palaeoecological and evolutionary context.
Among the Carboniferous macroflora researchers, the most prominent were A. Kotasowa and T. Migier. During many years of work in the second half of the twentieth century they documented numerous Polish Carboniferous floras, but mainly in terms of biostratigraphy (Kmiecik & Migier, 1979; Kotasowa, 1968, 1975b, 1979; Migier, 1971b, 1973, 1979). Only a few of their papers described the analyzed plants from a more botanical point of view. Migier (1971a) published a monographic description of medullosalean seed fern leaves of the genus Neuropteris (Paripteris subgroup) from all Polish coal basins. She discussed their taxonomy, variability and biostratigraphic significance. Kotasowa (1975a) devoted a long paper to the very important lyginopteridalean Lyginopteris hoeninghausii – the first reconstructed seed fern and an important biostratigraphic marker. Based on material collected from the Upper Silesian Coal Basin, she discussed and illustrated its variability (including subspecies distinguished by earlier researchers) and commented on reconstruction of the whole plant. In 1978, Kotasowa documented callistophytalean seed fern leaves from Upper Silesia (Kotasowa, 1978). Summarizing their research, Kotasowa and Migier (2001) published a valuable synthesis of knowledge of the Polish Carboniferous floras, including not only taxonomic descriptions but also high-quality photographic documentation.
. Anatomy, Morphology, and Palaeoecology of Carboniferous Plants
Biologically oriented palaeobotany papers are rather rare in research on Polish Carboniferous plants. Rydzewski (1919) prepared a monograph on Polish Carboniferous arborescent lepidodendrid lycopsids. Stopa (1957b) prepared a monograph, the only to date in Polish palaeobotany, on fern and seed fern leaves (called together pteridophylla if preserved in sterile state). He documented and illustrated many taxa, proposed some new ones, and discussed problems in descriptive terminology and systematics, proposing some solutions. In detail, Stopa discussed the stratigraphic and geographic ranges of the recognized species and documented their biostratigraphic importance. Bocheński (1960) analyzed in developmental terms the venation of the medullosalean pteridosperms Alethopteris and Lonchopteris. He confirmed the evolutionary connection between these genera, described and illustrated how pinnate venation may transform to reticulate venation during the origination of the genus Lonchopteris, and corrected errors made by earlier researchers in describing venation details in these genera.
Sagan conducted detailed research on Carboniferous floras of Lower Silesia. He published an extensive paper on the lyginopteridalean seed fern Sphenopteris bermudensiformis (Sagan, 1965). On the basis of rich material from the Namurian A of the Wałbrzych beds, he described new morphological details of the construction of the leaf, including the triply branched axis. He stated that the middle branch of the leaf axis was fertile and had cupules located on it. Sagan (1980) also described rich material of the herbaceous lycopsid Eleutherophyllum from the Namurian A of Lower Silesia. In E. mirabile he found sporangia and spores for the first time. In E. drepanophyciforme and E. waldenburgense he described the development and construction of sporangia and their location on the shoots. Wolf et al. (1977) described the leaves of the seed fern Sphenopteris (Lyginopteris) völkeli and the pollen organs connected with them from the Namurian A. On the basis of rich palaeobotanic material he had collected, Sagan published two atlases (Sagan, 1981, 1983), in which many sphenopsid and lycopsid species were described and illustrated.
Domagałowa (1971) revised the arborescent lycopsid Sigillaria from the Upper Silesian Coal Basin. She discussed the biostratigraphic utility and intraspecies variability of different Sigillaria species, proposed some new species, and documented all the described species with high-quality photos.
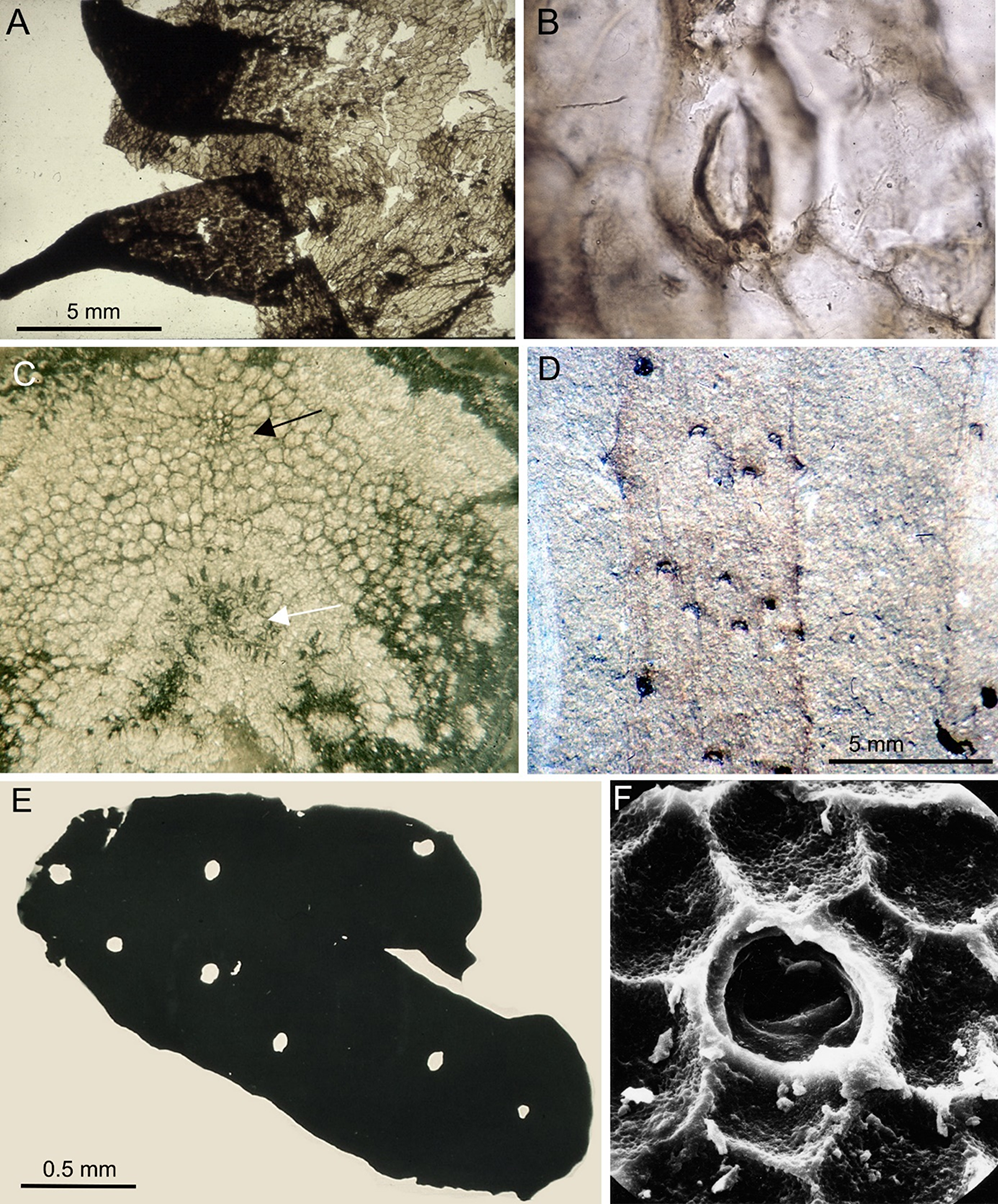
In several papers, Brzyski (Brzyski, 1965, 1969b, 1980, 2001; Brzyski & Stuchlik, 1992) described petrified plant remains with well-preserved tissue structures from the Carboniferous (Namurian A) of the Upper Silesian Coal Basin (Figure 5A–D). Coal balls have not been discovered in Polish coal measures so far (Brzyski, 1969a). The fossils described by Brzyski are the only well-documented anatomically preserved fossil plants from the Polish Carboniferous. Although coniferophyte secondary wood (genus Dadoxylon), sometimes preserved as large logs, has been described from the Lower and Upper Silesian Upper Carboniferous (Reymanówna, 1962), only the material described by Brzyski showed all tissues preserved, including even the delicate parenchyma. About twenty species were recognized by Brzyski, including lepidodendrid branches (Lepidodendron, Lepidophloios, Sigillaria; Figure 5A), calamite shoots (Arthroxylon), fern phyllophores (Ankyropteris, Etapteris; Figure 5B), arborescent fern stem fragments (Psaronius; Figure 5D), seed fern stems (Medullosa; Figure 5C) and petioles (Rachiopteris), and coniferophyte wood (Dadoxylon). He reported many genera and species for the first time from the Carboniferous of Poland. The plant tissues were permineralized by different, sometimes coexisting minerals like pyrite or calcite, and also coalified to some extent, so slide preparation was very difficult and time-consuming. Brzyski very carefully documented all observable anatomical features and compared in detail the Polish specimens to those preserved in coal balls from Western Europe.
An interesting phenomenon characteristic especially for the Carboniferous coal basins is preservation of upright plant stems in the growth position. Brzyski et al. (1976) and Gradziński and Doktor (1995) recorded upright Calamites and Sigillaria stems from several localities in Upper Silesia. They documented the “candelabrum” growth of Stylocalamites side stems in plants that had been buried by flood sediment. The longest stems of Sigillaria were up to 6 m high. Gradziński and Doktor (1995) concluded that the burial and preservation of the upright stems was facilitated by fast accretion of sediment and by local subsidence caused by intense compaction of peat. This compaction resulted in lowering of the local depositional surface, which promoted rapid formation of anomalously thick crevasse splays, enhancing the burial and preservation of stems to considerable heights. Gradziński et al. (1982) documented accumulation of drifted logs mainly of Cordaites and other large plant debris in a Carboniferous fluvial channel at Czerwionka Quarry in Upper Silesia. Most of the logs were filled with sand. Long logs lay horizontally and were preferentially oriented transversally to the channel axis. The accumulation of large plant debris at Czerwionka was deposited in an active channel of a large river. Heavy, partly rotted and waterlogged logs were dragged as bedload along the bottom, successively trapped, and buried with sand.
Florjan (e.g., Florjan & Żołdani, 1999) studied dispersed cuticles isolated from Carboniferous coals (Figure 5E,F), a topic very rarely taken up earlier by Polish researchers. In cooperation with the Czech palaeobotanist Šimůnek, he documented various dispersed cuticles of Carboniferous cordaitaleans from numerous Silesian coal seams (Šimůnek, 2015; Šimůnek & Florjan, 2013a, 2013b, 2019). They found that the most important difference between cordaitalean cuticles from coal (peat-forming vegetation) and cuticles from mudstone (roof-shale floras, “in situ” cuticles isolated from leaf compressions) is the presence of papillae on cuticles from coal samples but their absence or very seldom occurrence on cuticles isolated from compressions. Their cuticular studies suggest that the peat-swamp cordaitaleans were taxonomically distinct from those of more seasonally dry settings. For the purpose of classifying cordaitalean dispersed cuticles and because it is impossible to match adaxial and abaxial cuticles of the same species within dispersed cuticular assemblages, two new fossil genera were proposed: Cordaadaxicutis for adaxial cuticles and Cordaabaxicutis for abaxial cuticles, with over a dozen new species. Cuticular analysis of cordaitalean foliage appears to indicate larger numbers of species than does analysis based on leaf morphology alone. Based on the literature and on their own observations during research on fossil plants, Florjan and Worobiec (2016) prepared a textbook on plant taphonomy and fossilization modes.
Figure 5
Carboniferous plants from the Upper Silesian Coal Basin. Petrified remains (A–D) and dispersed cuticles (E,F). Transverse sections through petrified remains from the Upper Carboniferous (Namurian A): (A) Lepidophloios cf. wünschianus Carruthers – transverse section of young stem, specimen II-B, Jankowice coal mine; (B) Etapteris cf. diupsylon Williamson – transverse section of leaf axis (phyllophore), H-shaped xylem stele visible, specimen XII-A, Marcel coal mine near Rybnik; (C) Medullosa sp. – transverse section of stem fragment with secondary xylem visible, specimen unnumbered, Chwałowice coal mine near Rybnik; (D) Psaronius sp. – transverse section of fragment of inner layer of adventitious roots mantle, specimen XIII-A, Radzionków coal mine. Cuticle dispersae from Upper Carboniferous coals of the Upper Silesian Coal Basin: (E) Small leaf fragment, Jaworzno coal mine, seam 302; (F) indetermined cuticle, Ziemowit coal mine, seam 308. Specimens from (A–D) stored in PC IB UJ; specimens from (E,F) from the S. Florjan collection, stored in Nature Education Centre, Jagiellonian University (all photos by S. Florjan).

Krawczyński et al. (1997) preliminarily, and later Pacyna and Zdebska (2012, with references) in detail documented the first Polish Carboniferous flora preserved within sideritic nodules from the Langsettian stage of Sosnowiec (Upper Silesia) (Figure 6). Superb preservation of the plants within an ironstone concretion was made possible by its quick formation, enabling preservation of the original three-dimensional structure of plant organs such as cones, as well as subtle morphological details. The plants preserved within concretions include rare taxa not known or rare in other modes of preservation, such as lycopsid cones, isolated sporophylls and seed fern pollen organs. The dissimilarities in composition between floras preserved in ironstone concretions and those from surrounding sediments likely result from the concretion formation process, which selectively preserves small, delicate plant organs.
. Studies of Carboniferous Plant Reproductive Organs
As mentioned above, reproductive organs of Carboniferous plants are poorly known in Poland. In the 1930s, Bocheński wrote two papers on lycopod cones that to this day are frequently cited in the literature (e.g., Bateman et al., 1992; Bek, 2017). Bocheński examined the cones of several lycopod species from the Upper Silesian Coal Basin. In addition to standard maceration in Schulze mixture, he also gradually prepared compressions that preserved some tissues. In this way he described a number of previously unknown details and clarified earlier researchers’ doubts about the anatomy of Lepidostrobus major (now Lepidocarpon major), Sigillariostrobus rhombibractiatus, and S. ciliatus (now a synonym of S. rhombibractiatus). Bocheński (1936) described two new species: Sigillariostrobus czarnockii and Lepidostrobus bohdanowiczii (now Lepidocarpon bohdanowiczii or Achlamydocarpon bohdanowiczii). Bocheński (1936, 1939) succeeded in isolating spores from the sporangia of all the species investigated; he illustrated their variability and compared them with dispersed spores. His research constitutes a milestone in research on lycopsids producing seed-like fructifications. His innovative research techniques and excellent photographic documentation of specimens place that work among the classic publications on the topic.
Lipiarski (1965) isolated spores from Langsettian fertile leaves of the fern Pecopteris (Senftenbergia) pennaeformis, described details of the sporangia and spores, and compared spores isolated in situ with dispersed ones. Karczewska (1969) documented microsporangia with saccate pollen of the lyginopteridalean pteridosperm Paracalathiops stachei from the Namurian A of the Chełm I borehole.
In a comprehensive monograph, Pacyna and Zdebska (2010) documented seed fern pollen organs from the Upper Carboniferous of the Lower and Upper Silesian coal basins. Before their research, these organs were very poorly known from the Polish Carboniferous and were mentioned in only a few papers, without appropriate documentation. They proposed a new genus and species of the lyginopteridalean pollen organ: Silesiatheca formosa from sideritic nodules from Sosnowiec (Figure 6A,B). The most diversified were the pollen organs of medullosalean seed ferns. Ten species were documented, including one new genus and seven new species, including single, compound, and aggregated synangia (Figure 6C–E). They confirmed the model for the evolution of medullosalean pollen organs proposed by Taylor et al. (2009) and placed the new taxa described within it. This paper is an important contribution to knowledge of Carboniferous seed fern pollen organs, cited widely in the international literature (e.g., Serbet et al., 2016; Šimůnek, 2020; Zodrow & Mastalerz, 2019; Zodrow & Pšenička, 2021).
Pacyna (2012) critically reviewed studies of Carboniferous and Lower Permian plant reproductive organs from Poland, gave a comprehensive list of taxa published so far, provided a useful dictionary of locality names before and after the border changes of 1945, and clarified the nomenclature and synonymy of taxa described so far from Poland. This is the first such thorough revision of the Palaeozoic flora from Poland.
Figure 6
Seed fern pollen organs preserved within sideritic nodules from the Upper Carboniferous (Langsettian) of Sosnowiec. Lyginopteridalean pollen organ Silesiatheca formosa: (A) Whole pollen organ – Pinnately branching axis bearing terminal stellate synangia, S/100/P5b; (B) Numerous stellate synangia consisted of whorls of four–five ovate longitudinally striated microsporangia, S/100/P5a. Medullosalean pollen organ Boulayatheca ciliata: (C) Negative and positive of pyriform microsynangium, S/100/P4; (D) Enlargement of microsynangium surface, numerous hair bases visible as dark spots, S/100/P7; (E) Enlargement of synangium margin, needle-shaped hairs visible, they are expanded to bases, S/100/P7. All specimens stored in PC IB UJ (photos by S. Florjan and G. Pacyna).

. Permian Floras – The Great Transition From Pteridophyte to Gymnosperm Dominance
Permian (298–251 Ma) macrofloras are rare in Poland (Table 1). The first and still most famous such flora was described by Raciborski (1891) from the Karniowice Travertine (Karniowice – Filipowice area, west of Kraków). Modern research on this flora was done by Lipiarski (1971, 1972a, 1972b), who collected additional fossil material. Based on Permian indicator species, he dated this flora as Earliest Permian. Lipiarski (1972a, 1972b) fully documented Lilpopia, a genus very characteristic for this locality, not only revising the species Lilpopia raciborskii described earlier by Lilpop (1937) but also proposing the new species Lilpopia polonica. Although Permian in age, the Karniowice Travertine flora possesses many wetland-adapted species characteristic for the Carboniferous. They grew in a large oasis fed by underground calcium-rich waters in a rather arid environment. Lipiarski (Lipiarski, 1980; Lipiarski & Sarnecka, 2001) also documented other Early Permian floras from Poland, including from Lower Silesia.
One hundred years ago, Czarnocki and Samsonowicz (Czarnocki, 1923; Czarnocki & Samsonowicz, 1913) preliminarily documented the only Polish Late Permian flora, from Kajetanów in the Holy Cross Mountains. This is a very interesting flora dominated by advanced voltzialean conifers, and peltaspermalean seed ferns; it requires a modern revision.
. Summary
Although access to Ordovician, Silurian, and Devonian rock is meagre in Poland (sparse exposures, scant data from cores), and only some of it contains identifiable plant remains, the results of work on this material, especially Zdebska’s work on Early Devonian fossils, are significant and have entered world science.
Carboniferous strata have provided a huge amount of plant fossils, but the research opportunities offered by this material have been poorly exploited so far. Mainly biostratigraphic research has been done; biologically oriented papers are rare. Statistical analyses of floras and palaeoecological papers are lacking. Research on Carboniferous floras has almost stopped in Poland recently, in line with the global palaeobotanical trend. Rich collections of Polish Carboniferous plants await modern revisions and interpretation.
Data on Polish Permian floras are sparse as a result of the rarity of fossils from outcrops and cores. They also need modern revision. Permian floras are being researched frequently around the world recently, as they document floras changing as a result of aridification and increases of CO2 level.
