. Introduction
Grasslands represent one of the most extensive and diverse formations in the world, yet they are underestimated and poorly studied (García-Mijangos et al., 2021). Alpine meadows and pastures are known to be among the most distinctive and species-rich communities, especially in mountainous landscapes (Li et al., 2020). Semi-arid grasslands even hold world records for vascular plant species richness at spatial scales of 10 to 50 m2, exceeding 100 vascular plant species in 25 m2 plots in some areas of Central and Eastern Europe (Chytrý et al., 2015; Wilson et al., 2012). In the floristically very rich area of Middle Asia, grasslands, especially those located high in the mountains, are of great conservation value but also economically important. Montane and alpine, semi-dry, and mesic meadows are considered to be among the most species-rich phytocoenoses in Middle Asia (Afanasjev, 1956; Safarov, 2018). Thus, alpine meadows and pastures have attracted the attention of botanists for many decades.
One of the first comprehensive typology of the grasslands of western Tian Shan was proposed by Golovkova (1959). It has identified three groups: tall-grass meadows, grasslands of the subalpine belt, and the alpine short-grass swards. Additionally, in the northwestern part of Kyrgyzstan, the Kobresia mats dominated by K. capillifolia have been distinguished. More recently, although in a restricted area of the upper parts of the Arslanbob Valley in Ferghana Mts., German botanists have conducted vegetation studies on grassland communities (Borchardt et al., 2011). They distinguished tall-forb with Aconogonon coriarium and Prangos pabularia and subalpine meadow with Geranium collinum and Phlomoides oreophila. Additionally, in the work of Wagner (2009), some insights into steppe and forb-meadow vegetation in the Aksu-Jabagly Nature Reserve in western Tian Shan were published. It reveals nine distinct plant communities, among others Dactylis glomerata-Karatavia kultiassovii and Nepeta mariae-Aconogonon coriarium which seems to some extent be similar to alpine meadows despite the data collected in the montane belt (1740–2390 m a.s.l.). In recent years, interesting works from the Tuva region of the Altai Mountains have also been completed (Ermakov & Zibzeev, 2012; Zibzeev, 2008; Zibzeev & Nedovesova, 2015). They presented a fairly consistent classification of many vegetation types at the class level and including them mostly in large Euro-Siberian units such as Juncetea trifidi, Salicetea herbaceae and Kobresietea bellardii. Worth noting is the order Festucetalia krylovianae, which captures alpine meadows in the Altai Mountains (Ermakov & Zibzeev, 2012) and despite being composed mainly of Euro-Siberian species, is quite similar to alpine meadows of the western Tian Shan.
Still, the sound classification of grassland communities for the entire Middle Asian area poses considerable difficulties. This is due to the fact that major phytogeographical divisions (e.g. between Middle and Central Asia) pass through this territory, but also because there is a lack of reliable phytosociological data from the whole region to compare vegetation samples. In addition, the high habitat diversity and the close contact between the dominant steppes, lowland meadows (of Eurosiberian character), forb steppes of the subalpine belt (with a dominance of Irano-Turanian flora), alpine meadows, and tall-forbs of the class Prangetea ulopterae (Nowak et al., 2016, 2018, 2020, 2022, 2023) in the landscape make it difficult to establish a comprehensive classification, especially at a high-rank subdivision. The problem of proper classification of Middle Asian grasslands also stems from the fact that neighboring territories (Tibet, southern Siberia, or Persia) do not have a sound, based on modern phytosociological methods, vegetation typology either (see Nowak & Nowak, 2022). Zohary (1973), in his work on the vegetation of the Middle East, describes this vegetation type as “malacophilous subalpine and alpine steppes” and only mentions in general alpine and nival meadows without any proposal for higher syntaxonomic division. In the former Soviet Union, including Middle Asian countries, there is a long tradition of studying vegetation by analyzing the composition of dominant species. The recent works do not refer to detailed investigations of the Middle Asian grasslands (e.g. Korotkov et al., 1991), although they are floristically related to the meadows of nemoral Eurasia (e.g. Rubtsov, 1955; Vykhodtsev, 1956). Particularly grasslands in central Tian Shan are considered to be very similar to Altai meadows (Golovkova, 1959). It is also worth noting that some structural and spatial similarities between dry meadows in the subalpine belt of Middle Asia, which occur within transitional forest zone and just above the timberline, and steppe meadows occurring north of the forest-steppe zone in boreal regions, such as in the southern Urals (e.g. Didukh & Korotchenko, 1996; Vasilevich & Bibikova, 2008).
The subalpine and alpine grasslands of Middle Asia were defined in very different ways. It was recognized as a subalpine meadow of Calamagrostis spp. (beynikovye luga) that were reported from Hissar, Vahsh, Karateginian, and Hazratishokh ranges at 2,500–2,800 m a.s.l. (Safarov, 2018). The Geranium regelii and Nepeta podostachys were indicated as codominant species in these communities. In the western Pamir and Darvaz ranges, semi-dry grasslands are identified as the group of Alopecurus seravschanicus-Hordeum turkestanicum meadows. Both types were assigned to so-called cryophytic meadows (Megagramihyion holarcticum; Safarov, 2018). However, this vegetation type by Ovchinnikov (1948, 1957) and Stanyukovich (1960) is included in tall-forbs as “krupnotravnye luga” (supposedly Prangetea ulopterae in today’s classification). Additionally, in Middle Asia, a region with a Mediterran-type climate and intensive grazing, a distinct formation of the so-called meadow-steppe (lugostepy or ostepennye luga) has also been distinguished. This vegetation type includes phytocenoses with a dominance of Adonis turkestanicus, Artemisia dracunculus and A. pamirica (Afanasjev, 1956; Safarov, 2018). They are related to more humid habitats, although they are able to tolerate drought in the topsoil. However, they are not adapted to extremely low temperatures.
Given the vital importance of a good understanding and proper classification of Middle Asian grassland communities, in 2017, we began surveys in Tajikistan and Kyrgyzstan in all grassland and pasture types. In total, we collected more than 750 samples in different habitat types. We divided the database along the main lines of demarcation, obtaining three major units: a subalpine and alpine belt of the Pamir-Alai (mainly Tajikistan) and Tian Shan (mainly Kyrgyzstan), and the upper alpine and subnival belts of the entire study area. In this paper, we present the classification of the subalpine and alpine grasslands in the Central and Western Tian Shan. We aimed at addressing the following questions during our study: (1) What is the diversity of alpine grasslands of the subalpine and alpine belts in the south-western Tian Shan Mountains? (2) What are the environmental and habitat requirements of the described plant communities? (3) What is the species composition and structure of this vegetation? and (4) Which species have important diagnostic value for a particular community? Additionally, we are aiming at comparing alpine grasslands to montane grasslands studied by Wagner (2009).
. Methods
. Study area
A vegetation survey was conducted in the central part of Middle Asia within the administrative boundaries of Kyrgyzstan in an area of about 57,000 km2 (Figure 1). Study sites covered several mountain ranges of the western Tian Shan, including Talas Ala-Too and Ferghana Mountains in eastern Kyrgyzstan, the Kyrgyz range in the north, At-Bashy Range in the south, the surrounding of an alpine lake Song-Köl and adjacent Songköl Too ridge and Borbor Alabas mountains in central Kyrgyzstan, as well as areas to the south of Issyk-Kul lake in the Terskey Ala-Too in the most eastern parts of the country. The prevailing mountainous character of Kyrgyzstan, with numerous mountain ranges having peaks lying in the alpine and subalpine belts, makes optimal conditions for the development of natural grasslands. However, these areas are not untouched by human activities, as they are typically used by local communities as summer pastures for sheep, goats and, less often, for horses and yaks. Study sites lie within three ecoregions: Tian Shan montane conifer forests, Tian Shan foothill arid steppe, and Tian Shan montane steppe and meadows (Dinerstein et al., 2017).
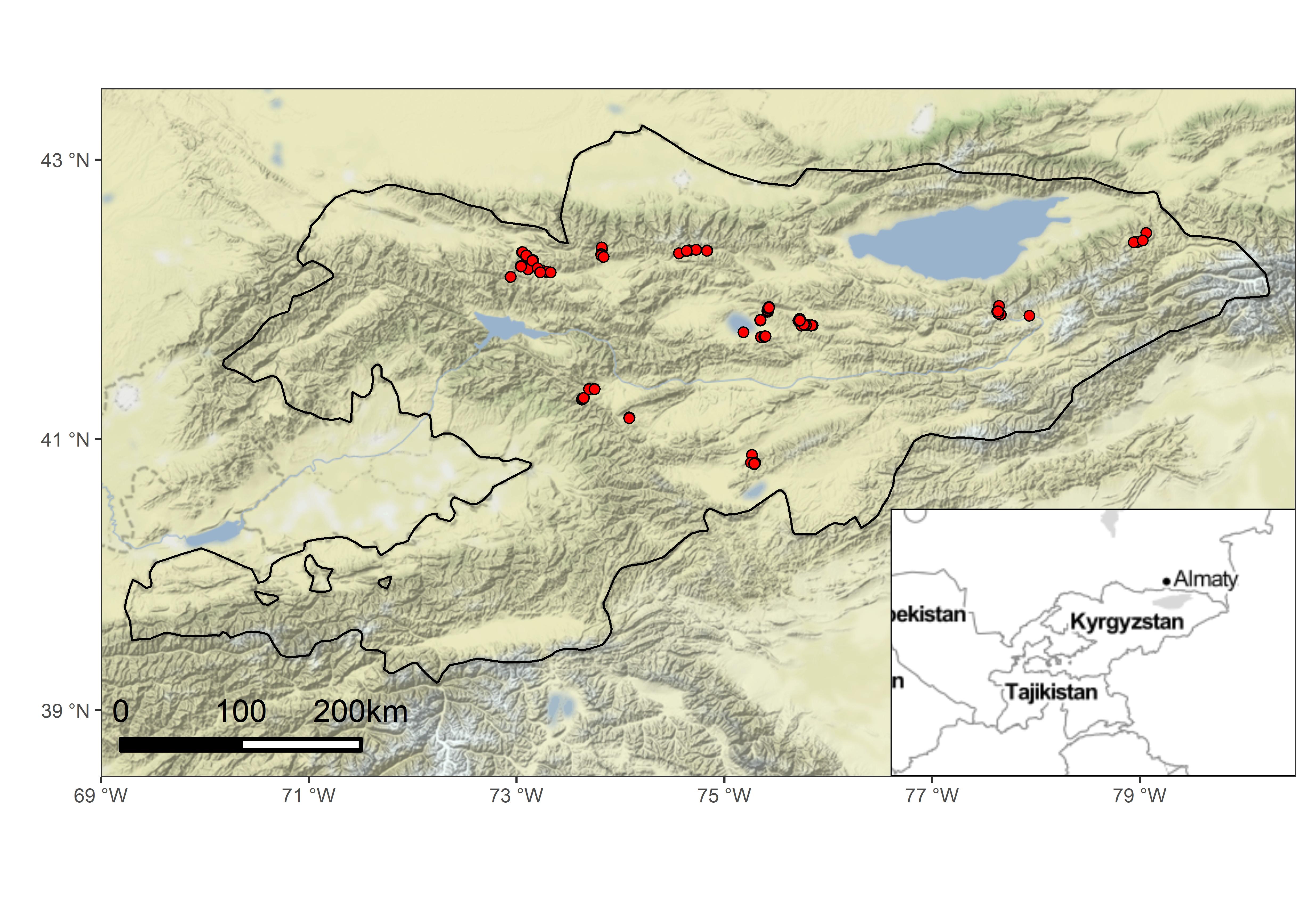
The climatic conditions of the study area are difficult to generalize. According to Djamali et al. (2012), the Irano-Turanian type of bioclimate is prevailing in Kyrgyzstan. This type of bioclimate shows a great spatial variation in winter versus summer precipitation, depending on continentality and elevation. Areas covered by our study belong to subregions with an overall dry climate characterized by extremely high continentality and the highest precipitation in summer. The study area is, however, under the strong influence of the Euro-Siberian bioclimatic type from the north and the Central-Asiatic type from the east. Moreover, climatic conditions among different mountain ranges vary and depend on multiple factors such as altitude, aspect, and inclination of slopes. The studied sites were located in a wide altitudinal gradient from about 1,900 to 3,700 m a.s.l. within differentiated relief (aspect, inclination) and bedrock types above the coniferous forest belt, which is present in many mountain ranges. Up to 2,000–2,200 m a.s.l. temperate climate dominates with warm summers and cold, snowy winters. At higher altitudes, up to 3,000–3,500 m a.s.l. Summer is much cooler, whereas winters are almost snowless.
. Data sampling and data analyses
A total of 251 plots (phytosociological relevés) were sampled in subalpine and alpine grasslands across Kyrgyzstan in remote but accessible mountain ranges of the western Tian Shan Mountains (Figure 1). A field survey was conducted in the years 2015–2022, but most of the data (90%) were collected in 2022. We used a standardized plot size of 10 m2 each. Vascular plant species and bryophytes were recorded using the Braun-Blanquet cover-abundance scale (Westhoff & van der Maarel, 1973). Geographical coordinates were measured using a GPSMAP 60CSx device with an accuracy of about 5 m and a WGS84 reference frame. Plots were stored in the Vegetation of Middle Asia database registered in the Global Index of Vegetation-Plot Databases under the code AS-00-003 (Nowak et al., 2017).
Prior to the classification, the Braun-Blanquet cover-abundance scale was transformed into percentage values as follows: r (0.1%), + (2%), 1 (3%), 2, (13%), 3 (38%), 4 (63%) and 5 (88%). We used the agglomerative hierarchical Ward’s method (Jongman et al., 1997; Podani, 2000) in the PC-ORD program ver. 7.08 (McCune & Mefford, 2011) employing the Euclidean distance measure. The classification level of 13 clusters was chosen subjectively based on field experience and ecological interpretation of delimited clusters after inspecting variants with a different number of clusters from 10 to 20 groups. The resulting groups were interpreted at the level of associations or left rankless as a plant community and then were characterized in terms of species richness, the cover of herb and bryophyte layers, and species composition. The shortened synoptic table was created and presented in the description of each syntaxa. Delimited syntaxa were characterized by diagnostic, constant, and dominant species. Diagnostic species were determined using the phi coefficient as a fidelity measure (Chytrý & Tichý, 2003) following their statistical validation by Fisher exact test (p < 0.01). Values of phi were multiplied by 100 for the presentation of the results in shortened synoptic tables. The fidelity of diagnostic species was determined each time after standardizing the site group to equal size (Tichý & Chytrý, 2006). We considered diagnostic species for a cluster if that species had a phi value ≥ 20 on a 0–100 scale and frequency ≥ 20%. Species with their relative frequency in a given cluster higher than 30% were considered constant for that cluster. Dominant species were those that had at least 25% cover in 5% of the plots belonging to the cluster. To check the floristic-sociological groupings and highlight the relationships between plots and species, non-metric multidimensional scaling (NMDS) was performed (with down-weighting of rare species, response data were log-transformed). It was computed using the ‘vegan’ package version 2.5.4 (Oksanen et al., 2019) in R version 4.2.2 (R Core Team, 2022). For ecological interpretation of the ordination axes, bioclimatic variables, and soil characteristics were plotted onto an NMDS ordination diagram as supplementary variables. Climatic data were extracted from the CHELSA database version 2.1 (http://chelsa-climate.org; Karger et al., 2017), and soil data were extracted from SoilGrids version 2.0 (https://soilgrids.org/; Poggio et al., 2021). Prior to analysis, correlations between 19 bioclimatic and 9 physical and chemical soil variables were calculated using the Spearman correlation coefficient to reduce the number of available variables. Variables indicating strong multicollinearity (r > 0.7 in all pairwise comparisons) were removed, and we retained variables most clearly interpretable from an ecological point of view. The climatic variables used for analysis were: mean annual temperature, mean diurnal temperature range, temperature seasonality, and annual precipitation. The soil parameters were: clay content, silt, sand, coarse fragments, bulk density, pH, cation exchange capacity, organic carbon, and nitrogen.
A shortened synoptic table with the fidelity and relative percentage frequency of all diagnostic species is shown in Table 1. Analytic table, including type relevés (Table S1) and a full synoptic table (Table S2), are available in the Figshare Digital Repository (https://doi.org/10.6084/m9.figshare.22586293).
Table 1
Shortened synoptic table with percentage frequency and fidelity (superscript) values. Only diagnostic species are given. See supplementary material 2 for the full version of this table (Table S2). Group No.: 1.1.1. Potentillo pamiroalaicae-Pedicularidetum allorramphae; 1.1.2. Scutellario oligodontae-Hedysaretum narynense; 1.1.3. Aulacospermo tianschanici-Pulsatilletum campanulae; 1.1.4. Seselido tallasici-Caricetum alajicae; 1.1.5. Poo alpinae-Eritrichietum villosae; 1.1.6. Oxytropis globifera-Leontopodium ochroleucum community; 1.1.7. Kobresio capillifoliae-Hedysaretum kirghisori; 1.1.8. Seselideto valentinae-Oxytropidetum atbaschi; 1.1.9. Caraganetum jubatae; 1.1.10. Caraganetum aurantiacae; 2.1.1. Potentillo moorcroftii-Stipetum tremulae; 2.1.2. Ptilagrostio malyschevii-Trisetetum altaici; 3.1.1. Onobrychideto arenariae-Stachyetum betoniciformae.
| Group | 1.1.1 | 1.1.2 | 1.1.3 | 1.1.4 | 1.1.5 | 1.1.6 | 1.1.7 | 1.1.8 | 1.1.9 | 1.1.10 | 2.1.1 | 2.1.2 | 3.1.1 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of relevés | 34 | 8 | 9 | 36 | 25 | 21 | 23 | 7 | 12 | 5 | 28 | 10 | 33 | |||||||||||||
| Potentillo pamiroalaicae-Pedicularidetum allorramphae | ||||||||||||||||||||||||||
| Alchemilla tianschanica | 50 | 32 | . | 11 | 33 | 36 | 5 | 4 | . | 8 | . | 18 | . | . | ||||||||||||
| Agrostis turkestanica | 35 | 30 | . | 33 | 8 | 24 | . | . | . | . | . | . | . | . | ||||||||||||
| Galium turkestanicum | 38 | 27 | . | 33 | 19 | 12 | . | . | . | . | . | . | . | 27 | ||||||||||||
| Trifolium repens | 26 | 25 | . | 11 | 11 | 16 | . | . | . | 8 | . | . | . | 3 | ||||||||||||
| Pedicularis allorrhampha | 29 | 25 | . | 11 | 14 | 8 | 5 | . | . | . | . | . | . | 24 | ||||||||||||
| Cerastium tianschanicum | 32 | 23 | . | 11 | 19 | . | . | 13 | . | 25 | . | . | . | 18 | ||||||||||||
| Scutellario oligodontae-Hedysaretum narynense | ||||||||||||||||||||||||||
| Hedysarum narynense | . | 100 | 100 | . | . | . | . | . | . | . | . | . | . | . | ||||||||||||
| Dracocephalum nodulosum | . | 88 | 88 | . | . | . | . | . | . | . | . | . | . | 9 | ||||||||||||
| Dracocephalum discolor | . | 88 | 71 | 11 | 11 | . | 10 | . | . | . | . | 18 | . | 3 | ||||||||||||
| Scutellaria oligodonta | . | 88 | 69 | 11 | 19 | . | 19 | . | . | 8 | . | . | . | 3 | ||||||||||||
| Potentilla nervosa | 21 | 100 | 62 | 33 | 33 | 12 | 24 | . | . | . | 8 | . | 4 | . | 6 | |||||||||||
| Thesium alatavicum | . | 50 | 58 | 11 | . | . | 5 | . | . | . | . | . | . | 3 | ||||||||||||
| Pyrethrum karelinii | . | 63 | 56 | 11 | 3 | 16 | 5 | . | . | 8 | . | . | . | 6 | ||||||||||||
| Carex stenophylla subsp. stenophylloides | 3 | 50 | 55 | . | . | . | . | . | 14 | 8 | . | . | . | . | ||||||||||||
| Pedicularis ludwigii | 6 | 88 | 53 | . | 25 | . | 33 | 13 | . | 33 | . | 7 | . | 21 | ||||||||||||
| Elytrigia gmelinii | 15 | 50 | 52 | . | 8 | . | . | . | . | 8 | . | . | . | . | ||||||||||||
| Plantago arachnoidea | . | 63 | 51 | 22 | 3 | 4 | 14 | 4 | . | 8 | . | 11 | . | . | ||||||||||||
| Ligularia knorringiana | 3 | 75 | 48 | 11 | 25 | . | 19 | . | . | 25 | . | 21 | . | 21 | ||||||||||||
| Aster alpinus var. serpentimontanus | 9 | 100 | 46 | 22 | 6 | 32 | 43 | 26 | . | 33 | . | 46 | 12 | 30 | 15 | |||||||||||
| Erysimum altaicum | . | 25 | 46 | . | 3 | . | . | . | . | . | . | . | . | . | ||||||||||||
| Bupleurum densiflorum | 6 | 75 | 44 | 33 | 11 | . | 29 | . | . | 17 | . | 29 | 20 | 9 | ||||||||||||
| Cirsium sieversii | 6 | 50 | 39 | 11 | 19 | . | 19 | 9 | . | . | . | . | . | 12 | ||||||||||||
| Hordeum brevisubulatum | . | 63 | 37 | . | 31 | 12 | . | 24 | 13 | . | 42 | . | 18 | 10 | 3 | |||||||||||
| Psathyrostachys kronenburgii | . | 38 | 37 | 11 | 6 | 16 | . | 4 | . | 8 | . | . | . | . | ||||||||||||
| Adenophora himalayana | . | 38 | 33 | 11 | 14 | . | 5 | . | . | 17 | . | . | . | 12 | ||||||||||||
| Potentilla asiae-mediae | 3 | 63 | 26 | 44 | 17 | 4 | 43 | 30 | 29 | 17 | . | 18 | 30 | 18 | ||||||||||||
| Aulacospermo tianschanici-Pulsatilletum campanulae | ||||||||||||||||||||||||||
| Pulsatilla campanella | 35 | 38 | 100 | 48 | 14 | 24 | 52 | 17 | . | 8 | . | 46 | . | 15 | ||||||||||||
| Jurinea lanipes | 3 | . | 22 | 40 | 3 | . | . | . | . | . | . | . | . | . | ||||||||||||
| Helictotrichon desertorum | 18 | 13 | 56 | 39 | 17 | 12 | 10 | . | . | 8 | . | 21 | . | . | ||||||||||||
| Aulacospermum tianschanicum | 24 | 38 | 56 | 32 | 11 | 28 | 10 | 9 | . | 17 | . | 4 | . | 9 | ||||||||||||
| Seselido tallasici-Caricetum alajicae | ||||||||||||||||||||||||||
| Phlomoides oreophila | 53 | 75 | 44 | 100 | 39 | 8 | 29 | 43 | . | 42 | . | 18 | 30 | 24 | ||||||||||||
| Allium hymenorhizum | 12 | . | . | 42 | 34 | 20 | 10 | . | . | 8 | . | . | . | 21 | ||||||||||||
| Seseli talassicum | 9 | 38 | . | 58 | 31 | 12 | 10 | 39 | . | 25 | . | 4 | 20 | 12 | ||||||||||||
| Ligularia thomsonii | 9 | . | 11 | 28 | 27 | 8 | . | . | . | . | . | . | . | 21 | ||||||||||||
| Campanula glomerata | 29 | . | 33 | 50 | 23 | 8 | . | 4 | . | 42 | 60 | . | . | 15 | ||||||||||||
| Carex alajica | 21 | 38 | 22 | 50 | 23 | 20 | 5 | 13 | . | 8 | . | 14 | 40 | 12 | ||||||||||||
| Delphinium oreophilum | 3 | 50 | 11 | 36 | 20 | 20 | 19 | 13 | . | 8 | . | . | . | 3 | ||||||||||||
| Poo alpinae-Eritrichietum villosae | ||||||||||||||||||||||||||
| Eritrichium villosum | . | . | . | . | 40 | 53 | . | . | . | 8 | . | . | . | 3 | ||||||||||||
| Festuca alaica | 18 | . | 56 | 11 | 84 | 51 | 5 | 9 | . | 8 | . | 21 | 10 | . | ||||||||||||
| Erigeron allochrous | 24 | 19 | . | . | 3 | 48 | 48 | 10 | . | . | . | . | . | . | 3 | |||||||||||
| Poa alpina | 26 | . | 11 | 3 | 76 | 47 | 5 | 4 | . | 33 | . | 29 | 20 | 3 | ||||||||||||
| Crepis oreades | 6 | 13 | . | 8 | 48 | 45 | 10 | . | . | 8 | . | . | . | 3 | ||||||||||||
| Aconitum rotundifolium | 21 | . | 33 | 11 | 60 | 42 | 14 | 9 | . | . | . | . | 10 | . | ||||||||||||
| Lomatocarpa albomarginata | 3 | . | . | . | 20 | 40 | . | . | . | . | . | . | . | . | ||||||||||||
| Alchemilla sibirica | 9 | . | . | . | 24 | 37 | . | . | . | . | . | 4 | . | . | ||||||||||||
| Cerastium pusillum | 44 | . | 33 | 39 | 72 | 35 | 19 | 22 | 14 | 17 | . | 14 | . | 15 | ||||||||||||
| Dichodon cerastoides | 12 | . | . | . | 20 | 33 | . | . | . | . | . | . | . | . | ||||||||||||
| Eremogone meyeri | 9 | . | 11 | 11 | 36 | 28 | 19 | . | . | . | . | 14 | . | 12 | ||||||||||||
| Koeleria macrantha | 24 | . | . | 25 | 36 | 27 | 19 | . | . | . | . | 4 | . | 9 | ||||||||||||
| Gentianella turkestanorum | 38 | . | 56 | 31 | 64 | 27 | 48 | 9 | . | . | . | 11 | 30 | 24 | ||||||||||||
| Bistorta elliptica | 56 | 18 | . | 33 | 53 | 16 | 68 | 26 | . | 30 | . | 58 | . | 4 | 50 | 6 | ||||||||||
| Draba subamplexicaulis | 21 | . | 33 | 14 | 32 | 26 | . | . | . | . | . | 4 | . | . | ||||||||||||
| Erigeron aurantiacus | 3 | . | . | 14 | 24 | 25 | . | . | . | 25 | . | . | . | . | ||||||||||||
| Euphrasia drosophylla | 41 | . | 44 | 17 | 60 | 24 | 43 | 22 | . | . | . | 29 | 50 | 6 | ||||||||||||
| Potentilla hololeuca | 15 | . | . | 6 | 24 | 24 | 5 | 13 | . | . | . | . | . | 6 | ||||||||||||
| Kobresio capillifoliae-Hedysaretum kirghisori | ||||||||||||||||||||||||||
| Kobresia capillifolia | 3 | . | 11 | 22 | 20 | 5 | 100 | 61 | . | . | . | 21 | 50 | 6 | ||||||||||||
| Sibirotrisetum sibiricum | 3 | . | 11 | . | . | . | 30 | 43 | . | . | . | . | . | . | ||||||||||||
| Hedysarum kirghisorum | 3 | . | . | 25 | . | 5 | 39 | 30 | . | 25 | . | 14 | . | 9 | ||||||||||||
| Festuca alatavica | . | . | 11 | 33 | 4 | . | 57 | 30 | . | 42 | . | 29 | 40 | 12 | ||||||||||||
| Myosotis asiatica | 59 | 13 | 44 | 64 | 60 | 19 | 87 | 22 | . | 58 | 80 | 36 | 60 | 48 | ||||||||||||
| Seselideto valentinae-Oxytropidetum atbaschi | ||||||||||||||||||||||||||
| Eremopyrum distans | . | . | . | . | . | . | . | 100 | 100 | . | . | . | . | . | ||||||||||||
| Oxytropis atbaschi | . | . | . | . | . | . | . | 100 | 100 | . | . | . | . | . | ||||||||||||
| Lappula stricta | . | . | . | . | . | . | . | 100 | 98 | . | . | . | . | 3 | ||||||||||||
| Astragalus nivalis | . | . | . | . | 4 | . | . | 86 | 90 | . | . | . | . | . | ||||||||||||
| Stipa caucasica | . | . | . | . | 4 | 10 | . | 86 | 80 | . | . | 11 | . | . | ||||||||||||
| Allium oreoprasum | . | . | . | . | . | . | . | 57 | 74 | . | . | . | . | . | ||||||||||||
| Seseli valentinae | . | 13 | . | . | . | . | . | 71 | 72 | 8 | . | . | . | . | ||||||||||||
| Oxytropis ruebsaamenii | . | . | . | . | . | . | 4 | 57 | 71 | . | . | . | . | . | ||||||||||||
| Stipa subsessiliflora | . | . | . | . | . | . | . | 43 | 64 | . | . | . | . | . | ||||||||||||
| Androsace fedtschenkoi | 18 | . | 33 | . | 8 | 5 | . | 57 | 48 | . | . | . | . | . | ||||||||||||
| Poa relaxa | 3 | . | . | . | . | . | . | 29 | 47 | . | . | . | . | 3 | ||||||||||||
| Hedysarum krylovii | . | . | . | . | . | . | . | 29 | 46 | . | . | . | . | 6 | ||||||||||||
| Goniolimon orthocladum | . | . | . | . | . | . | . | 29 | 44 | . | . | . | . | 9 | ||||||||||||
| Artemisia viridis | 12 | 50 | 44 | 14 | 12 | 43 | 30 | 86 | 38 | 8 | . | 14 | 30 | 6 | ||||||||||||
| Caraganetum jubatae | ||||||||||||||||||||||||||
| Caragana jubata (sl) | . | . | . | . | . | . | . | . | 50 | 69 | . | . | . | . | ||||||||||||
| Sanguisorba officinalis | . | . | . | . | . | . | . | . | 42 | 63 | . | . | . | . | ||||||||||||
| Juniperus sabina (sl) | . | . | . | . | . | . | . | . | 33 | 53 | . | . | . | 3 | ||||||||||||
| Carex regeliana | . | . | . | . | . | . | . | . | 33 | 53 | . | . | . | 3 | ||||||||||||
| Phlomoides pratensis | 6 | . | . | . | . | . | . | . | 42 | 49 | . | 7 | . | 9 | ||||||||||||
| Scrophularia griffithii | . | . | . | . | . | . | . | . | 25 | 45 | . | . | . | 3 | ||||||||||||
| Carex melanantha | . | . | . | . | . | . | . | . | 25 | 45 | . | . | . | 3 | ||||||||||||
| Aconitum nemorum | . | . | . | 6 | . | . | . | . | 25 | 43 | . | . | . | . | ||||||||||||
| Festuca pratensis | 3 | . | . | 6 | . | . | . | . | 25 | 41 | . | . | . | . | ||||||||||||
| Codonopsis clematidea | 3 | . | . | 28 | 18 | . | 5 | . | . | 50 | 41 | 20 | . | . | 15 | |||||||||||
| Elymus longe-aristatus | . | . | 11 | . | . | 5 | 9 | . | 42 | 38 | . | 21 | 16 | . | 6 | |||||||||||
| Pedicularis oederi | . | . | . | . | . | . | . | . | 25 | 37 | . | 14 | 19 | . | . | |||||||||||
| Astragalus lepsensis | . | . | . | . | . | . | 9 | . | 25 | 35 | . | 7 | . | 3 | ||||||||||||
| Polygonatum roseum | . | . | . | . | . | . | . | . | 25 | 34 | 20 | . | . | . | ||||||||||||
| Aconogonon alpinum | 3 | . | . | 11 | . | 5 | . | . | 25 | 33 | . | 4 | . | . | ||||||||||||
| Lonicera semenovii | 6 | . | 11 | 3 | . | . | 9 | . | 25 | 29 | . | . | . | 3 | ||||||||||||
| Geranium saxatile | 82 | 16 | 63 | 78 | 83 | 16 | 84 | 17 | 43 | 35 | . | 100 | 26 | 100 | 29 | . | 21 | |||||||||
| Caraganetum aurantiacae | ||||||||||||||||||||||||||
| Philonotis fontana (ml) | . | . | . | . | . | . | . | . | . | 100 | 100 | . | . | . | ||||||||||||
| Caragana aurantiaca (sl) | . | . | . | . | . | . | . | . | . | 100 | 100 | . | . | . | ||||||||||||
| Seseli mucronatum | . | . | . | 3 | . | . | . | . | 8 | 100 | 94 | . | . | . | ||||||||||||
| Cerastium falcatum | . | . | . | . | . | . | . | . | . | 80 | 89 | . | . | . | ||||||||||||
| Hygroamblystegium varium (ml) | . | . | . | . | . | . | . | . | . | 80 | 89 | . | . | . | ||||||||||||
| Bryum pseudotriquetrum (ml) | . | . | . | . | . | . | . | . | . | 80 | 87 | . | . | 3 | ||||||||||||
| Aconitum soongaricum | . | . | . | . | . | . | . | . | . | 60 | 76 | . | . | . | ||||||||||||
| Rumex paulsenianus | . | . | . | . | . | . | . | . | 8 | 60 | 71 | . | . | . | ||||||||||||
| Galium tianschanicum | 3 | . | 11 | . | . | . | . | . | . | 60 | 68 | . | . | . | ||||||||||||
| Aconitum leucostomum | . | . | . | . | . | . | . | . | . | 40 | 62 | . | . | . | ||||||||||||
| Carex karoi | . | . | . | . | . | . | . | . | . | 40 | 62 | . | . | . | ||||||||||||
| Alopecurus pratensis | . | . | . | . | . | . | . | . | . | 40 | 62 | . | . | . | ||||||||||||
| Ligularia heterophylla | 3 | . | . | 6 | . | . | . | . | 17 | 60 | 60 | . | . | 6 | ||||||||||||
| Allium fedtschenkoanum | 9 | . | . | . | 12 | . | 9 | . | . | 60 | 54 | . | 20 | . | ||||||||||||
| Carduus nutans | . | . | . | 6 | . | 10 | . | . | . | 40 | 51 | . | . | . | ||||||||||||
| Potentillo moorcroftii-Stipetum purpureae | ||||||||||||||||||||||||||
| Plantago griffithii | 6 | . | . | . | . | 10 | . | . | . | . | 29 | 40 | . | . | ||||||||||||
| Potentilla moorcroftii | . | 13 | 11 | . | 8 | 24 | . | . | 8 | . | 43 | 35 | . | 6 | ||||||||||||
| Stipa tremula* | . | . | . | . | . | . | . | . | 8 | . | 18 | 33 | . | . | ||||||||||||
| Stipa regeliana | . | . | . | . | . | 19 | 22 | . | . | . | 25 | 25 | . | 6 | ||||||||||||
| Thalictrum alpinum | 3 | . | . | 3 | . | 5 | 17 | . | . | . | 25 | 21 | 30 | 3 | ||||||||||||
| Ptilagrostio malyschevii-Trisetetum altaici | ||||||||||||||||||||||||||
| Tulipa dasystemon | 9 | . | . | 6 | 12 | . | 13 | . | 8 | . | . | 90 | 73 | 6 | ||||||||||||
| Trisetum altaicum | 3 | . | 11 | 3 | . | 10 | 13 | . | . | . | 11 | 90 | 72 | 6 | ||||||||||||
| Swertia graciliflora | . | . | . | . | . | . | . | . | . | . | . | 20 | 40 | 3 | ||||||||||||
| Artemisia aschurbajewii | 35 | 18 | . | 22 | 8 | 20 | 5 | 17 | . | 8 | . | . | 60 | 38 | 6 | |||||||||||
| Ranunculus rubrocalyx | . | . | 11 | 3 | . | 10 | 13 | . | 8 | . | 7 | 40 | 37 | . | ||||||||||||
| Carex stenocarpa | 3 | . | . | . | 4 | . | 4 | . | 8 | . | 7 | 30 | 35 | 3 | ||||||||||||
| Gastrolychnis uralensis | 9 | . | 11 | 25 | 52 | 19 | 29 | 48 | . | 25 | . | 25 | 70 | 31 | 18 | |||||||||||
| Gentiana kirilowii | 26 | 25 | 67 | 31 | 24 | 24 | 9 | . | 17 | . | 50 | 14 | 70 | 27 | 24 | |||||||||||
| Onobrychideto arenariae-Stachyetum betoniciformae | ||||||||||||||||||||||||||
| Stachys betoniciflora | . | . | . | . | . | . | . | . | . | . | . | . | 24 | 48 | ||||||||||||
| Geranium regelii | 3 | . | . | 11 | . | 10 | 17 | . | . | . | . | . | 48 | 47 | ||||||||||||
| Onobrychis arenaria | . | . | . | . | . | . | . | . | . | . | . | . | 21 | 45 | ||||||||||||
| Stipa capillata | . | . | . | . | . | . | . | . | . | . | . | . | 21 | 45 | ||||||||||||
| Potentilla asiatica | 3 | . | . | 8 | . | . | . | . | . | . | . | . | 27 | 41 | ||||||||||||
| Bromus inermis | 6 | . | . | . | 4 | 14 | . | . | . | . | . | . | 30 | 38 | ||||||||||||
| Carex turkestanica | 9 | . | . | 19 | . | 24 | . | . | 17 | . | . | . | 42 | 35 | ||||||||||||
| Phleum pratense | 3 | . | 11 | 8 | . | . | 4 | . | . | 40 | . | . | 33 | 28 | ||||||||||||
| Festuca valesiaca | 6 | 13 | 33 | 6 | . | 19 | 4 | 29 | . | . | . | . | 42 | 28 | ||||||||||||
| Achillea millefolium | 18 | . | 11 | 11 | . | . | . | . | . | . | . | . | 24 | 26 | ||||||||||||
| Artemisia dracunculus | 6 | . | 22 | 6 | . | 19 | . | . | . | 40 | 4 | . | 36 | 25 | ||||||||||||
| Galium pamiroalaicum | 50 | 50 | 33 | 61 | 17 | . | 29 | 26 | . | 8 | 40 | 18 | 50 | 70 | 22 | |||||||||||
| Species diagnostic for more than two units | ||||||||||||||||||||||||||
| Potentilla pamiroalaica | 79 | 40 | 13 | 33 | 22 | 68 | 32 | 10 | . | 43 | 8 | . | . | . | 9 | |||||||||||
| Thymus incertus | 44 | 31 | . | 56 | 42 | 8 | 16 | 5 | . | . | . | . | . | . | 9 | |||||||||||
| Dracocephalum nutans | 29 | 27 | . | . | . | 44 | 45 | . | . | . | 8 | . | . | . | . | |||||||||||
| Astragalus alpinus | 41 | 25 | . | . | 36 | 21 | 8 | 33 | 4 | . | 17 | . | 7 | . | 15 | |||||||||||
| Primula algida | 50 | 24 | . | 33 | 14 | 88 | 52 | . | 9 | . | 25 | . | 11 | . | 6 | |||||||||||
| Trisetum spicatum | 41 | 23 | 25 | 22 | 11 | 44 | 25 | 10 | 17 | . | . | . | 7 | . | 3 | |||||||||||
| Minuartia biflora | 26 | 23 | . | 11 | 3 | 48 | 47 | . | . | . | . | . | . | . | . | |||||||||||
| Pedicularis korolkowii | 47 | 20 | . | 44 | 19 | 72 | 39 | 14 | 9 | . | . | . | . | 40 | 3 | |||||||||||
| Helictotrichon hookeri | 24 | 100 | 47 | 22 | 22 | 4 | 43 | 13 | . | 33 | . | 11 | 70 | 28 | 9 | |||||||||||
| Androsace lactiflora | . | 50 | 46 | . | . | . | 29 | 23 | . | . | . | . | 14 | . | 6 | |||||||||||
| Thalictrum simplex | 18 | 88 | 44 | . | 50 | 18 | . | 14 | 4 | . | 17 | 100 | 52 | . | . | 12 | ||||||||||
| Elymus czimganicus | 12 | 75 | 40 | 56 | 26 | 33 | 10 | 4 | 19 | 4 | . | 17 | . | 21 | 10 | 6 | ||||||||||
| Festuca amblyodes | 38 | 88 | 31 | 33 | 28 | 36 | 48 | 52 | . | 17 | . | 39 | 80 | 26 | 12 | |||||||||||
| Oxytropis melanotricha | 26 | . | 67 | 45 | 3 | 60 | 39 | 10 | 4 | . | . | . | . | . | 6 | |||||||||||
| Poa litvinoviana | 38 | . | 67 | 28 | 11 | 36 | 14 | 22 | . | 25 | . | 18 | 90 | 44 | . | |||||||||||
| Lamium album | 6 | . | . | 31 | 26 | . | . | . | . | 50 | 48 | . | . | . | 6 | |||||||||||
| Trollius dschungaricus | 12 | . | . | 39 | 22 | 24 | 5 | 9 | . | 17 | . | . | 60 | 41 | 3 | |||||||||||
| Rumex acetosa | 21 | . | 11 | 36 | 21 | 52 | 35 | . | 4 | . | 17 | . | . | 10 | 9 | |||||||||||
| Carex orbicularis | 15 | . | . | 6 | 48 | 32 | . | . | . | . | 80 | 61 | 7 | . | . | |||||||||||
| Papaver croceum | 32 | . | 11 | 17 | 56 | 31 | . | 13 | . | 58 | 33 | . | 4 | . | 18 | |||||||||||
| Leontopodium ochroleucum | 15 | 88 | 44 | 25 | 52 | 81 | 22 | 52 | . | 33 | . | 68 | 14 | 90 | 27 | 21 | ||||||||||
| Potentilla tephroleuca | . | . | 11 | 19 | 4 | 48 | 20 | 35 | . | 8 | . | 39 | 14 | 90 | 51 | . | ||||||||||
| Oxytropis globiflora | 21 | 50 | 44 | 36 | 16 | 71 | 20 | 65 | 16 | . | 17 | . | 57 | 90 | 31 | 27 | ||||||||||
| Bistorta vivipara | . | . | . | 11 | . | 5 | 52 | 37 | . | 17 | . | 43 | 28 | 20 | 3 | |||||||||||
| Parnassia laxmannii | 12 | . | 11 | 47 | 14 | . | 19 | 65 | 26 | . | 50 | . | 29 | 90 | 42 | 18 | ||||||||||
| Ptilagrostis malyschevii | . | . | . | . | . | . | 39 | 23 | . | . | . | 18 | 100 | 76 | 6 | |||||||||||
| Valeriana ficariifolia | 9 | 13 | . | 3 | . | . | . | . | 50 | 33 | 80 | 59 | 4 | . | 6 | |||||||||||
| Kobresia persica | 29 | 25 | 56 | 6 | 28 | 43 | 13 | . | . | . | 93 | 38 | 100 | 43 | 15 | |||||||||||
| Euphorbia alatavica | 68 | 23 | . | 22 | 22 | 72 | 26 | 24 | 43 | . | 25 | . | 4 | 100 | 43 | 21 | ||||||||||
The nomenclature of the vascular plants follows Plants of the World Online (POWO, 2023), and problematic taxonomic issues were based on The World Flora Online (WFO, 2023). Nomenclature of Stipa spp. follows Nobis et al. (2020, 2022) and Geranium spp. follows Cherepanov (1995). The plant material collected during field studies was deposited in the Herbarium of Middle Asia Mountains, hosted in OPUN (University of Opole, Poland) and KRA (Jagiellonian University, Poland).
. Results
. Classification of the vegetation units
The numerical classification of 251 plots yielded 13 ecologically meaningful clusters corresponding to 12 associations and one rankless unit (Figure 2, Figure 3). Ten associations represent subalpine and alpine mesic forb meadows and pastures of the newly described alliance Aconito rotundifoliae-Potentilion pamiroalaicae. This alliance also included compositionally similar but physiognomically distinct shrubby subalpine chionophilous heaths. We also identified two associations of Kobresia mats from the Carici rupestris-Kobresietea bellardii and assigned them to the newly established alliance Stipo regelianae-Ptilagrostion malyschevii. Also, an association of subalpine forb steppe vegetation was described, and we propose to include it in the new syntaxonomical class Eremogono griffithii-Nepetea podostachys.
Figure 2
Dendrogram for the classification of the vegetation of the alpine grasslands identified by Ward’s method, resulting in 13 groups: 1.1.1. Potentillo pamiroalaicae-Pedicularidetum allorramphae; 1.1.2. Scutellario oligodontae-Hedysaretum narynense; 1.1.3. Aulacospermo tianschanici-Pulsatilletum campanulae; 1.1.4. Seselido tallasici-Caricetum alajicae; 1.1.5. Poo alpinae-Eritrichietum villosae; 1.1.6. Oxytropis globifera-Leontopodium ochroleucum community; 1.1.7. Kobresio capillifoliae-Hedysaretum kirghisori; 1.1.8. Seselideto valentinae-Oxytropidetum atbaschi; 1.1.9. Caraganetum jubatae; 1.1.10. Caraganetum aurantiacae; 2.1.1. Potentillo moorcroftii-Stipetum tremulae; 2.1.2. Ptilagrostio malyschevii-Trisetetum altaici; 3.1.1. Onobrychideto arenariae-Stachyetum betoniciformae. A detailed dendrogram showing the assignment of each relevé to a particular cluster is presented in Supplementary material (Figure S1).
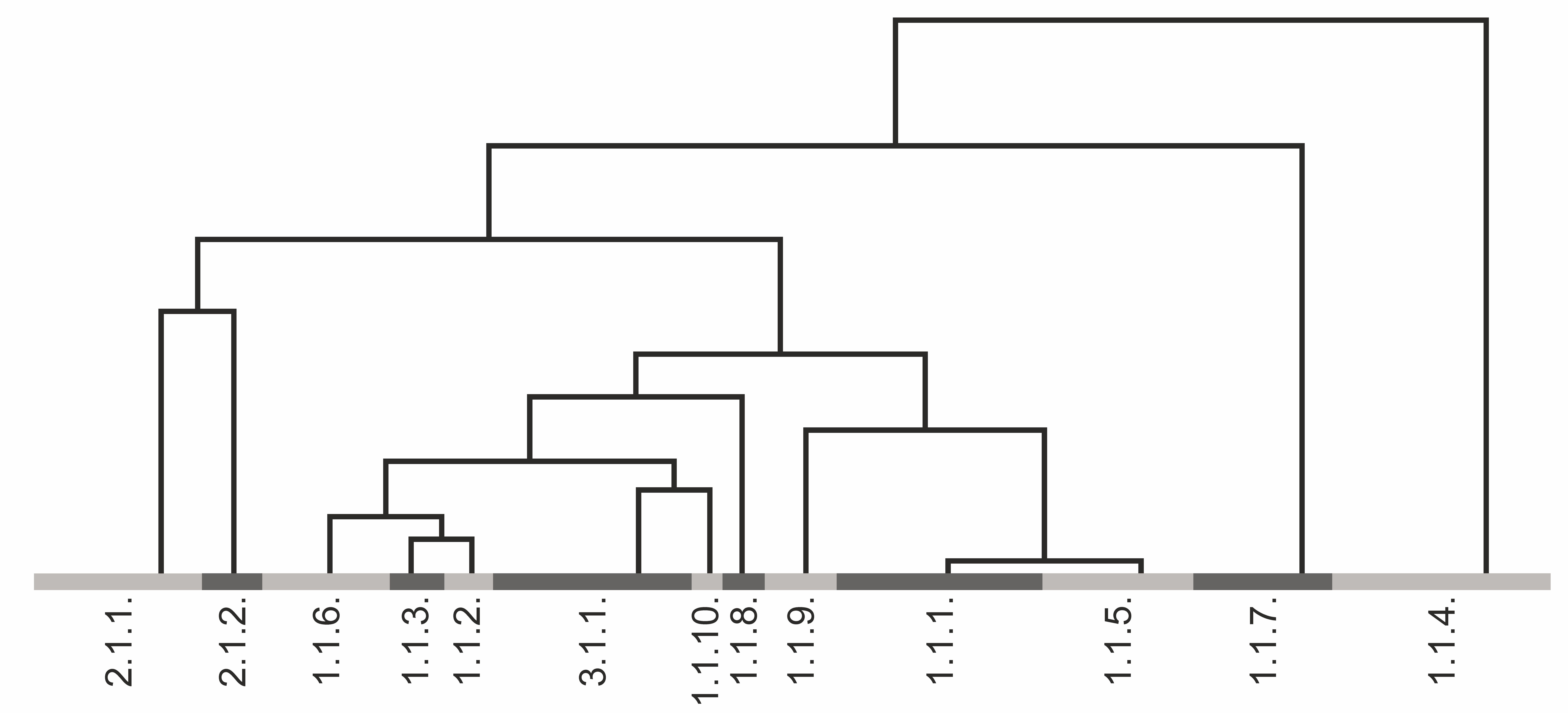
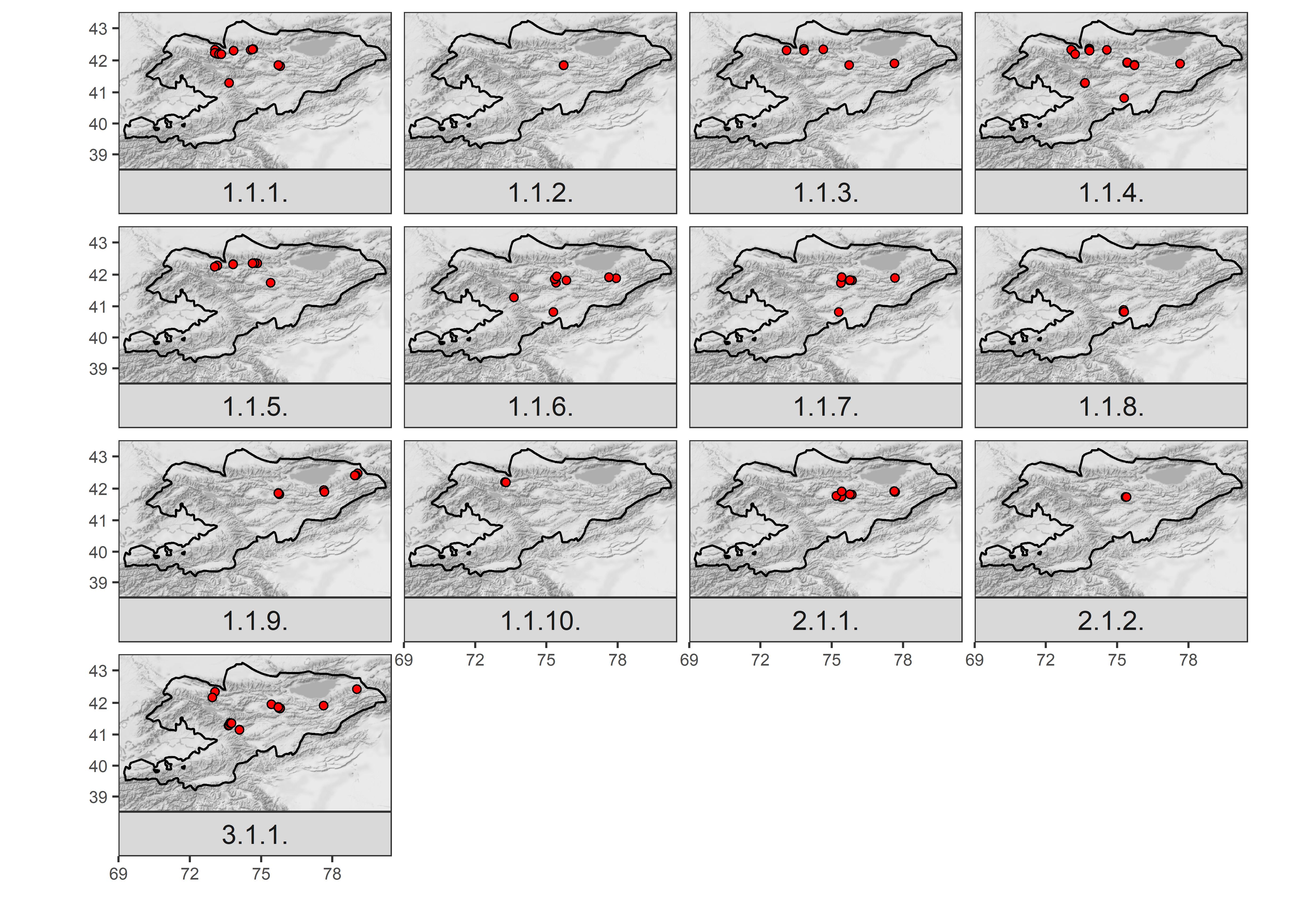
Figure 3
Distribution of plots assigned to the particular vegetation units (n = 251). The names of syntaxa (1.1.1–3.1.1.) are explained in the Syntaxonomic synopsis.
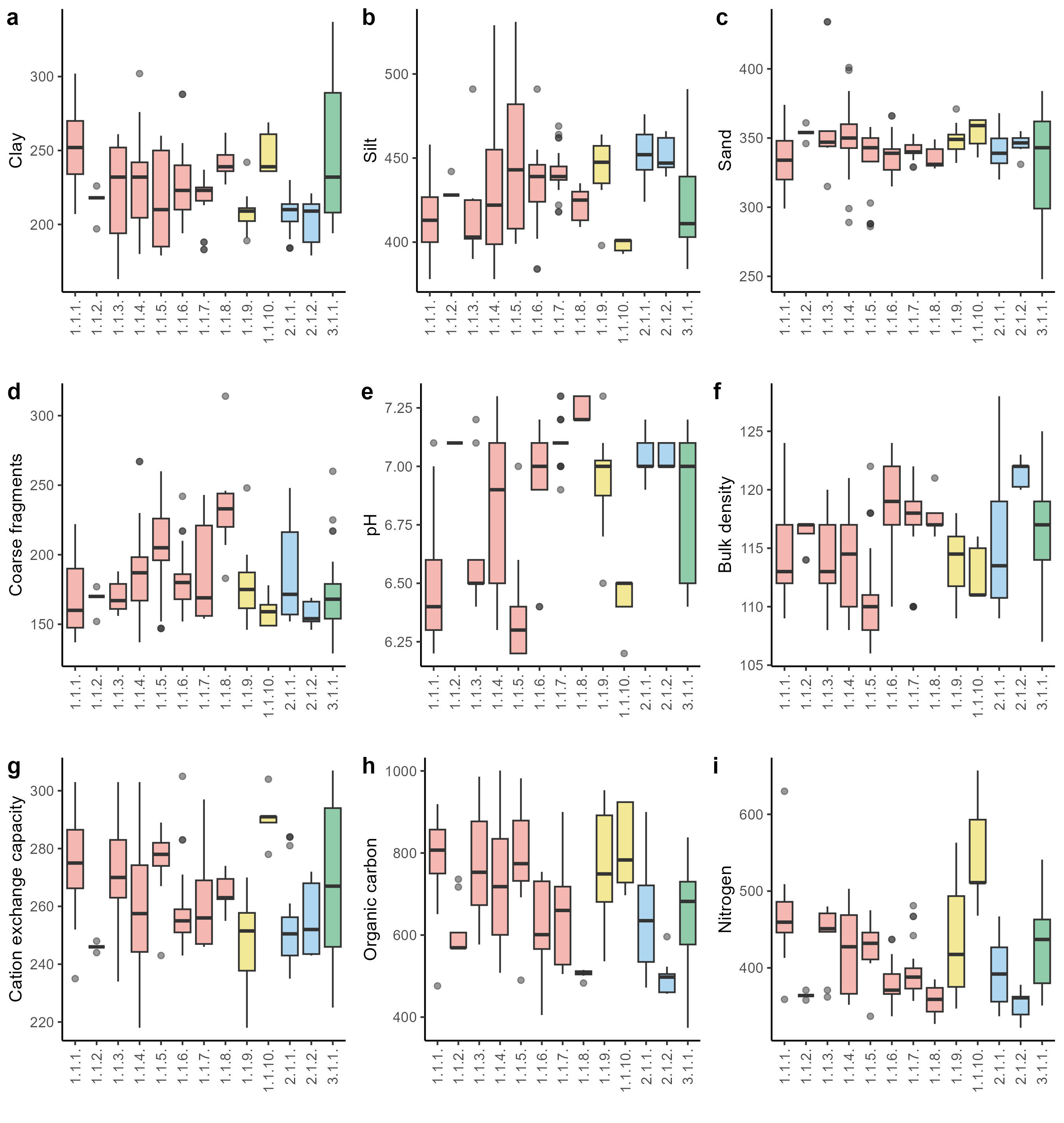
Climatic and soil parameters vectors were fitted against NMDS ordination, and the most important variables (p < 0.01) are shown in the ordination plot (Figure 4). The first axis represents a gradient of temperature seasonality, mean diurnal temperature range, and a sum of annual precipitation (Figure 4a). Furthermore, displays a gradient of pH, cation exchange capacity, and nitrogen availability (Figure 4b). The second axis can be described as bulk density and organic carbon content gradient. The ordination (Figure 3) shows that the lower precipitation (Figure 5f) and higher pH (Figure 6e), and silt content in the soil (Figure 6b) are restricted to the Kobresia mats from the Stipo regelianae-Ptilagrostion malyschevii alliance. Subalpine and alpine mesic forb meadows and pastures from the alliance Aconito rotundifolii-Potentillion pamiroalaicae are characterized by high variability with respect to climatic and soil variables (Figure 4). The exception is the association Seselideto valentinae-Oxytropidetum atbaschi which is located in the lower part of the ordination plot. These plots are characterized by higher bulk density (Figure 6f) and soil pH (Figure 6e), and low nutrient availability (Figure 6h, Figure 6i), as well as higher mean diurnal temperature range (Figure 5d) and the sum of annual precipitation (Figure 5f). On the other hand, the shrubby subalpine chionophilous heaths (Caraganetum jubatae and Caraganetum aurantiacae) occupy habitats with higher organic carbon and nitrogen content in the soil (Figure 6h, Figure 6i). The last group, Onobrychideto arenariae-Stachyetum betoniciformae, belonging to the subalpine meadow forb steppe vegetation, is characterized by high mean annual temperature (Figure 5c), annual precipitation (Figure 5f), clay content (Figure 6a) and cation exchange capacity (Figure 6g), compared to the other associations.
Figure 4
The first two axes of a three-dimensional NMDS ordination of alpine grasslands in the western Tian Shan and northern Pamir-Alai Mountains. Numbers represent three classes of analyzed vegetation: Festucetea alaico-krylovianae (1.1.1–1.1.10), Carici rupestris-Kobresietea bellardii (2.1.1–2.1.2) and Eremogono griffithii-Nepetea podostachys (3.1.1) Arrows indicate bioclimatic (a) and soil (b) variables and only significant ones are shown (p < 0.01). The ordination was run with the use of three dimensions with a stress value = 0.083. The names of syntaxa are written in the Syntaxonomic synopsis. Abbreviations: bio1 – mean annual temperature; bio2 – mean diurnal temperature range; bio4 – temperature seasonality; bio12 – annual precipitation; clay – clay content; silt – silt content; bulk_density – bulk density; catione_exchange – cation exchange capacity; C_org – organic carbon and N – nitrogen.
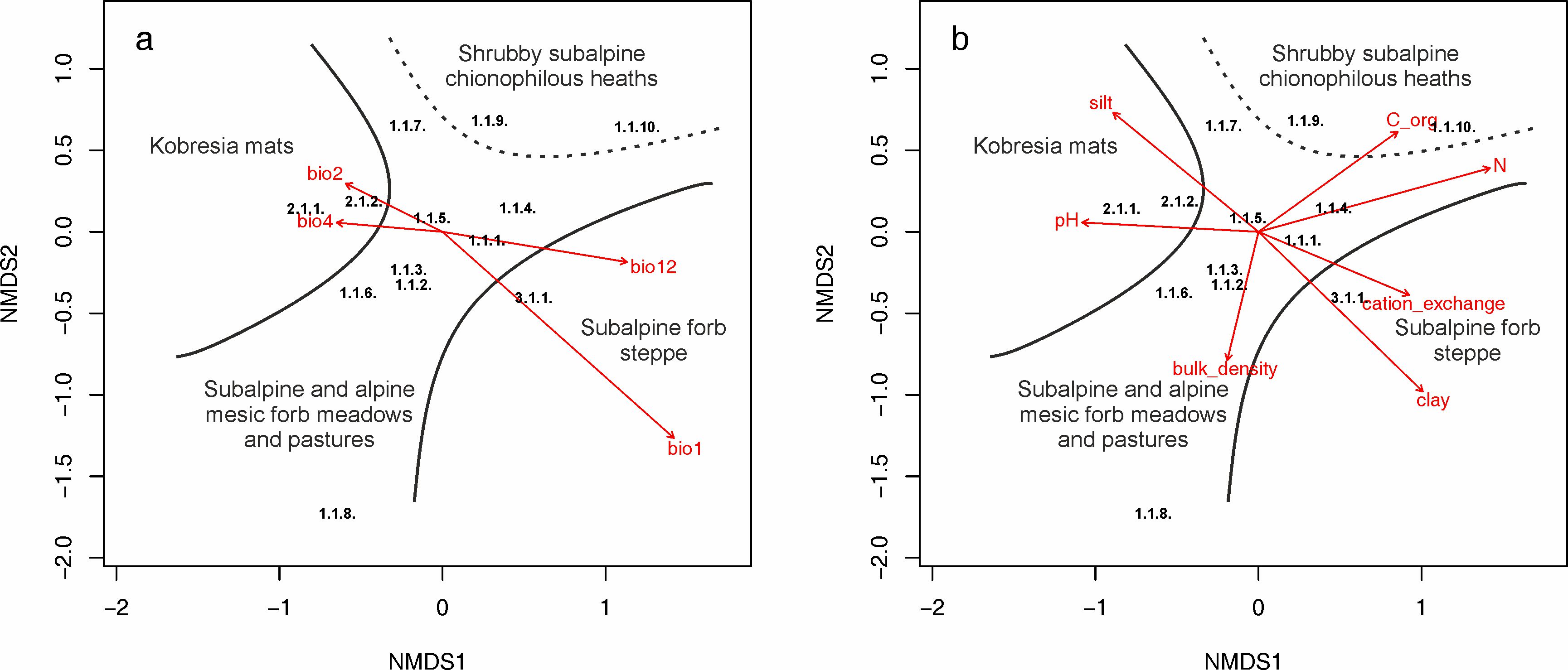
Figure 5
Boxplots showing median (line), quartiles, outliers and the range of (a) elevation, (b) slope, (c) mean annual temperature, (d) mean diurnal temperature range, (e) temperature seasonality, (f) sum of annual precipitation, (g) species richness, (h) cover of herb layer and (i) cover of cryprogram layer. The abbreviations of the syntaxonomic units are explained in the Syntaxonomic synopsis.
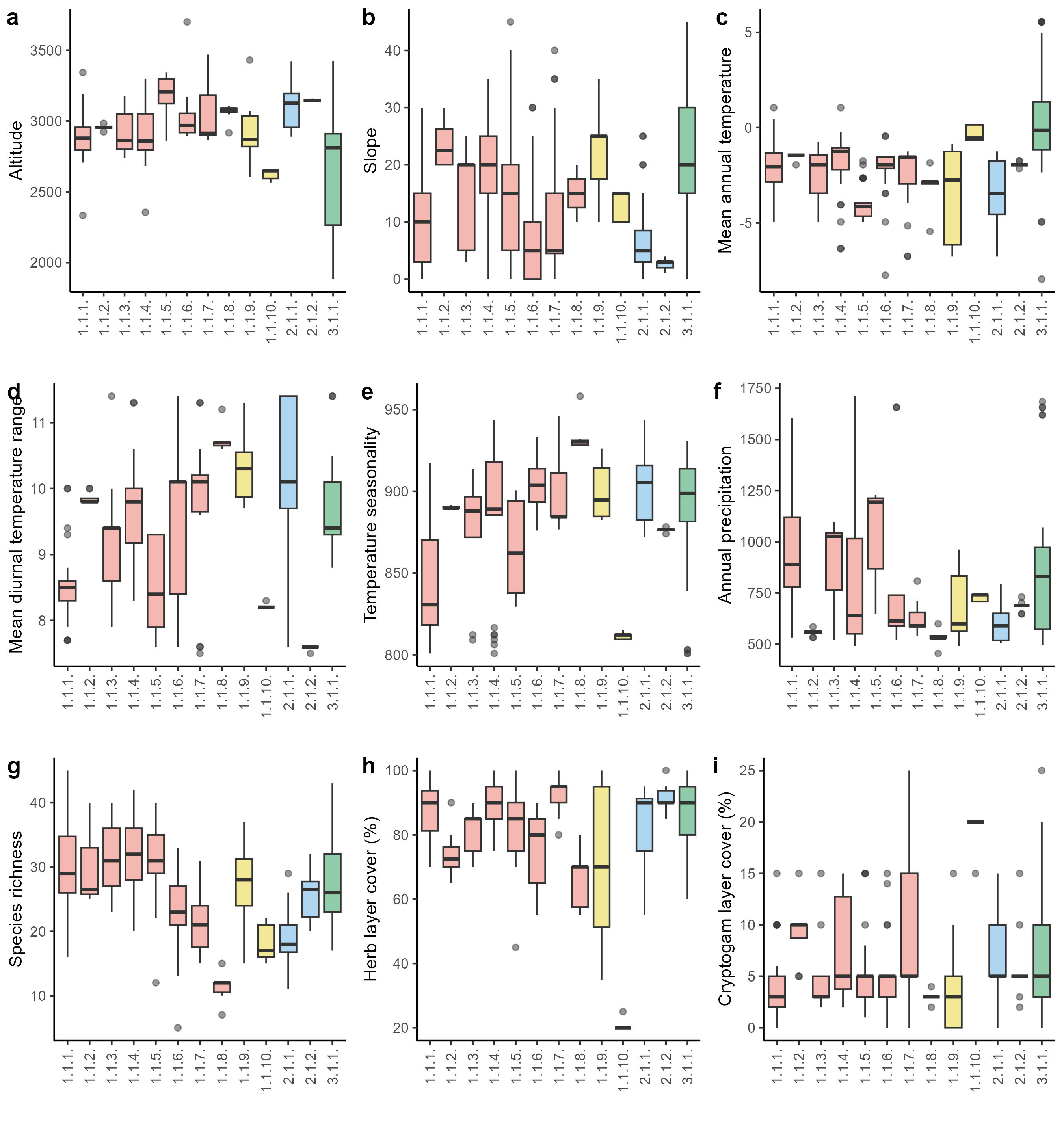
Figure 6
Boxplots showing median (line), quartiles, outliers and the range of (a) clay content (g/kg), (b) silt (g/kg), (c) sand (g/kg), (d) coarse fragments (cm3/dm3), (e) pH, (f) bulk density (cg/cm3), (g) cation exchange capacity (mmol(c)/kg), (h) organic carbon (dg/kg) and (i) nitrogen (cg/kg). The abbreviations of the syntaxonomic units are explained in Syntaxonomic synopsis.
. Syntaxonomic synopsis
Subalpine and alpine mesic forb meadows and pastures in the western Tian Shan in Kyrgyzstan
Class: Festucetea alaico-krylovianae Cl. prov.
1. Order: Geranio saxatlilis-Festucetalia alaicae ord. prov.
1.1 Alliance: Aconito rotundifolii-Potentillion pamiroalaicae A. Nowak et al. 2023 all. nova
1.1.1 Potentillo pamiroalaicae-Pedicularidetum allorramphae G. Swacha et al. 2023 ass. nova
1.1.2 Scutellario oligodontae-Hedysaretum narynense G. Swacha et al. 2023 ass. nova
1.1.3 Aulacospermo tianschanici-Pulsatilletum campanulae G. Swacha et al. 2023 ass. nova
1.1.4 Seselido tallasici-Caricetum alajicae G. Swacha et al. 2023 ass. nova
1.1.5 Poo alpinae-Eritrichietum villosae G. Swacha et al. 2023 ass. nova
1.1.6 Oxytropis globifera-Leontopodium ochroleucum community
1.1.7 Kobresio capillifoliae-Hedysaretumkirghisori G. Swacha et al. 2023 ass. nova
1.1.8 Seselideto valentinae-Oxytropidetum atbaschi G. Swacha et al. 2023 ass. nova
Shrubby subalpine chionophilous heaths of Middle Asia
Kobresia mats
Class: Carici rupestris-Kobresietea bellardii Ohba 1974
Subalpine forb steppe vegetation
Class: Eremogono griffithii-Nepetea podostachys Cl. prov.
Subalpine and alpine mesic forb meadows and pastures in the western Tian Shan in Kyrgyzstan
1.1. Alliance: Aconito rotundifolii-Potentillion pamiroalaicae A. Nowak, G. Swacha, S. Świerszcz, S. Nowak et M. Nobis 2023 all. nova hoc loco
Nomenclatural type: Aulacospermo tianschanici-Pulsatilletum campanulae G. Swacha et al. 2023
General remarks: This alliance comprises the alpine and subalpine grasslands of the Irano-Turanian region, including northern and eastern Middle Asia. The communities of this association form dense, short swards, inhabiting mountain tops, gentle slopes, and passes on mesic habitats. In Kyrgyzstan and northern Tajikistan, they serve as the most important summer pastures for sheep, goats, horses, and to some extent also cows.
Diagnostic species:Alchemilla tianschanica, Bistorta elliptica, Campanula glomerata, Cerastium pusillum, Erigeron tianschanicus, Geranium saxatile, Pedicularis korolkowii, Phlomoides oreophila, Potentilla nervosa, P. pamiroalaica, Primula algida, Rumex acetosa, Thalictrum simplex, Trisetum spicatum
Constant species: Bistorta elliptica, Geranium saxatile, Myosotis asiatica, Phlomoides oreophila
Dominant species: Geranium saxatile, Kobresia capillifolia, Phlomoides oreophila
Geographical range: Alpine and subalpine belts of northern and eastern ranges in the Irano-Turanian region (mainly Kyrgyzstan, northern Tajikistan, Kazakhstan).
Habitat characteristics: This vegetation occurs on the mesic habitats in subalpine and alpine belts of Middle Asian mountain ranges, mainly Tian Shan, northern Pamir-Alai, and probably also Altay. The communities develop on slopes with an inclination of approx. 5–40° and additionally on mountain tops and passes at the altitude of approx. 1,900–3,700 m a.s.l.
1.1.1. Potentillo pamiroalaicae-Pedicularidetum allorramphae G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 7a)
Figure 7
Photographs of the alpine grasslands in the western Tian Shan and northern Pamir-Alai Mountains belonging to: (a) Potentillo pamiroalaicae-Pedicularidetum allorramphae in the north from Kazan-Kuygna in the Naryn district; (b) Scutellario oligodontae-Hedysaretum narynense near the Dolon Pass in the Songköl Too Range; (c) Aulacospermo tianschanici-Pulsatilletum campanulaein Otmok Pass near Suusamyr Valley; (d) Seselido tallasici-Caricetum alajicae in Otmok Pass; (e) Poo alpinae-Eritrichietum villosae near the Songköl lake; (f) Oxytropis globifera-Leontopodium ochroleucum community in Fergana range west from Aral. Photographs were taken by A. Nowak.

Type relevé:Table S1, relevé number 5, holotypus hoc loco
Diagnostic species:Cerastium tianschanicum, Galium turkestanicum, Pedicularis allorrhampha, Trifolium repens
Constant species:Agrostis turkestanica, Alchemilla tianschanica, Astragalus alpinus, Cerastium tianschanicum, Dracocephalum nutans, Euphorbia alatavica, Galium turkestanicum, Minuartia biflora, Pedicularis allorrhampha, P. korolkowii, Potentilla pamiroalaica, Primula algida, Thymus incertus, Trifolium repens, Trisetum spicatum
Dominant species:Agrostis turkestanica, Alchemilla tianschanica, Bistorta elliptica, Geranium saxatile, Helictotrichon pubescens, Phlomoides oreophila, Poa angustifolia, Potentilla pamiroalaica
Habitat and distribution: Stands of this association were found at an elevational range of 2,333–3,343 m a.s.l. (mean 2,891 m a.s.l.; Figure 5a) in north-eastern (Talas Ala-Too, Ferghana Mountains, and Kyrgyz range) and central Kyrgyzstan (near alpine lake Song-Köl) (Figure 4). This association occupies usually gentle slopes and sporadically develops on slopes with steeper inclinations (Figure 5b).
Structure: Sward is very compact and short, with a mean cover of herb layer oscillating about 90% (Figure 5h) and a poorly developed moss layer, which occasionally reaches cover up to 15% (Figure 5i). The number of vascular plant species ranges from 16 to 45, with a mean of 30 species per plot (Figure 5g). High variation in species richness can be attributed to the intensity of grazing. There are no evident dominant species. Usually, several species co-dominate in various combinations, including Potentillapamiroalaica, Phlomoidesoreophila, Agrostisturkestanica, and Geraniumsaxatile.
1.1.2. Scutellario oligodontae-Hedysaretum narynense G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 7b)
Type relevé:Table S1, relevé number 3, holotypus hoc loco
Diagnostic species:Adenophora himalayana, Androsace lactiflora, Aster alpinus var. serpentimontanus, Bupleurum densiflorum, Carex stenophylla subsp. stenophylloides, Cirsium sieversii, Dracocephalum discolor, D. nodulosum, Elymus czimganicus, Elytrigia gmelinii, Erysimum altaicum, Festuca amblyodes, Hedysarum narynense, Helictotrichon hookeri, Hordeum brevisubulatum, Ligularia knorringiana, Pedicularis ludwigii, Plantago arachnoidea, Potentilla asiae-mediae, P. nervosa, Psathyrostachys kronenburgii, Pyrethrum karelinii, Scutellaria oligodonta, Thalictrum simplex, Thesium alatavicum
Constant species:Androsace lactiflora, Artemisia viridis, Aster alpinus var. serpentimontanus, Bupleurum densiflorum, Carex stenophylla subsp. stenophylloides, Cirsium sieversii, Delphinium oreophilum, Dracocephalum discolor, D. nodulosum, Elymus czimganicus, Elytrigia gmelinii, Festuca amblyodes, Galium pamiroalaicum, Geranium saxatile, Hedysarum narynense, Helictotrichon hookeri, Hordeum brevisubulatum, Leontopodium ochroleucum, Ligularia knorringiana, Lomelosia alpestris, Oxytropis globiflora, Pedicularis ludwigii, Phlomoides oreophila, Plantago arachnoidea, Potentilla asiae-mediae, P. nervosa, Pyrethrum karelinii, Scutellaria oligodonta, Thalictrum simplex, Thesium alatavicum
Dominant species:Dracocephalum nodulosum, Elymus czimganicus, Hedysarum narynense, Scutellaria oligodonta
Habitat and distribution: Stands of this association were only reported from Dolon Pass, which is located in alpine and subalpine belts between the Songköl Too and the Bayduluu Range in the Naryn Region, in the vicinity of Song-Köl Lake in central Kyrgyzstan (Figure 4). This vegetation type was found and sampled within a very narrow elevational range of 2,924–2,984 m a.s.l. (mean 2,955 m a.s.l.; Figure 5a) on steep slopes with an inclination of 20–30° (Figure 5b).
Structure: Sward is usually dense with a mean cover of herb layer of about 75% (Figure 5h) and moss layer of about 10% (Figure 5i). Hedysarum narynense is present in all patches of this vegetation type and most often dominates over the other species, and is accompanied by other codominants such as Scutellaria oligodonta, Elymus czimganicus, and Dracocephalum nodulosum. The mean number of vascular plant species per plot is high (29 species), ranging from 25 to 40 species per plot (Figure 5g).
1.1.3. Aulacospermo tianschanici-Pulsatilletum campanulae G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 7c)
Type relevé:Table S1, relevé number 43, holotypus hoc loco
Diagnostic species:Aulacospermum tianschanicum, Elymus czimganicus, Helictotrichon desertorum, Jurinea lanipes, Oxytropis melanotricha, Poa litvinoviana, Pulsatilla campanella, Thymus incertus
Constant species:Artemisia viridis, Aulacospermum tianschanicum, Elymus czimganicus, Erigeron tianschanicus, Euphrasia drosophylla, Festuca alaica, Gentiana kirilowii, G. squarrosa, Gentianella turkestanorum, Geranium saxatile, Helictotrichon desertorum, Kobresia persica, Leontopodium ochroleucum, Ligularia alpigena, Myosotis asiatica, Oxytropis globiflora, O. melanotricha, Pedicularis korolkowii, Phlomoides oreophila, Poa litvinoviana, Potentilla asiae-mediae, Pulsatilla campanella, Silene graminifolia, Thymus incertus
Dominant species:Kobresia persica, Pulsatilla campanella
Habitat and distribution:Aulacospermo tianschanici-Pulsatilletum campanulae is distributed in northern parts of Kyrgyzstan in the alpine belt of the Kyrgyz range, surrounding Dolon Pass near Song-Köl Lake and Terskey Ala-Too to the south of Issyk-Kul lake (Figure 4) at elevations ranging from 2,736–3,176 m a.s.l. (mean 2,913 m a.s.l.; Figure 5a) on slopes with various inclinations from almost flat surfaces to moderately steep slopes with inclination up to 25° (Figure 5b).
Structure: This species-rich grassland, generally dominated by Pulsatilla campanella and Kobresia persica as co-dominant, creates moderately dense swards with a mean cover of herb layer of 82% (Figure 5h) and poorly developed moss layer (averaged cover of about 5%; Figure 5i). They are generally species-rich. The number of vascular plant species ranges from 26 to 40 species per plot, with an average of 31 species (Figure 5g).
1.1.4. Seselido tallasici-Caricetum alajicae G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 7d)
Type relevé:Table S1, relevé number 77, holotypus hoc loco
Diagnostic species:Allium hymenorhizum, Astragalus alpinus, Campanula glomerata, Carex alajica, Delphinium oreophilum, Lamium album, Ligularia thomsonii, Phlomoides oreophila, Rumex acetosa, Seseli talassicum, Trollius dschungaricus
Constant species:Allium hymenorhizum, Bistorta elliptica, Campanula glomerata, Carex alajica, Galium pamiroalaicum, Geranium saxatile, Myosotis asiatica, Parnassia laxmannii, Phlomoides oreophila, Poa pratensis, Seseli talassicum, Thalictrum simplex
Dominant species:Alchemilla retropilosa, Festuca alatavica, Geranium saxatile, Hordeum brevisubulatum, Phlomoides oreophila
Habitat and distribution: The association Seselido tallasici-Caricetum alajicae is widespread across Kyrgyzstan (Figure 4) and is one of the most common plant communities in the group of mesic alpine and subalpine grasslands. It was recorded from every mountain range covered by this study. It occurs in the subalpine and alpine belts across a wide elevational range from 2,355–3,299 m a.s.l. (mean 2,911 m a.s.l.; Figure 5a) and develops mostly on north-facing moderately steep slopes with an inclination of about 20° (Figure 5b).
Structure: Herb layer is dense and, on average, covers about 90% of the surface (Figure 5h), whereas the moss layer is usually low and rarely exceeds 10% of coverage (Figure 5i). These species-rich grasslands host, on average, 32 species per plot. Species-poorest plots were composed of 20 species, whereas the richest had 42 species (Figure 5g). Specific physiognomy of this vegetation type, resembling tall-forb vegetation, is due to Phlomoides oreophila, which often takes dominance. Besides, only Geranium saxatile has relatively high cover values.
1.1.5. Poo alpinae-Eritrichietum villosae G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 7e)
Type relevé:Table S1, relevé number 107, holotypus hoc loco
Diagnostic species:Aconitum rotundifolium, Alchemilla sibirica, Bistorta elliptica, Carex orbicularis, Cerastium pusillum, Crepis oreades, Dichodon cerastoides, Draba subamplexicaulis, Dracocephalum nutans, Eremogone meyeri, Erigeron allochrous, E. aurantiacus, Eritrichium villosum, Euphorbia alatavica, Euphrasia drosophylla, Festuca alaica, Gentianella turkestanorum, Koeleria macrantha, Lomatocarpa albomarginata, Minuartia biflora, Oxytropis melanotricha, Papaver croceum, Pedicularis korolkowii, Poa alpina, Potentilla hololeuca, P. pamiroalaica, Primula algida, Rumex acetosa, Trisetum spicatum
Constant species:Aconitum rotundifolium, Bistorta elliptica, Carex orbicularis, Cerastium pusillum, Crepis oreades, Dracocephalum nutans, Erigeron allochrous, E. tianschanicus, Eritrichium villosum, Euphorbia alatavica, Euphrasia drosophylla, Festuca alaica, Gastrolychnis uralensis, Gentiana squarrosa, Gentianella turkestanorum, Geranium saxatile, Leontopodium ochroleucum, Ligularia alpigena, Minuartia biflora, Myosotis asiatica, Oxytropis melanotricha, Papaver croceum, Pedicularis korolkowii, Poa alpina, Potentilla pamiroalaica, Primula algida, Rumex acetosa, Trisetum spicatum
Dominant species:Allium atrosanguineum, Festuca amblyodes, F. alaica, Oxytropis melanotricha, Poa alpina, Potentilla hololeuca.
Habitat and distribution: This association is distributed mainly in the Kyrgyz range but was also recorded in the surroundings of Song-Köl in Central Kyrgyzstan (Figure 4). It was found at an elevation range from 2,862 to 3,345 m a.s.l. (mean 3,180 m a.s.l.; Figure 5a) on slopes with varying inclinations, from flattened surfaces to very steep ones with an inclination of 40° (Figure 5b).
Structure: The plant communities of this association create moderately dense to very compact swards with low moss coverage (about 5%; Figure 5i) and mean herb layer coverage of about 85% (Figure 5h) dominated by various combinations of most abundant Oxytropis melanotricha, Poa alpina and Festuca alaica. The number of species is, on average high, accounting for 31 species per plot, ranging from 11 to 40 species per plot (Figure 5g).
1.1.6. Oxytropis globifera-Leontopodium ochroleucum community (Figure 7f)
Diagnostic species:Androsace lactiflora, Leontopodium ochroleucum, Oxytropis globiflora, Potentilla tephroleuca
Constant species:Artemisia viridis, Aster alpinus var. serpentimontanus, Erigeron tianschanicus, Euphrasia drosophylla, Festuca amblyodes, Gentiana squarrosa, Gentianella turkestanorum, Geranium saxatile, Helictotrichon hookeri, Kobresia persica, Leontopodium ochroleucum, Oxytropis globiflora, Potentilla asiae-mediae, Potentilla tephroleuca, Pulsatilla campanella
Dominant species:Artemisia aschurbajewii, Artemisia viridis, Astragalus alpinus, Festuca amblyodes, Kobresia capillifolia, K. pamiroalaica, K. persica, Leontopodium ochroleucum, Oxytropis globiflora, Potentilla pamiroalaica
Habitat and distribution: This widespread plant community was recorded from all mountain ranges explored in this study except for the Kyrgyz range (Figure 4). It develops in the alpine belt at elevations ranging from 2,892–3,701 m a.s.l. (mean 3,028 m a.s.l.; Figure 5a) usually on flat surfaces or gentle slopes (Figure 5b).
Structure: Plant communities of Oxytropis globifera-Leontopodium ochroleucum create short and moderately dense swards on stony habitats, with the mean cover of herb layer accounting for 75% (Figure 5h) and low contribution of mosses (Figure 5i). Most stands are dominated by Leontopodium ochroleucum. However, there are many other species that can be dominants, such as Oxytropis globiflora or Kobresia pamiroalaica. Species richness varies considerably within this group. The number of vascular plant species ranges from 5 to 33 per plot, with a mean of 22 per plot (Figure 5g).
1.1.7. Kobresio capillifoliae-Hedysaretum kirghisori G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 8a)
Figure 8
Photographs of the tall-forb vegetation belonging to: (a) Kobresio capillifoliae-Hedysaretum kirghisori between the Songköl Too and the Bayduluu Range in Naryn district; (b) Seselideto valentinae-Oxytropidetum atbaschi in the At-Bashy Range; (c) Caraganetum jubatae near Sary-Tash; (d) Caraganetum aurantiacae in Suusamyr Valley near Alabel Pass; (e) Potentillo moorcroftii-Stipetum tremulae north from Kazan-Kuygna in the Naryn district; (f) Ptilagrostio malyschevii-Trisetetum altaici near Songköl lake; (g) Onobrychideto arenariae-Stachyetum betoniciformae between Aral and Jalalabad. Photographs were taken by A. Nowak.

Type relevé:Table S1, relevé number 142, holotypus hoc loco
Diagnostic species:Bistorta vivipara, Festuca alatavica, Hedysarum kirghisorum, Kobresia capillifolia, Myosotis asiatica, Parnassia laxmannii, Ptilagrostis malyschevii, Sibirotrisetum sibiricum
Constant species:Bistorta vivipara, Euphorbia alatavica, Festuca alatavica, F. amblyodes, Gastrolychnis uralensis, Kobresia capillifolia, Leontopodium ochroleucum, Ligularia alpigena, Myosotis asiatica, Oxytropis globiflora, Parnassia laxmannii, Phlomoides oreophila
Dominant species:Astragalus alpinus, Bistorta vivipara, Festuca alatavica, F. valesiaca, Hedysarum kirghisorum, Kobresia capillifolia, Stipa regeliana
Habitat and distribution: The association of Kobresio capillifoliae-Hedysaretum kirghisori was recorded mainly from the surroundings of Song-Köl alpine lake in Central Kyrgyzstan, but also from several sites in the Terskey Ala-Too and At-Bashy Range (Figure 4) within an elevational range 2,865–3,470 m a.s.l. (3,070 m a.s.l.; Figure 5a). It develops on gentle slopes, usually with approximately 10° inclination on shallow and dry soils (Figure 5b).
Structure: Plant communities of this association are always dominated by grass-like rhizomatous sedge Kobresia capillifolia, which forms very dense mats. Sporadically other species, such as Festuca alatavica and Hedysarum kirghisorum reach high coverage but always compete with the most abundant Kobresia capillifolia. The cover of the herb layer is usually more than 90% (Figure 5h), with a significant contribution of mosses creating a ground layer with coverage of about 10% (Figure 5i). Despite the strong dominance of Kobresia capillifolia, the stands of this association are moderately species-rich. The number of species per plot ranges from 15 to 31 species, with a mean of 21 (Figure 5g).
1.1.8. Seselideto valentinae-Oxytropidetum atbaschi G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 8b)
Type relevé:Table S1, relevé number 157, holotypus hoc loco
Diagnostic species:Allium oreoprasum, Androsace fedtschenkoi, Artemisia viridis, Astragalus nivalis, Eremopyrum distans, Goniolimon orthocladum, Hedysarum krylovii, Lappula stricta, Oxytropis atbaschi, O. ruebsaamenii, Poa relaxa, Seseli valentinae, Stipa caucasica, S. subsessiliflora
Constant species:Allium oreoprasum, Androsace fedtschenkoi, Artemisia viridis, Astragalus nivalis, Eremopyrum distans, Lappula stricta, Oxytropis atbaschi, O. ruebsaamenii, Potentilla pamiroalaica, Seseli valentinae, Stipa caucasica, S. subsessiliflora
Dominant species:Artemisia viridis, Oxytropis atbaschi, Stipa caucasica
Habitat and distribution: Plant communities of this association were only recorded from the At-Bashy Range in southern Kyrgyzstan (Figure 4) within a narrow elevational range of 2,917–3,105 m a.s.l. (mean 3,060 m a.s.l.; Figure 5a). It develops on stony and overgrazed habitats on gentle and moderately steep slopes with an inclination of 10–20° (Figure 5b).
Structure: Every patch of this vegetation type recorded in our study was dominated by Oxytropis atbaschi, accompanied either by nearly as much abundant Artemisia viridis or Stipa caucasica. The total cover of the herb layer is moderately low, reaching, on average, 65% (Figure 5h). The association of Seselideto valentinae-Oxytropidetum atbaschi has the lowest number of species recorded among all delimited associations, and this is probably due to high-intensity grazing and trampling. Species richness ranges from 7 to 15 species per plot, with a mean of 11 species (Figure 5g).
1.1.9. Caraganetum jubatae G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 8c)
Type relevé:Table S1, relevé number 172, holotypus hoc loco
Diagnostic species:Aconitum nemorum, Aconogonon alpinum, Astragalus lepsensis, Caragana jubata, Carex melanantha, C. regeliana, Codonopsis clematidea, Elymus longe-aristatus, Festuca pratensis, Geranium saxatile, Juniperus sabina, Lamium album, Lonicera semenovii, Papaver croceum, Pedicularis oederi, Phlomoides pratensis, Polygonatum roseum, Sanguisorba officinalis, Scrophularia griffithii, Valeriana ficariifolia
Constant species:Bistorta elliptica, Campanula glomerata, Caragana jubata, Codonopsis clematidea, Elymus longe-aristatus, Festuca alatavica, Geranium saxatile, Hordeum brevisubulatum, Lamium album, Ligularia alpigena, Myosotis asiatica, Papaver croceum, Parnassia laxmannii, Phlomoides oreophila, P. pratensis, Sanguisorba officinalis, Valeriana ficariifolia
Dominant species:Caragana jubata, Geranium saxatile
Habitat and distribution: Plant communities of the Caraganetum jubatae were recorded from subalpine and alpine belts in the surroundings of Song-Köl Lake and Terskey Ala-Too Mountains (Figure 4) at elevations of 2,608–3,431 m a.s.l. (mean 2,927 m a.s.l.; Figure 5a), primarily on steep slopes with inclinations ranging from 10 to 35° (Figure 5b).
Structure: Plant communities of this association are distinct from other alpine grasslands, except Caraganetum aurantiacae, due to the dominance of shrub-like physiognomy of dominant Caragana jubata. The herb layer under the canopy of Caragana jubata is relatively dense and covers, on average, 70% of the habitat surface (Figure 5h). The herb layer is rich in species. On average, 28 vascular plant species were recorded (Figure 5g). Sampled patches had a minimum of 15 and a maximum of 37 species per plot. In the herb layer, Geranium saxatile is the most prominent dominant species.
1.1.10. Caraganetum aurantiacae G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 8d)
Type relevé:Table S1, relevé number 176, holotypus hoc loco
Diagnostic species:Aconitum leucostomum, A. soongaricum, Allium fedtschenkoanum, Alopecurus pratensis, Bryum pseudotriquetrum, Caragana aurantiaca, Carduus nutans, Carex karoi, C. orbicularis, Cerastium falcatum, Galium tianschanicum, Hygroamblystegium varium, Ligularia heterophylla, Philonotis fontana, Rumex paulsenianus, Seseli mucronatum, Thalictrum simplex, Valeriana ficariifolia
Constant species:Aconitum leucostomum, A. soongaricum, Allium fedtschenkoanum, Alopecurus pratensis, Artemisia dracunculus, Bryum pseudotriquetrum, Campanula glomerata, Caragana aurantiaca, Carduus nutans, Carex karoi, C. orbicularis, Cerastium falcatum, Galium pamiroalaicum, G. tianschanicum, Geranium saxatile, Hygroamblystegium varium, Ligularia heterophylla, Myosotis asiatica, Philonotis fontana, Phleum pratense, Poa pratensis, Rumex paulsenianus, Seseli mucronatum, Thalictrum simplex, Valeriana ficariifolia,
Dominant species:Caragana aurantiaca
Habitat and distribution: This association was only recorded from the Kyrgyz Range (Figure 4) within a narrow elevational range of 2,564–2,655 m a.s.l. (mean 2,623 m a.s.l.; Figure 5a) in habitats situated on gentle and east-facing slopes.
Structure: The physiognomy of this association is alike to Caraganetum jubatae. Three distinct layers are distinguished. Caragana aurantiaca forms a dense shrub layer, under which a sparse herb and moss layer is developed. The total herb layer covers, on average, 21% of the habitat surface (Figure 5h). Similar coverage was observed for the moss layer (19%; Figure 5i). A minimum of 14 and a maximum of 19 species were recorded per plot, with an average of 18 species (Figure 5g). The herb layer lacks distinct dominants. The most abundant are Aconitum soongaricum, Geranium saxatile, Thalictrum simplex, and Seseli mucronatum. The highest cover in the moss layer has mire species Philonotis fontana and Hygroamblystegium varium.
2.1. Alliance: Stipo regelianae-Ptilagrostion malyschevii A. Nowak, G. Swacha, S. Świerszcz, S. Nowak et M. Nobis 2023 all. nova hoc loco
Nomenclatural type:Potentillo moorcroftii-Stipetum tremulae G. Swacha et al. 2023
General remarks: This alliance comprise alpine mat grasslands of the eastern Irano-Turanian region, including northern and eastern Middle Asia and Central Asia. The communities of this association form dense stands of graminoids, mainly Kobresia spp., Stipa spp., and Ptilagrostis malyschevii. This vegetation inhabits flattened tops and passes and gentle slopes as well. It also occurs in alpine U-shaped valleys of brooks and streams. It creates vast lands and dominates the landscape of northern Middle and Central Asia. The grasslands serve as summer pastures for horses and yaks.
Diagnostic species:Aster alpinus var. serpentimontanus, Bistorta vivipara, Gentiana kirilowii, Gentiana squarrosa, Kobresia persica, Leontopodium ochroleucum, Oxytropis globiflora, Plantago griffithii, Potentilla moorcroftii, P. tephroleuca, Ptilagrostis malyschevii, Thalictrum alpinum, Trisetum altaicum, Tulipa dasystemon
Constant species:Aster alpinus var. serpentimontanus, Festuca amblyodes, Gentiana kirilowii, G. squarrosa, Kobresia persica, Leontopodium ochroleucum, Myosotis asiatica, Oxytropis globiflora, Parnassia laxmannii, Potentilla tephroleuca
Dominant species:Bistorta vivipara, Kobresia persica, Ptilagrostis malyschevii
Geographical range: Alpine belt of northern and eastern ranges in the Irano-Turanian region (mainly Kyrgyzstan, northern Tajikistan, Kazakhstan).
Habitat characteristics: This vegetation occurs on mesic habitats in alpine belts of Middle & Central Asian mountain ranges, mainly Tian Shan, northern Pamir-Alai, and also Altay. The communities develop on slopes with an inclination of approx. 5–30° and additionally on mountain tops and passes at the altitude of approx. 2,200–4,000 m a.s.l.
2.1.1. Potentillo moorcroftii-Stipetum tremulae G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 8e)
Type relevé:Table S1, relevé number 201, holotypus hoc loco
Diagnostic species:Bistorta vivipara, Kobresia persica, Plantago griffithii, Potentilla moorcroftii, Stipa tremula, Stipa regeliana, Thalictrum alpinum
Constant species:Aster alpinus var. serpentimontanus, Bistorta vivipara, Gentiana kirilowii, G. squarrosa, Kobresia persica, Leontopodium ochroleucum, Oxytropis globiflora, Potentilla moorcroftii, Pulsatilla campanella
Dominant species:Bistorta vivipara, Carex stenocarpa, Kobresia persica, Potentilla tephroleuca
Habitat and distribution:Potentillo moorcroftii-Stipetum tremulae is the vegetation of alpine belts at elevations ranging from 2,890–3,420 m a.s.l. (mean 3,108 m a.s.l.; Figure 5a). Locations of this association were documented in the surrounding of Dolon Pass between the Songköl Too and the Bayduluu Range as well as in the area of Sarymoynak Pass to the south of Issyk-Kul Lake in the Terskey Ala-Too Mountains (Figure 4). It develops on gentle slopes and flat surfaces (Figure 5b).
Structure: It is usually dominated by grass-like rhizomatous sedge Kobresia persica and co-dominated by Bistorta vivipara. Other co-dominants, including Potentilla tephroleuca and Carex stenocarpa occur sporadically. Physiognomy in the form of low and dense mats is due to the high coverage of Kobresia persica. The total cover of the herb layer is usually about 85% (Figure 5h). A moss layer is also present, but it rarely exceeds 10% and usually ranges from 5 to 10% (Figure 5i). The mean number of species is 19 per plot, ranging from a minimum of 11 up to 29 species (Figure 5g).
2.1.2. Ptilagrostio malyschevii-Trisetetum altaici G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 8f).
Type relevé:Table S1, relevé number 213, holotypus hoc loco
Diagnostic species:Artemisia aschurbajewii, Carex stenocarpa, Euphorbia alatavica, Festuca amblyodes, Gastrolychnis uralensis, Gentiana kirilowii, Helictotrichon hookeri, Kobresia persica, Leontopodium ochroleucum, Oxytropis globiflora, Parnassia laxmannii, Poa litvinoviana, Potentilla tephroleuca, Ptilagrostis malyschevii, Ranunculus rubrocalyx, Swertia graciliflora, Trisetum altaicum, Trollius dschungaricus, Tulipa dasystemon
Constant species:Artemisia aschurbajewii, Bistorta elliptica, Carex alajica, Euphorbia alatavica, Euphrasia drosophylla, Festuca alatavica, F. amblyodes, Galium pamiroalaicum, Gastrolychnis uralensis, Gentiana kirilowii, G. squarrosa, Helictotrichon hookeri, Kobresia capillifolia, K. persica, Leontopodium ochroleucum, Ligularia alpigena, Myosotis asiatica, Oxytropis globiflora, Parnassia laxmannii, Pedicularis korolkowii, Poa litvinoviana, P. pratensis, Potentilla tephroleuca, Ptilagrostis malyschevii, Ranunculus rubrocalyx, Trisetum altaicum, Trollius dschungaricus, Tulipa dasystemon
Dominant species:Kobresia persica, Ptilagrostis malyschevii
Habitat and distribution: The association of Ptilagrostio malyschevii-Trisetetum altaici was only recorded from the near vicinity of alpine Songköl Lake in central Kyrgyzstan (Figure 4) at a narrow elevational range of 3,140–3,160 (3,147 m a.s.l.; Figure 5a). It develops on shallow soils around limestone rock outcrops on gentle slopes and flat surfaces with an average inclination of 3° (Figure 5b).
Structure: The physiognomy of this association is quite similar to the steppe vegetation due to the dominance of Ptilagrostis malyschevii, which assemblages resemble those of feather grass windswept communities at lower altitudes. Plant communities of this association are dominated by Ptilagrostis malyschevii and Kobresia persica as frequent codominant. Both species create dense swards with a mean cover herb layer of about 90% (Figure 5h). This association is species-rich. The number of species oscillates per plot between 20 and 32 species, with an average value of 25 species (Figure 5g).
3.1. Alliance: Artemision dracunculi nom. prov.
General remarks: This alliance includes typical pasture vegetation of the alpine and subalpine belts of the western part of Middle Asia. This alliance includes vegetation with fairly high productivity, especially in spring and early summer. However, the individual vegetation patches vary considerably in terms of density and species richness, which are strongly controlled by grazing. This relationship was first described in Świerszcz et al. (2023) as provisional. We decided to validate it on the basis of the larger dataset with a type association of Erigeronto seravschanicae-Eremuretum hissaricae.
Diagnostic species:Achillea millefolium, Artemisia dracunculus, Bromus inermis, Carex turkestanica, Festuca valesiaca, Galium pamiroalaicum, Geranium regelii, Onobrychis arenaria, Phleum pratense, Potentilla asiatica, Stachys betoniciflora, Stipa capillata
Constant species:Artemisia dracunculus, Carex turkestanica, Festuca valesiaca, Galium pamiroalaicum, Geranium regelii, Myosotis asiatica
Dominant species:Dracocephalum nodulosum, Festuca alatavica, F. olgae, F. valesiaca, Poa fragilis, Stachys betoniciflora
Geographical range: Alpine and subalpine belts of the western part of Middle Asia.
Habitat characteristics: The communities of this group develop in relatively moist habitats with a well-developed soil profile, most often in river and stream valley bottoms of the Tian Shan and Pamir-Alai mountain ranges. The communities develop on flat places on mountain slopes with an inclination of approx. 5–45° and additionally on mountain tops and passes at the altitude of approx. 1,900–3,500 m a.s.l.
3.1.1. Onobrychideto arenariae-Stachyetum betoniciformae G. Swacha, S. Świerszcz, S. Nowak, M. Nobis et A. Nowak 2023 ass. nova hoc loco (Figure 8g)
Type relevé:Table S1, relevé number 243, holotypus hoc loco
Diagnostic species:Achillea millefolium, Artemisia dracunculus, Bromus inermis, Carex turkestanica, Festuca valesiaca, Galium pamiroalaicum, Geranium regelii, Onobrychis arenaria, Phleum pratense, Potentilla asiatica, Stachys betoniciflora, Stipa capillata
Constant species:Carex turkestanica, Festuca valesiaca, Galium pamiroalaicum, Geranium regelii, Myosotis asiatica
Dominant species:Aconogonon coriarium, Allium hymenorhizum, Artemisia dracunculus, Carex haematostoma, C. turkestanica, Dracocephalum nodulosum, Festuca alatavica, F. olgae, F. valesiaca, Lomelosia alpestris, L. songarica, Nepeta nuda, Oxytropis pagobia, Phleum pratense, Poa fragilis, P. pratensis, Rhinanthus songaricus, Stachys betoniciflora
Habitat and distribution: The association of Onobrychideto arenariae-Stachyetum betoniciformae has a wide distribution range in Kyrgyzstan. It was recorded from each mountain range covered by this study except At-Bashy Range in southern Kyrgyzstan (Figure 4). It develops in a wide elevational gradient. Patches of this association were sampled at altitudes of 1,883–3,421 m a.s.l. (mean 2,614 m a.s.l.; Figure 5a).
Structure: This plant association has a typical meadow-like physiognomy, which is determined by a variety of plants with different life forms and architecture. The structure of vegetation is complex and multilayered and consists of tussock and rhizomatous grass and sedge species, as well as numerous forbs of different sizes. In contrast to other associations delimited in this study, Onobrychideto arenariae-Stachyetum betoniciformae is maintained mainly by mowing instead of grazing, as these meadows are used for hay and honey production. The herbaceous layer is moderately dense with a mean cover of 85% (Figure 5h). The moss layer is always present and its density ranges from 2 to 25% in coverage (Figure 5i). The number of vascular plant species ranges from 17 to 42, with a mean of 28 species per plot (Figure 5g). Usually, a single species dominates within this group, however, there is high heterogeneity in the dominance structure. Most frequent dominants are Stachys betoniciflora, Poa fragilis, Festuca valesiaca and Festuca alatavica. Our data indicate that this community is highly heterogeneous. It is likely that some of the plots may be reclassified to other communities after additional sampling.
. Discussion
. Distinction of the alpine meadows in Middle Asia
So far, the extensively studied alpine grasslands of Middle and Central Asia have not been organized into a higher-level classification scheme. Neither for Middle nor Central Asia exists a comprehensive study that presents the vegetation units that characterize individual mountain ranges (e.g. the Alai or the Tian Shan) in a hierarchical system according to the ecological-floristic approach. Our research, which was also preceded by research of meadows, steppe, semi-deserts, and tall-farb vegetation in montane and alpine belts in the Pamir, Hissaro-Alai, and western Tian Shan Mts. shows a considerable distinction of the alpine grasslands of Middle Asia. Thus, it is highly probable that a separate vegetation class should be proposed for this grassland. However, it depends on the approach (see Pignatti et al., 1995). Considering the floristic composition of all the samples collected, it seems that such a class could be well defined by the Festuca kryloviana and Festuca alaica. Both species are distributed in the alpine belt of continental Asia, with a center of occurrence in Middle and Central Asia, mainly Pamir-Alai, Tian Shan, and Altai (GBIF; www.gbif.org). This vegetation is structurally and ecologically similar to vicariant alpine and subalpine calcicolous swards of the nemoral mountain ranges of Europe Elyno-Seslerietea Br.-Bl. 1948 or, to some extent, Juncetea trifidi (Mucina et al., 2016). Also, in Middle Asia, there is a noticeable difference in the distribution of this vegetation between Tajikistan, which lies mainly outside the nemoral zone, and Kyrgyzstan, which has a typical coniferous forest belt in many mountain ranges. Between these two sub-regions runs one of the main phytogeographical boundary separating Middle Asia, with a very strong Irano-Turanian influence, and Central Asia, where, especially in the alpine belt, the contribution of the Euro-Siberian flora is pronounced (Nowak et al., 2023; Świerszcz et al., 2020). The distinctiveness of alpine grasslands in Kyrgyzstan is well indicated by the proportion of endemic species. Endemic plants are an expression of uniqueness, so-called sufficient biological content, and well-defined ecology, which is needed for the definition of a vegetation class (Pignatti et al., 1995). Among the most prominent endemics of alpine grasslands in Central Asia are the following: Schmalhausenia nidulans, Phlomoides pratensis, Oxytropis ochroleuca, Aconitum nemorum, Saussurea sordida, Alfredia acantholepis, Astragalus nivalis, Seseli valentinae, Oxytropis ruebsaamenii, Artemisia viridis, A. aschurbaevii, Tulipa dasystemon, Festuca alatavica, Hedysarum kirghisorum, and many others. This specific distinctiveness is due to geographical differences and environmental conditions, which provide refuge or are the cradle of many species (Nowak & Nobis, 2020; Nowak et al., 2011; Raduła et al., 2021). The alpine short grasslands in Western and Central Tian Shan are closely related to ranges further north and east, particularly in the Altai and Sayan Mts. One of the closely related vegetation is meadow-like tall-forbs Triseto sibiricae–Aconition septentrionalis (Ermakov et al., 2000) (Mulgedio-Aconitetea Hadač et Klika in Klika et Hadač 1944; Mirkin & Naumova, 2012). This grassland type shares several species with alpine meadows of Kyrgyzstan, including Anthoxanthum alpinum, Minuartia biflora, Galium boreale, Helictotrichon pubescens, H. hookeri and Trisetum sibiricum (=Sibirotrisetum sibiricum; Ermakov & Zibzeev, 2012; Zibzeev & Nedovesova, 2015). In addition, this vegetation shares some similarities with subalpine tall-forb vegetation Aconito-Geranietea albiflori reported from the Svjatoj Nos range in Lake Baikal district (syn. of Mulgedio-Aconitetea; Chytrý et al., 1993; Ermakov et al., 2000). They are not only species-rich and unique but also have a characteristic use, which is relatively intensive grazing. The most important animal species that are grazed on the subalpine grasslands in both regions are sheep and goats. Geographically, the alpine vegetation of Middle Asia, especially in its northern part (western and central Tian Shan), is closely linked to areas of mountain glaciation and subsequent reestablishment of vegetation migrating from the south and populating the alpine belt above the upper forest limit. These compositional similarities can be clearly seen at the genus level, including such common taxa as Trisetum, Oxytropis, Aconitum, Seseli, Geranium, Bistorta, Phlomoides, Primula, etc.
The final decision on whether Middle Asian alpine short grasslands should be distinguished at the rank of the class or only order requires further thorough comparative studies. In our opinion, it should be ranked as the class that comprises the Altai-Sayan order of alpine swards Festucetalia krylovii and Pamir-Alai-western Tian Shan order Geranio saxatlilis-Festucetalia alaicae (Ermakov & Zibzeev, 2012; Zibzeev & Nedovesova, 2015). There can be two approaches to the alpine vegetation classification development at the scale of continents. One of them is a description of a series of “narrow” ecologically vicarious local classes based on only formal floristic differences. Another approach is an application of a wide geographical concept of classes. The latter one was proposed by Pignatti et al. (1995). Then, the high regional floristic peculiarities observed in the alpine communities may be demonstrated at the level of orders. We decided, for this unified vegetation in all mountainous alpine zones in Asia in terms of structure and phylogeny, to use the latter approach.
. Comparisons of the alpine meadows of western Tian Shan in Middle Asia to related vegetation in surrounding regions and zones
Alpine meadows in western Tian Shan share a number of species with alpine meadows and subalpine forb steppes in Pamir-Alai (Świerszcz et al., 2023). These include, among others, Festuca pratensis, Elymus longe-aristatus, Saussurea sordida, Ligularia alpigena, Geranium saxatile and Pulsatilla campanella. However, the frequency and cover of these species is not particularly high. Still, the high floristic differences on the two sides of the main phytogeographical boundary with Irano-Turanian influences in the Pamir-Alai and temperate climates in the Tian Shan make the alpine meadows in Kyrgyzstan very distinct. Despite the endemic taxa that were mentioned above, a lot of Euro-Siberian species contribute to this vegetation, among others, Sanguisorba officinalis, Luzula multiflora subsp. sibirica, Pedicularis oederi, Aconogonon alpinum, Myosotis asiatica, Botrychium lunaria, Dianthus superbus, Sibirotrisetum sibiricum, Poa alpina, Achillea millefolium, Minuartia biflora, and others. These species, with the addition of others like e.g. Veronica porphyriana, Eritrichium villosum, Leontopodium ochroleucum, or Poa litvinoviana, make these grasslands comparable to Altai alpine vegetation (Ermakov & Zibzeev, 2012; Zibzeev & Nedovesova, 2015). Provisionally established order for this grasslands - Violo altaicae-Festucetalia krylovianae (Ermakov & Zibzeev, 2012) seems to be a geographical vicariant for the western Tian Shans’ alpine meadows that are included in Geranio saxatlilis-Festucetalia alaicae.
From the closer subhumid areas in SE Kazakhstan in the Ala-tau Mts. some data on alpine meadows were reported by Dimeyeva et al. (2016). They mostly represent typical alpine grasslands, which mainly consist of Geranium saxatile, Phlomoides oreophila, Dracocephalum imberbe, Alopecurus pubescens, Helictotrichon pubescens, and Pedicularis kryloviana. The structure and species composition of these communities appears to be very similar to the alpine grasslands of western Tian Shan. They have been found at slightly lower elevations and are less species-rich, which is understandable in the context of the latitudinal gradient. It is ca. 10–25 species in Ala-tau in comparison to up to 45 (mean 26) in western Tian Shan (Dimeyeva et al., 2016). From the highest parts of the Zhalanash area in SE Kazakhstan, many species that are common to Tian Shan have been found, among others Bistorta vivipara, B. elliptica, Primula algida, Festuca kryloviana, Poa alpina, Geranium saxatile, Trisetum altaicum, T. spicatum, Leontopodium fedtschenkoanum, Viola altaica, etc. The decision whether include this vegetation in the Ala-tau Mts. in Geranio saxatlilis-Festucetalia alaicae or Festucetalia krylovianae requires further comparative studies.
There is still no obvious and certain solution for the cold-adapted Caragana scrub classification. The patches of Caragana jubata and C. aurantiaca found in the Tian Shan, and Pamir-Alai have the character of an alpine meadow with a considerable share of fen or mire species in the herbaceous layer. In Mongolia, it was included in Kobresietalia myosuroidis and Caraganion jubatae alliance (Mirkin et al., 1983). However, for the territory of Sangilen Mts., it was placed in Saussureo schanginianae–Caraganetum jubatae and Loiseleurio–Vaccinietea Eggler 1952 (Zibzeev et al., 2018). This vegetation embraces the alpine and arctoalpine shrubby heath communities of Eurasia and North America. To some extent, patches of Caragana jubata and C. aurantiaca resembles the vegetation of Vaccinio microphylli-Juniperetalia nanae Rivas-Mart. et M. Costa 1998, a sub-xeric and sub-thermophilous low juniper scrub in the supramontane to subalpine belts of Southern Europe and the Caucasus (Mucina et al., 2016). They form in the landscape very clearly distinguishable patches in places where grazing is less intensive. It constitutes the primary heaths of the upper montane to alpine belt above the coniferous forests’ upper line (Picea schrenkiana and Abies semenovii). This vegetation type requires further study in Central Asia, a region that is a hotspot of the diversity of Caragana genus (Zhang et al., 2016).
The floristic linkages with more distant alpine meadow habitats are insignificant. Mention can be made of Primula algida, which occurs frequently in the alpine meadows of the Caucasus (Etzold et al., 2016), or Stipa purpurea, which is an important element of the alpine grasslands in the Himalayas and Tibet (Nobis et al., 2020, 2022). But it is important to point out that some of our samples belong to a separate, much more widespread class in the mountains of Asia - Carici rupestris-Kobresietea bellardii. This graminoid tundra-like short grasslands on wind-exposed sites in the alpine and subnival belts of Eurasia occurs also in Tian Shan on summits, passes and gentle slopes with long-lasting snow cover. In Asia, mat-like grasslands were reported from Caucasus (Kobresietalia capilliformis Tsepkova 1987; Onipchenko, 2002), Altai (Ermakov & Zibzeev, 2012), and Alatau Mts (Dimeyeva et al., 2016). These communities are replaced by cryophilous steppes (Nowak et al., 2018) or alpine semi-deserts (Nowak et al., 2023) towards dryer habitats and alpine meadows towards more fertile habitats. In both cases, the snow cover in such places is much shorter than in Kobresia mats. In western Tian Shan, short graminoid grasslands have close relationships and floristic similarities with alpine swards of Karakorum and the Himalayan range. We propose an alliance that will group this vegetation together (Stipo regelianae-Ptilagrostion malyschevii) with two associations: Potentillo moorcroftii-Stipetum purpureae and Ptilagrostio malyschevii-Trisetetum altaici. These communities form dense swards with high overall plant cover, a typical feature of this vegetation type with significant contribution of Pamir and Himalayan mountain ranges like Potentilla moorcroftii (=Sibbaldianthe moorcroftii) or Stipa regeliana.
The collected data also includes several relevés from the lower subalpine (upper montane) sites from the Naryn Valley near the Kara-Koo Pass (2,925 m a.s.l.) and Talas Valley close to Alabel Pass (3,184 m a.s.l.). These were classified as forb steppe grasslands, which have their center of distribution in the Pamir-Alai in Tajikistan and probably are also widely distributed in the Hindu Kush. The only association, Onobrychideto arenariae-Stachyetum betoniciformae occupies gentle slopes at approx. 1,800–2,200 m a.s.l. and has an important contribution of steppe or pseudo-steppe species like Festuca valesiaca, Delphinium biternatum, Astragalus sieversianus or Stipa kirghisorum. There also occur species that were considered diagnostic for Nepetalia podostachys (Świerszcz et al., 2023), like Geranium regelii or Ligularia thomsonii. This transitional vegetation between the steppe and meadow was already reported from more distant regions of continental Asia, among others from the southern Urals and Inner Mongolia in China (Ren et al., 2008; Wang & Ba, 2008). This vegetation is characterized by species from Salvia, Phlomis, Seseli, Stachys, Veronica, and Erysimum genera. Species diversity of this forb steppes can be high, having up to 50 species per 10 m2 with an average cover of about 60–65%. This type of vegetation is in close contact with typical alpine meadows, cryophilous steppes, or pseudo-steppes. It can also have a close relationship with dry tall forbs that develop within juniper woodland belts - in their lower part, mainly thermophilous communities of Prangetea ulopterae, and in the upper part communities with a considerable share of Eremurus spp. and Phlomoides spp. taxa. (Nowak et al., 2020, 2022). These montane grasslands are secondary vegetation after forests that were removed in favor of pastures (Wagner, 2009). Since the last century, these meadows have been used mostly for hay-making (Karmysheva, 1982; Korovin, 1962).
. Conclusions
Our study has expanded the knowledge of the alpine grasslands on the borderland between vegetation influenced by the Irano-Turanian climate in Pamir-Alai and grassland vegetation of the alpine belt in Tian Shan, which is strongly affected by the Temperate climate. Despite the structural and functional similarities associated with a homogeneous response to climatic factors, including, among others, the length of snow cover, these communities show relatively high variability. Their final classification certainly still requires further studies and comparisons, but it seems that the alpine grasslands in the Altai region are the closest to the alpine swards of western Tian Shan. This is why we have proposed to include them in one class of alpine vegetation that is characterized by a large proportion of both Irano-Turanian and Euro-Siberian flora and thrives under a temperate climate with alpine influences. Some of the vegetation units are closely related to the Karakorum-Himalayan ranges and thus need to be linked to the semi-desert grasslands and alpine pastures of this area (Dvorskỳ et al., 2011; Hartmann, 1968, 1972).
. Supplementary material
The following supplementary material is available for this article:
Figure S1. Dendrogram illustrating the assignment of relevé groups identified by Ward’s method to particular syntaxonomic units.
Table S1. The analytic table of alpine grasslands in the western Tian Shan Mts.
Table S2. Synoptic table of alpine grasslands in the western Tian Shan Mts.
Tables S1 and S2 are available in the repository Figshare: https://doi.org/10.6084/m9.figshare.22586293
