. Introduction
Middle Asia is traditionally referred to Kyrgyzstan, Tajikistan, Uzbekistan, Turkmenistan, southern Kazakhstan, western China, and northern Afghanistan. Southern and eastern parts of Middle Asia encompass the high mountains of the Pamir-Alai and Tian Shan Mts., with a vast alpine belt occurring mainly above 2,600–3,000 m a.s.l. According to the ten-volume study of the flora of the former Soviet part of Middle Asia, more than 9,000 vascular plant species are known from this region (Khassanov, 2015). The species pool for grasslands only for Tajikistan includes approximately 2,000 species (about 1,350 in steppes, 370 in meadows and pastures, and 265 in alpine swards). Considering generally low endemism at high altitudes, Middle Asia, with 136 alpine and subnival narrow endemic taxa, clearly stands out in this regard. Middle Asia is recognized as one of the world’s 35 hotspots of biodiversity (Mittermeier et al., 2011). In view of the fact that mountain ecosystems are particularly sensitive to climate change (Baettig et al., 2007), it is essential to foster studies on their diversity.
Middle Asia is a large region of grasslands where approximately 2 million km2 is covered by this plant formation (Wesche et al., 2016). Vegetation of the alpine and subnival grasslands belongs to the major group of natural Arctic-alpine types of vegetation that develop in climates that are too cold to sustain forests (Dengler et al., 2014, 2020). Alpine grasslands are fragile ecosystems formed by plants that are adapted to survive and reproduce in harsh environments. Alpine and subnival changes along with environmental factors such as topography, duration and thickness of snow cover, nutrient and water availability, and wind exposure (Isselin-Nondedeu & Spiegelberger, 2014).
Recently, several studies have been conducted on alpine and subnival vegetation of Irano-Turanian and Middle Asia regions, revealing five main groups of grassland vegetation: (1) high-altitude thorn-cushion grasslands, (2) dry, cryophilous steppes, and semi-deserts, (3) alpine Kobresia matts, (4) snow-bed vegetation, and (5) alpine meadows and pastures (Akhani et al., 2013; Dimeyeva et al., 2016; Noroozi et al., 2010, 2017; Swacha et al., 2023; Świerszcz et al., 2023; Vanselow et al., 2016). The alpine and subnival grasslands extend along all Middle Asian mountain ranges and comprise distinct zonal vegetation between 2,500 and 4,000 m a.s.l. They are used as summer pastures and are grazed by yaks, horses, and sheep. Middle Asian alpine meadows and swards, as well as subnival vegetation, are distinct in terms of phytogeography and play a crucial role in the livelihood of the local people; however, they have been poorly studied. Significant research has recently been completed in the high Pamir (Vanselow, 2011; Vanselow et al., 2016). Several vegetation units of high alpine grasslands have been described, including alpine matts, meadows, swards, and pastures (Jusufbekov, 1968; Ladygina & Litvinova, 1971; Vanselow, 2011; Walter & Breckle, 1986).
Only a few phytosociological surveys have been conducted on alpine and subnival vegetation from adjacent areas. Several grassland communities have been recorded in the alpine belt of the Altai Mts. of south-eastern Kazakhstan (Rubtsov, 1948). They were included in high-altitude vegetation and called ‘low-herb alpine grasslands’ following the Russian classification approach. Traditionally named Kobresia-forb swards (with Kobresia capilliformis, K. humilis, Alchemilla retropilosa and Leontopodium fedtschenkoanum) and rupiculous alpine vegetation on fine debris (Dracocephalum imberbe, Saussurea glacialis, Rhodiola coccinea, Potentilla biflora, Oxygraphis glacialis, and Trollius altaicus) have been reported from the Altai Mts. (Dimeyeva et al., 2016; Rubtsov, 1948). Towards the main Himalayan chain, in eastern Ladakh, the Kobresia royleana–Kobresia schoenoides matts, and Poa attenuata–Potentilla pamirica grasslands were reported from elevations up to 5,700 m a.s.l. (Dvorskỳ et al., 2011).
In Alborz Mts., the alpine and subnival vegetation was defined as the Oxytropis persica-Catabrosella parviflora community group (Akhani et al., 2013). It inhabits a diverse range of ecological niches, including rupiculous, xerophytic, and hygrophilous habitats at approximately 3,500–4,000 m a.s.l. This chionophilous vegetation has already been mentioned by Klein (1982) in this region, who proposed the class Oxytropidetea persciae and the order Trachydietalia depressae for the snow-bed vegetation of the Middle East, which thrives at elevations of 3,100 to 4,350 m a.s.l. However, Noroozi et al. (2010) included this vegetation in the order Drabetalia pulchellae, which comprises three alliances: Astragalion iodotropidis, Cousinion multilobae, and Acantholimion demawendici. Generally, the floristic composition at genus rank is quite similar to the Pamir Alai alpine vegetation, with Astragalus, Alchemilla, Veronica, Oxytropis, Potentilla, Silene, Allium, and Cousinia as the richest genera (Noroozi et al., 2020). In this context, Klein’s proposal to consider this type of nival vegetation in Middle Asia described by Ukacheva (1975) Oxytropeta immersae as vicariant to the Iranian subnival grasslands and swards seems very relevant. Additionally, Noroozi et al. (2010, 2017) distinguished three alliances (Astragalion polyanthi, Astragalion sahendi and Taraxaco brevirostris-Polygonion serpyllacei) for thorn-cushion grasslands and snow-bed vegetation, with 15 associations and one additional subassociation for the alpine and subnival belts of Sahand Mts., Sabalan Mts. and Bozqush Mts. in NW Iran. While the thorn-cushion vegetation is very distinct in terms of the floristic composition, the snow-bed vegetation shares a considerable proportion of taxa common to Middle Asia (e.g., Festuca alaica, Oxytropis savellanica, Kobresia schoenoides, Oxyria digyna, and Gnaphalium supinum). The same applies to the rupiculous habitats of the subnival belt, where among the most common taxa in the Alborz Mountains are those also known from the highest elevations of Middle Asia (e.g. Didymophysa, Draba, Dracocephalum, Erysimum, Lamium, Oxytropis, Paraquilegia; Noroozi et al., 2020). This vegetation was classified in the Didymophyso aucheri-Dracocephaletea aucheri class and the Didymophysetalia aucheri order for the Iranian territory (Noroozi et al., 2014).
In the Hindu Kush, typical alpine grasslands are restricted to the Central and Eastern Hindu Kush (Breckle & Rafiqpoor, 2020). This vegetation type is intensively used as pastures during the short summer. In drier places, often with annual precipitation lower than 100 mm, the vegetation becomes more sparse, creating open alpine matts and swards (Breckle & Rafiqpoor, 2020). In the subnival belt, the patchy snow-bed or rupiculous vegetation consists of a few taxa, including Cystopteris dickieana, Primula macrophylla, Solmslaubachia flabellata, S. surculosum, and Sibbaldia cuneata, which occur up to 5,600 m a.s.l. (Breckle & Rafiqpoor, 2020). On the other hand, it is also important to highlight that the nival vegetation of Anatolia shows strong phytogeographical and ecological affinities with the mountain ranges located eastwards. At the subnival belt, mat- and cushion-forming vegetation dominates (Drabo-Androsacetalia, Astragalo-Brometea; Parolly, 2004; Quézel, 1973), in which small pulvinate, creeping, and suffruticose chamaephytes dominate together with caespitose hemicryptophytes (Parolly, 2020). In more damp places along meltwater runners or snow beds, vegetation rich in geophytes and hemicryptophytes is well established, creating dense carpet turfs (Trifolio anatolici-Polygonetalia arenastri, Astragalo-Brometea; Parolly, 2020).
To better understand the diversity and phytogeographical similarities of alpine and subnival grasslands, it is necessary to study them in detail in the central part of Asia, at the node of the great continental mountain chains meeting in Middle Asia and the Pamir.
In this paper, we attempt to classify the high alpine and subnival vegetation in the Pamir-Alai and western Tian Shan Mts. and relate it to the subalpine grasslands of this area. We aimed to address the following questions during our study: (1) What is the diversity of grassland plant communities of the alpine and subnival belts in the Pamir-Alai and western Tian Shan Mts? (2) What are the basic environmental conditions of the described plant communities? (3) What is the species composition and structure of the vegetation types? (4) Which species have important diagnostic value for the described syntaxa?
. Material and methods
. Study area
The vegetation survey was carried out in several ecoregions of Central Asia within the administrative boundaries of Kyrgyzstan and Tajikistan. The study area lies between latitudes of 37.485° and 42.483°, and longitudes of 68.237° and 79.124°, comprising a significant part of both countries. Most plots (139) were sampled in a hotspot of high-altitude vegetation in the Pamir alpine and desert tundra ecoregion (Dinerstein et al., 2017). Vegetation-plot data were also collected from other ecoregions: the Tian Shan montane steppe and meadows (97 plots) and from the transitional areas between the Tian Shan foothill and steppe and the Tian Shan montane conifer forests (67 plots). All these areas are part of two large systems of mountain ranges in Central Asia: the western Tian Shan and the Pamir-Alai Mountain system. Despite the wide spatial extent of the study area, we searched for accessible mountain ranges with the occurrence of habitats suitable for the development of alpine and subnival vegetation, which are found above treeline at altitudes between 2,600 and 4,800 m a.s.l. Study sites at lower altitudes are used by local societies as summer pastures for sheep and goats and less often for horses and yaks. Areas near the snow line with sparse vegetation, are grazed sporadically, or grazing never occurs. Here, as elsewhere in the mountains, climatic conditions depend on altitude, but locally, the vegetation can be more affected by exposition and relief rather than by elevation. Local relief in the alpine and nival belts is crucial for the radiation, temperature, and water regimes. In addition, it determines the depth and duration of snow cover. The coupled effect of land relief and elevation on vegetation is often more important than macroclimate, which is difficult to generalize for spatially distanced mountain ranges. Nevertheless, the common feature of all surveyed mountain ranges is that they are influenced by a relatively dry macroclimate with high continentality and the highest precipitation in summer. We expect high vegetation variability along environmental gradients considering high variation in macro- and micro-scale features of the surveyed mountain ranges, including mean annual air temperature (min: −13.1 °C, max: 7.6 °C, mean: −5.7 °C), annual precipitation (min: 181.5 mm yr−1, max: 1,507 mm yr−1, mean: 724.5 mm yr−1), and also in the number of days with snow cover (min: 110, max: 365, mean: 308) based on snowpack model by Paulsen and Körner (2014).
. Data sampling and statistical analysis
The research was conducted in the years 2013–2022. Altogether, 303 relevés were collected in the Pamir-Alai (Tajikistan) and western Tian Shan Mts. (Kyrgyzstan). The size of each vegetation plot ranged from 1 to 10 m2. In each plot, all vascular plant species and mosses were recorded using the seven-degree cover-abundance scale of the Braun-Blanquet approach. The geographical coordinates of each plot were recorded with the help of a GPSMAP 60CSx device with an accuracy of ±5 m, using the WGS84 standard.
Data were stored in the Vegetation of Middle Asia database (A. Nowak et al., 2017) and analyzed in R (R Core Team, 2023) and JUICE software (Tichý, 2002). To understand the distribution of samples and the relations between them, we performed an unsupervised k-means analysis with the Hellinger transformation. The number of clusters was determined according to gap statistics using the clusGap function in the ‘cluster’ package (Maechler et al., 2019) in the R environment. The algorithm indicated 18 groups as the most optimal for the analyzed dataset, which corresponds well with our field experience. Diagnostic species for each group were identified using the phi coefficient as a fidelity measure (Chytrý & Tichý, 2003). The size of all groups was standardized to equal size, and Fisher’s exact test (p < 0.05) was applied. The phi coefficient values were multiplied by 100. Species with a phi coefficient ≥ 40 and frequency > 20% were considered diagnostic for the associations, a threshold phi ≥ 30 was used for alliances and phi ≥ 25 for orders. Species with a frequency higher than 50% were defined as constant, and those with a maximum cover value exceeding 30% were defined as the dominant species of an individual community. For alliances and orders, the threshold for constant species was frequency > 40% and for dominant > 50%. Non-metric multidimensional scaling (NMDS) based on Euclidean distance was performed to assess floristic relationships among the classified vegetation types using the function metaMDS in the ‘vegan’ package (Oksanen et al., 2019). Prior to the analysis, all cover values of the Braun-Blanquet scale present in the analyzed dataset were transformed to a percentage scale (+, 1, 2, 3, 4 to 2%, 3%, 13%, 38%, 63% respectively) and log(x + 1) transformed. The final ordination was run with 999 random starts using three dimensions (stress value = 0.199). Next, we fitted bioclimatic variables post hoc to the ordination axes to explore their associations with each vegetation type using the function envfit with 999 permutations in the ‘vegan’ package (Oksanen et al., 2019). Climatic data were extracted from the CHELSA database version 2.1 (http://chelsa-climate.org; Karger et al., 2017). Prior to analysis, correlations between 19 bioclimatic variables were calculated using the Spearman correlation coefficient to reduce the number of available variables. Variables indicating strong multicollinearity (r > 0.75 in all pairwise comparisons) were removed, and we retained variables most clearly interpretable from an ecological point of view. The climatic variables used for analysis were mean annual temperature, mean diurnal temperature range, mean temperature of the wettest quarter, mean temperature of the driest quarter, annual precipitation, precipitation seasonality, and precipitation of coldest quarter.
A shortened synoptic table with the fidelity and relative percentage frequency of all diagnostic species and other species with frequency ≥ 20% is shown in Table 1. An analytic table including type relevés (Table S1) and a full synoptic table (Table S2) are available in the Figshare Digital Repository (https://doi.org/10.6084/m9.figshare.22758647.v2; S. Nowak et al., 2024).
Table 1
Shortened synoptic table with percentage frequency and fidelity values. Only diagnostic species are given and other species with frequency ≥ 20%. See supplementary material 2 for the full version of this table (Table S2). Group No.: 1 – Violo acutifoliae-Potentilletum biflorae, 2 – community of Leiospora bellidifolia and Ajania trilobata, 3 – Carici stenocarpae-Androsacetum akbaitalensis, 4 – Dracocephaletum imberbe, 5 – Dracocephaletum nodulosi-Rosularietum borissovae, 6 – Achoriphragmetum pinnatifidi, 7 – community of Oxyria digyna, 8 – Nepetetum longibracteatae, 9 – Nepeto pamirensis-Rhodioletum recticaulis, 10 – Acantholimonetum diapensioidis, 11 – Violo thianschanicae-Stipetum regelianae, 12 – Papavero crocei-Ranunculetum popovii, 13 – Littledaleo alaicae-Stipetum trichoidis, 14 – Bistorto ellipticae-Pedicularietum korolkowii, 15 – Poo attenuatae-Callianthemetum alatavici, 16 – Poo bucharicae-Oxytropidetum savellanicae, 17 – Drabo alajicae-Omalothecetum supinae, 18 – community of Trollius komarovii and Erigeron heterochaeta. Abbreviations of vegetation layers in the second column: ml – moss layer, hl – herb layer and sl – shrub layer.
The syntaxa nomenclature follows Mucina et al. (2016), Noroozi et al. (2014), A. Nowak et al. (2018) and Świerszcz et al. (2023). The nomenclature of the vascular plants follows Plants of the World Online (POWO, 2023), and problematic taxonomic issues were based on The World Flora Online (WFO, 2023). The nomenclature of Stipa spp. follows Nobis et al. (2020) and that of Geranium spp. follows Cherepanov (1995). The plant material collected during field studies was deposited in the Herbarium of Middle Asia Mountains, hosted in OPUN (University of Opole, Poland) and KRA (Jagiellonian University, Poland).
. Results
. Classification of the vegetation units
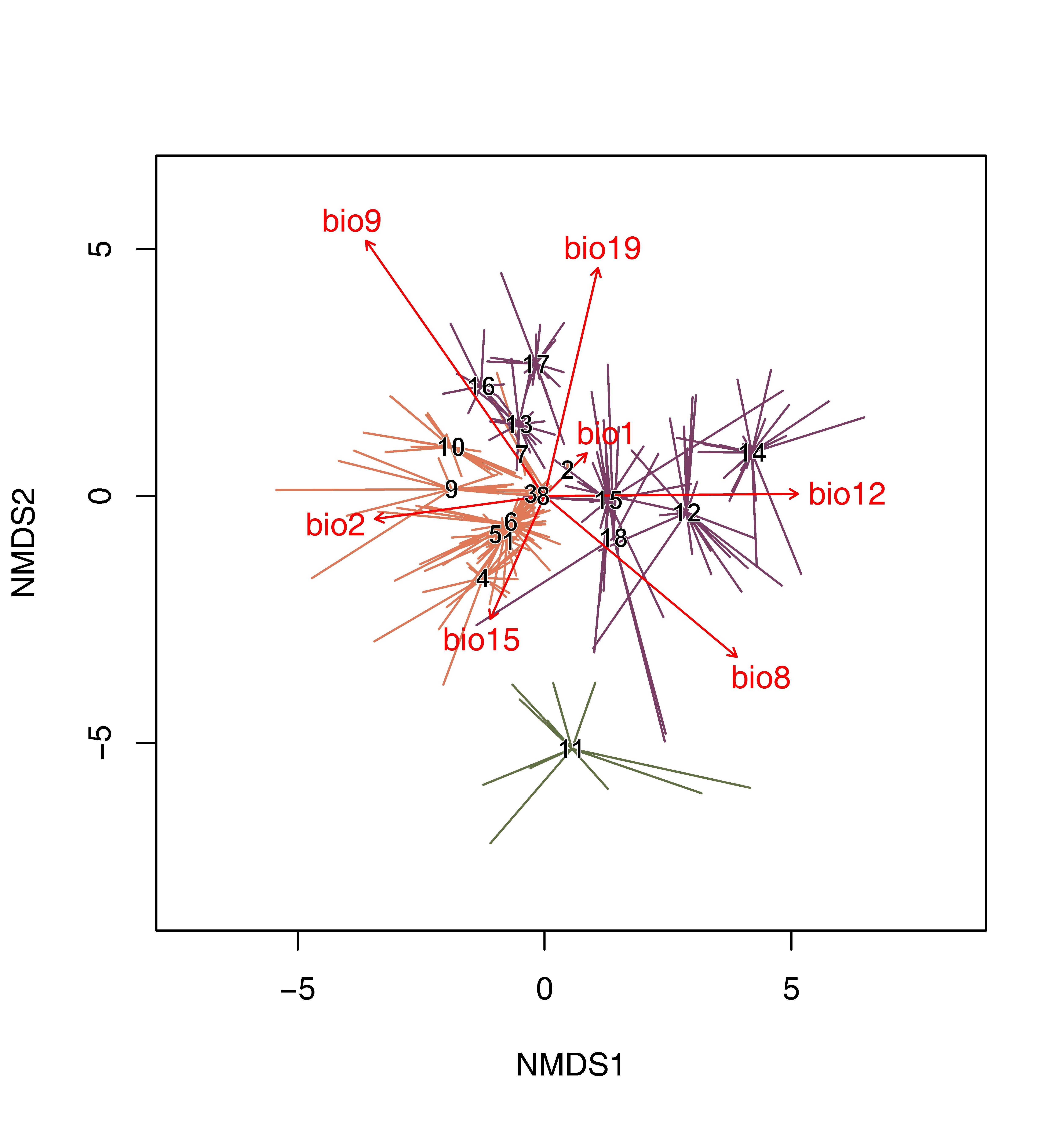
We identified 18 communities from three classes of high alpine and subnival vegetation: Didymophyso aucheri-Dracocephaletea aucheri (groups 1–10), Carici rupestris-Kobresietea bellardii (group 11), and Festucetea alaico-krylovianae (groups 12–18) in Tajikistan and Kyrgyzstan (Figure 1). The classification results were fitted to NMDS ordination (Figure 2), and the shortened synoptic table is presented in Table 1. The first axis of the NMDS reflects a substantial mean diurnal air temperature range–precipitation gradient. The plots that were classified into high altitude subnival rupiculous and snow-bed vegetation (groups 1–10) of the order Drabo pamiricae-Androsacetalia akbaitalensis are associated with a higher temperature range of temperatures and a lower sum of annual precipitation compared to the plots classified as mesic alpine pastures (groups 12, 14, 15 and 18) of the order Geranio saxatilis-Festucetalia alaicae. The second axis is linked with mean temperature of the driest quarter and precipitation of the coldest quarter, where higher values of these bioclimatic variables differentiate Littledaleo alaicae-Stipetum trichoidis, Poo bucharicae-Oxytropidetum savellanicae and Drabo alajicae-Omalothecetum supinae associations from other communities within class Festucetea alaico-krylovianae. The outlier in the lower part of the diagram is the association Violo thianschanicae-Stipetum regelianae – the chionophilous summit graminoid and dwarf-scrub mountain tundra in the alpine and subnival belts that occupies the habitats with higher precipitation seasonality.
Figure 1
Distribution of relevés assigned to the particular vegetation units (n = 303): 1 – Violo acutifoliae-Potentilletum biflorae, 2 – community of Leiospora bellidifolia and Ajania trilobata, 3 – Carici stenocarpae-Androsacetum akbaitalensis, 4 – Dracocephaletum imberbe, 5 – Dracocephaletum nodulosi-Rosularietum borissovae, 6 – Achoriphragmetum pinnatifidi, 7 – community of Oxyria digyna, 8 – Nepetetum longibracteatae, 9 – Nepeto pamirensis-Rhodioletum recticaulis, 10 – Acantholimonetum diapensioidis, 11 – Violo thianschanicae-Stipetum regelianae, 12 – Papavero crocei-Ranunculetum popovii, 13 – Littledaleo alaicae-Stipetum trichoidis, 14 – Bistorto ellipticae-Pedicularietum korolkowii, 15 – Poo attenuatae-Callianthemetum alatavici, 16 – Poo bucharicae-Oxytropidetum savellanicae, 17 – Drabo alajicae-Omalothecetum supinae, 18 – community of Trollius komarovii and Erigeron heterochaeta.

Figure 2
The first two axes of a three-dimensional NMDS ordination of chionophilous vegetation in the high alpine and subnival belts of the Pamir-Alai and western Tian Shan Mts. Numbers correspond to plant communities included within three classes of analyzed vegetation: Didymophyso aucheri-Dracocephaletea aucheri (1–10), Carici rupestris-Kobresietea bellardii (11) and Festucetea alaico-krylovianae (12–18). Arrows indicate bioclimatic variables: bio1 – mean annual temperature, bio2 – mean diurnal temperature range, bio8 – mean temperature of the wettest quarter, bio9 – mean temperature of the driest quarter, bio12 – annual precipitation, bio15 – precipitation seasonality and bio19 – precipitation of the coldest quarter. The ordination was run with the use of three dimensions with stress value = 0.199. The names of syntaxa are given in Figure 1 and syntaxonomic synopsis.
. Description of syntaxa
Syntaxonomic synopsis
High altitude subnival rupiculous and snow-bed vegetation
Class: Didymophyso aucheri-Dracocephaletea aucheri Noroozi et al. 2014
Order: Drabo pamiricae-Androsacetalia akbaitalensis A. Nowak et al. 2024 in S. Nowak et al. 2024 ord. nov. hoc loco
Alliance: Oxytropido savellanicae-Nepetion pamiricae A. Nowak et al. 2024 in S. Nowak et al. 2024 all. nov. hoc loco
1. Violo acutifoliae-Potentilletum biflorae S. Nowak et al. 2024 ass. nov. hoc loco
2. Community of Leiospora bellidifolia and Ajania trilobata
3. Carici stenocarpae-Androsacetum akbaitalensis S. Nowak et al. 2024 ass. nov. hoc loco
4. Dracocephaletum imberbe S. Nowak et al. 2024 ass. nov. hoc loco
5. Dracocephaletum nodulosi-Rosularietum borissovae S. Nowak et al. 2024 ass. nov. hoc loco
6. Achoriphragmetum pinnatifidi A. Nowak et al. 2014
7. Community of Oxyria digyna
8. Nepetetum longibracteatae S. Nowak et al. 2024 ass. nov. hoc loco
9. Nepeto pamirensis-Rhodioletum recticaulis S. Nowak et al. 2024 ass. nov. hoc loco
Group of tussock and cushion subnival and alpine vegetation
10. Acantholimonetum diapensioidis S. Nowak et al. 2024 ass. nov. hoc loco
- variant with Lagotis decumbens
Chionophilous summit graminoid and dwarf-scrub mountain tundra in the alpine and subnival belts
Class: Carici rupestris-Kobresietea bellardii Ohba 1974
Order: Kobresietalia capilliformis Tsepkova 1987
Alliance: Stipo regelianae-Ptilagrostion malyschevii A. Nowak et al. 2023
11. Violo thianschanicae-Stipetum regelianae S. Nowak et al. 2024 ass. nov. hoc loco
Mesic grazed alpine pastures on fertile soils
Class: Festucetea alaico-krylovianae nom. prov.
Order: Geranio saxatilis-Festucetalia alaicae S. Nowak et al. 2024 ord. nov. hoc loco
Alliance: Poo alpinae-Bistortion ellipticae S. Świerszcz et al. 2023
12. Papavero crocei-Ranunculetum popovii S. Nowak et al. 2024 ass. nov. hoc loco
13. Littledaleo alaicae-Stipetum trichoidis A. Nowak et al. 2018
14. Bistorto ellipticae-Pedicularietum korolkowii S. Nowak et al. 2024 ass. nov. hoc loco
15. Poo attenuatae-Callianthemetum alatavici S. Nowak et al. 2024 ass. nov. hoc loco
16. Poo bucharicae-Oxytropidetum savellanicae S. Nowak et al. 2024 ass. nov. hoc loco
17. Drabo alajicae-Omalothecetum supinae S. Nowak et al. 2024 ass. nov. hoc loco
18. Community of Trollius komarovii and Erigeron heterochaeta
High altitude subnival rupiculous and snow-bed vegetation
Order: Drabo pamiricae-Androsacetalia akbaitalensis S. Nowak et al. 2024 ord. nov. hoc loco
Nomenclatural type: Oxytropido savellanicae-Nepetion pamiricae A. Nowak, S. Nowak, S. Świerszcz, M. Nobis, G. Swacha 2024 in S. Nowak et al. 2024 all. nov. hoc loco
General remarks: This order comprises the subnival rupiculous and snow-bed vegetation of the eastern Irano-Turanian highlands. It is distributed mainly in the Central Asia mountains. The communities of this order form sparse and short grasslands or cushions, composed mainly of Nepeta spp., Oxytropis spp., Paraquilegia spp., Sibbaldia spp., Festuca spp., Poa spp., and Richteria spp. This vegetation inhabits rupicolous, often eroded, and gravelly habitats in the extremely harsh environments of Middle and Central Asia alpine semi-deserts and cryophilous grasslands.
Diagnostic species: Bryum intermedium, Distichium capillaceum, Draba pamirica, Nepeta pamirensis, Paraquilegia anemonoides, Potentilla multifida, Richteria pyrethroides, Schistidium apocarpum, Sibbaldia tetrandra
Constant species: Poa litvinoviana
Dominant species: Dasiphora dryadanthoides, Dasiphora phyllocalyx, Dracocephalum imberbe, Nepeta pamirensis, Oxytropis savellanica, Parrya pinnatifida, Potentilla biflora, Sibbaldia tetrandra
Geographical range: Alpine and subnival belts of the northern and eastern ranges of Pamir-Alai and Tian Shan (mainly Kyrgyzstan, northern Tajikistan, Kazakhstan, Xinjang in China).
Habitat characteristics: This vegetation occurs in mesic high alpine and subnival habitats in mountain ranges, mainly Tian Shan, northern and eastern Pamir-Alai and the Altai Mts. These communities develop at an altitude of approximately 2,600–4,700 m a.s.l. on slopes with an inclination of 2–95°, within the zone of mean annual temperature below zero and precipitation from 180 to 1,200 mm.
Alliance: Oxytropido savellanicae-Nepetion pamiricae A. Nowak, S. Nowak, S. Świerszcz, M. Nobis, G. Swacha 2024 in S. Nowak et al. 2024 all. nov. hoc loco
Nomenclatural type: Nepeto pamirensis-Rhodioletum recticaulis S. Nowak et al. 2024 ass. nov. hoc loco
This alliance includes high-elevation grasslands in relatively dry areas with often patchy vegetation. It is a transitional vegetation between dense alpine swards and mats (in places with higher rainfall) and cushion-tragacanthic vegetation, which most often occupies drained slopes and eroded passes. Patches of these communities are very often adjacent to screes, rocky communities, or badlands, which means that they often occupy rupicolous habitats with poorly developed soil profiles. Phytocoenoses of this alliance occur over a wide altitudinal amplitude, occupying sites from approximately 2,600 to 4,700 m a.s.l. (Figure 3A). The vegetation of this alliance is characterized by fairly low species richness of vascular plants (Figure 3D) compared with the communities of the other two alliances. The patches of the Oxytropido savellanicae-Nepetion pamiricae phytocoenoses are relatively sparse and have a quite low cover of the herb layer (Figure 3F).
Diagnostic species: Bryum intermedium, Distichium capillaceum, Nepeta pamirensis, Paraquilegia anemonoides, Richteria pyrethroides Schistidium apocarpum,
1. Violo acutifoliae-Potentilletum biflorae S. Nowak et al. 2024 ass. nov. hoc loco (Figure 4A)
Figure 3
Boxplots showing median (line), quartiles, outliers and the range of (A) elevation, (B) slope, (C) total species richness, (D) vascular plant species richness (E) cryptogram species richness, (F) herb layer cover and (G) cryptogram layer cover. The names of syntaxa are given in Figure 1 and syntaxonomic synopsis.
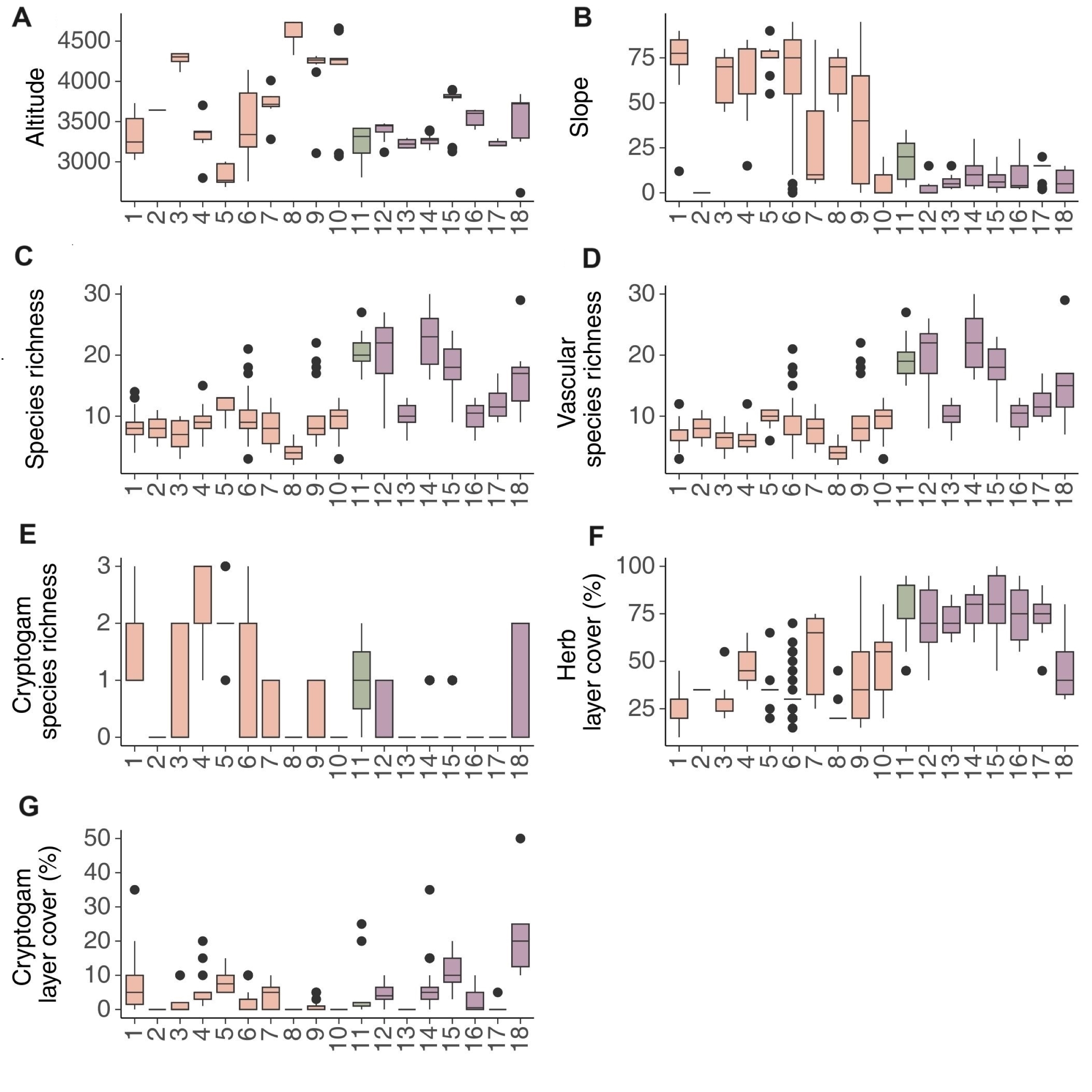
Figure 4
Photographs of the high altitude subnival rupiculous and snow-bed vegetation: (A) Violo acutifoliae-Potentilletum biflorae; (B) Leiospora bellidifolia and Ajania trilobata community; (C) Carici stenocarpae-Androsacetum akbaitalensis; (D) Dracocephaletum imberbe; (E) Dracocephaletum nodulosi-Rosularietum borissovae and (F) Achoriphragmetum pinnatifidi. All photographs were taken by A. Nowak.

Holotypus: Table S1, relevé number 19, holotypus hoc loco.
Diagnostic species: Aulacospermum tenuisectum, Bryum intermedium, Paraquilegia anemonoides, Potentilla biflora, Tortella tortuosa, Viola acutifolia
Constant species: -
Dominant species: Bryum intermedium, Cystopteris fragilis, Dasiphora parvifolia, Dasiphora phyllocalyx, Eritrichium pseudolatifolium, Paraquilegia anemonoides, Potentilla biflora, Saxifraga oppositifolia, Sibbaldia tetrandra
Floristic and habitat characteristics: Violo acutifoliae-Potentilletum biflorae is a subnival rupicolous vegetation found in north-eastern Kyrgyzstan in the Ak-Suu and Jeti-Ögüz districts of the Issyk-Kul region on the slopes of the Terskey Ala-Too and Kakshaal Too ranges, at elevations between 3,024 and 3,730 m a.s.l. (mean = 3,323 m a.s.l; Figure 3A). The association develops on slopes with varying exposures but mostly with a north-western aspect. The inclination of the slopes ranges from 12° up to 90° (mean = 75°; Figure 3B). The vascular species richness of this phytocoenosis is low and ranges from three to 12 taxa per plot (on average six; Figure 3D), and the cover of the herb layer is from 10 to 45% (average of about 25%; Figure 3F). Mosses also appeared on plots ranging from one to three with coverage up to 35% (Figure 3E, Figure 3G).
2. Community of Leiospora bellidifolia and Ajania trilobata (Figure 4B)
Diagnostic species: Ajania trilobata, Alopecurus apiatus, Leiospora bellidifolia, Oxytropis coelestis, Ranunculus alberti
Constant species: Festuca alaica
Dominant species: Ajania trilobata
Floristic and habitat characteristics: The community has a character of subnival rupicolous vegetation and has been recorded in western Tian-Shan at an altitude of approximately 3,650 m a.s.l. (Figure 3A) in the Jeti-Ögüz district of the Issyk-Kul region in north-eastern Kyrgyzstan. The number of vascular plant species ranges from five to 11 with a mean of eight (Figure 3D), and the cover of the herb layer of approximately 35% (Figure 3F). The vegetation is single-layered, and no mosses were found. The final classification and assignment of this community requires more data from Leiospora bellidifolia and Ajania trilobata distribution ranges.
3. Carici stenocarpae-Androsacetum akbaitalensis S. Nowak et al. 2024 ass. nov. hoc loco (Figure 4C)
Holotypus: Table S1, relevé number 33, holotypus hoc loco.
Diagnostic species: Androsace akbaitalensis, Carex stenocarpa, Eritrichium pamiricum
Constant species: Poa litvinoviana, Richteria pyrethroides
Dominant species: Androsace akbaitalensis, Androsace sericea, Scrophularia pamirica
Floristic and habitat characteristics: Carici stenocarpae-Androsacetum akbaitalensis is subnival rupicolous vegetation noted in the Pamirs in eastern Tajikistan (Gorno-Badakhshan). This association develops on steep slopes with an inclination of 45° to 80° (mean = 64°; Figure 3B), most often with a north-western and northern exposure, at alpine elevations ranging from 4,115 to 4,343 m a.s.l. (mean = 4,287 m a.s.l.; Figure 3A). The patches of this vegetation are species poor with three to 10 species per plot (mean = 7; Figure 3C). The cover of the herb layer is relatively low and ranges from 20 to 55% (average about 29%; Figure 3F).
4. Dracocephaletum imberbe S. Nowak et al. 2024 ass. nov. hoc loco (Figure 4D)
Holotypus: Table S1, relevé number 41, holotypus hoc loco.
Diagnostic species: Bryum intermedium, Allium atrosanguineum, Distichium capillaceum, Draba parviflora, Dracocephalum imberbe, Kobresia persica, Schistidium apocarpum
Constant species: Poa litvinoviana
Dominant species: Allium atrosanguineum, Dracocephalum imberbe, Hylotelephium ewersii, Ptilagrostis malyschevii, Sibbaldia tetrandra
Floristic and habitat characteristics: Dracocephaletum imberbe is a subnival rupicolous vegetation growing in northern Kyrgyzstan, in the Chüy and Issyk-Kul regions, and in north-central Kyrgyzstan in the Kochkor district. The association develops in the western Tian-Shan at high altitudes ranging from 2,799 to 3,702 m a.s.l. (Figure 3A), most often on the south-western, southern and south-eastern slopes with an inclination of about 15°–85° (mean = 61.5°; Figure 3B). The richness of vascular plant species is low, with 4 to 12 taxa per plot (mean = 6; Figure 3C). Mosses are not very important in the phytocenoses, with one to three species with low coverage (Figure 3E, Figure 3G). The mean value of the herb cover is about 45%, ranging from 35% to 65% (Figure 3F).
5. Dracocephaletum nodulosi-Rosularietum borissovae Nowak et al. 2024 ass. nov. hoc loco (Figure 4E)
Holotypus: Table S1, relevé number 67, holotypus hoc loco.
Diagnostic species: Allium talassicum, Androsace aflatunensis, Bupleurum aitchisonii, Dracocephalum nodulosum, Ferula transiliensis, Grimmia tergestina, Hylotelephium ewersii, Oxytropis trichosphaera, Potentilla asiae-mediae, Pyrethrum kovalevskiae, Rosularia borissovae, R. vvedenskyi, Schistidium apocarpum, Stipa caucasica, Syntrichia ruralis
Constant species: Poa litvinoviana
Dominant species: Potentilla asiae-mediae, Rosularia borissovae
Floristic and habitat characteristics: The Dracocephaletum nodulosi-Rosularietum borissovae association is a typical alpine rupicolous vegetation. Plots of this phytocoenosis have been noted in the Kochkor district of the Naryn region (north-central Kyrgyzstan). They occupy steep slopes with an inclination of 55° to 90° (mean = 75°; Figure 3B), mostly with a south-western exposition, at an altitude of 2,687 to 3,003 m a.s.l. (average 2,832 m; Figure 3A). The number of vascular plant species in a plot ranges from six to 11, with a mean of 10 (Figure 3D). The mean value of the herb cover is 36% ranging from 20% to 65% (Figure 3F), and the cover of the moss layer varies between five to 15% (average 8%; Figure 3G).
6. Achoriphragmetum pinnatifidi A. Nowak et al. 2014 (Figure 4F)
Diagnostic species: Parrya pinnatifida
Constant species: -
Dominant species: Artemisia viridis, Carex haematostoma, Dasiphora dryadanthoides, Dasiphora phyllocalyx, Delphinium oreophilum, Hylotelephium ewersii, Leymus lanatus, Minuartia pusilla, Paraquilegia anemonoides, Parrya pinnatifida, Parrya schugnana, Silene graminifolia
Floristic and habitat characteristics: This association appears over a wide range of altitudes from 2,757 to 4,143 m a.s.l. (mean = 3,420 m a.s.l.; Figure 3A) with varying slope aspects, most often western, north-western, northern, and north-eastern. The inclination of slopes ranged from 2° to 95° (mean = 67°; Figure 3B). The number of vascular plant species ranges from three to 21, with a mean of 10 (Figure 3D). The cover of the herb layer ranges from 15 to 70% (mean = 33%; Figure 3F). Noteworthy is the presence of mosses with a maximum cover of 10% (on mean = 3%; Figure 3G).
7. Community of Oxyria digyna
Diagnostic species: Allium tianschanicum, Oxyria digyna, Saxifraga hirculus
Constant species: -
Dominant species: Primula turkestanica, Saxifraga sibirica, Sibbaldia olgae
Floristic and habitat characteristics: Oxyria digyna community is an alpine and subnival rupicolus vegetation recorded in the western Tian Shan Mts. in the Jeti-Ögüz district of the Issyk-Kul region (north-eastern Kyrgyzstan), in the Kochkor district of the Naryn region (north-central Kyrgyzstan) and in the Pamirs in Gorno-Badakhshan region (eastern Tajikistan) at elevations of 3,280 to 4,011 m a.s.l. (mean = 3,714 m a.s.l.; Figure 3A). The community was noted mainly on the north-eastern and north-western slopes at varied inclination from 5° to 85° (Figure 3B). The cover of the herb layer ranges from 25 to 75% (Figure 3G). Plots were consisted of four to 13 species, with an average of approx. eight (Figure 3C).
8. Nepetetum longibracteatae S. Nowak et al. 2024 ass. nov. hoc loco (Figure 5A)
Figure 5
Photographs of the high altitude subnival rupiculous and snow-bed vegetation: (A) Nepetetum longibracteatae; (B) Nepeto pamirensis-Rhodioletum recticaulis; (C) Acantholimonetum diapensioidis and chionophilous summit graminoid and dwarf-scrub mountain tundra in the alpine and subnival belts: (D) Violo thianschanicae-Stipetum regelianae. All photographs were taken by A. Nowak.

Holotypus: Table S1, relevé number 128, holotypus hoc loco.
Diagnostic species: Didymophysa fedtschenkoana, Elymus czimganicus, Erigeron pamiricus, Nepeta longibracteata, Parrya stenocarpa
Constant species: Richteria pyrethroides
Dominant species: Dasiphora dryadanthoides
Floristic and habitat characteristics: Nepetetum longibracteatae is a typical subnival rupicolous vegetation. It has been recorded in the Gorno-Badakhshan region, near the Verkhniy Muzhkul Nature Reserve, at the highest altitudes of 4,327–4,740 m a.s.l. (Figure 3A). Plots of this association were found mainly on the south-western and southern slopes with varying inclinations from 45° to 80° (mean = 64°; Figure 3B). Despite subnival elevation, this area is characterized by low annual precipitation. Species richness is low, with only 2–7 species per plot (mean = 4; Figure 3C), with the cover of the herb layer varying from 20 to 45% (mean = 25%; Figure 3F)
9. Nepeto pamirensis-Rhodioletum recticaulis S. Nowak et al. 2024 ass. nov. hoc loco (Figure 5B)
Holotypus: Table S1, relevé number 136, holotypus hoc loco.
Diagnostic species: Grimmia funalis, Hymenolaena nana, Nepeta pamirensis, Rhodiola recticaulis
Constant species: Poa litvinoviana
Dominant species: Artemisia skorniakowii, Carex melanantha, Eritrichium pamiricum, Nepeta pamirensis, Parrya asperrima, Parrya turkestanica, Poa litvinoviana, Potentilla pamiroalaica, Ranunculus badachschanicus, Semenovia furcata, Semenovia pamirica
Floristic and habitat characteristics: Patches of Nepeto pamirensis-Rhodioletum recticaulis have been recorded in the Pamir Mts. in the Shughnon district (central-western part of Gorno-Badakhshan) in eastern Tajikistan and in the Kyrgyz Ala-Too range in the North Tian-Shan (Chüy region). The association forms alpine and subnival patchy swards at elevations of 3,107–4,315 m a.s.l. (Figure 3A). It inhabits slopes with an average inclination of approx. 45°, ranging from 3° to 95° (Figure 3B), most often with a north-western and western aspect. The number of vascular plant species varies between five and 22, with a mean of 10 (Figure 3D). The total cover of the herb layer in the plots ranges between 15% and 95% (average approximately 42%; Figure 3F).
Group of tussock and cushion subnival and alpine vegetation
10. Acantholimonetum diapensioidis S. Nowak et al. 2024 ass. nov. hoc loco (Figure 5C)
Holotypus: Table S1, relevé number 150, holotypus hoc loco.
Diagnostic species: Acantholimon diapensioides, Draba pamirica, Lagotis decumbens, Oxytropis savellanica, Potentilla multifida
Constant species: Poa litvinoviana
Dominant species: Acantholimon diapensioides, Dracocephalum paulsenii, Hedysarum poncinsii, Nepeta pamirensis, Oxytropis savellanica, Potentilla multifida, Potentilla pamirica, Saussurea glacialis
Floristic and habitat characteristics: Acantholimonetum diapensioidis can be defined as alpine cushion vegetation that develops on the gentle slopes of the Kyrgyz Ala-Too range in the North Tian-Shan (N Kyrgyzstan) and on the gentle slopes and flattened barelands of the Eastern Pamir (e.g. Shugnan Mts., Muzkol Mts.). The association occurs on slopes of various exposures with an inclination of 3° to 20° (Figure 3B) at altitudes from 3,070 to 4,660 m a.s.l. (Figure 3A). The species richness is low, with 3–13 species per plot (Figure 3C). The cover of the herb layer ranges from 20 to 80% (Figure 3F). The vegetation is single-layered with no mosses.
Variant with Lagotis decumbens
Diagnostic species: Lagotis decumbens
Within this association, a habitat variant with Lagotis decumbens stands out. Patches with this species develop in wetter depressions or in the vicinity of watercourses near Dzhiland or Akbaital Pass at high altitudes from 4,224 to 4,285 m a.s.l. They resemble subnival wet rupicolous vegetation.
Chionophilous summit graminoid and dwarf-scrub mountain tundra in the alpine and subnival belts
Alliance: Stipo regelianae-Ptilagrostion malyschevii A. Nowak et al. 2023
The alliance includes compact and short alpine and subnival grasslands in relatively moist habitats and long-lasting snow cover. They are used as pastures for horses and yaks. The vegetation belonging to this alliance is dominated by graminoids, mainly Kobresia spp., Stipa spp., Poa spp, Carex spp. The patches of Stipo regelianae-Ptilagrostion malyschevii occupy elevations of approximately 2,500 to 3,500 m a.s.l. (Figure 3A) and have moderate species richness compared to other alpine vegetation in the study area (mean = 14; Figure 3C).
Diagnostic species: Alchemilla krylovii, Astragalus lepsensis, Bistorta elliptica, Bryum pseudotriquetrum, Bupleurum densiflorum, Callianthemum alatavicum, Carex haematostoma, C. melanantha, C. regeliana, Cerastium tianschanicum, Dasiphora parvifolia, Draba oreades, Erigeron aurantiacus, E. umbrosus, Gentiana tianschanica, Geranium saxatile, Helictotrichon mongolicum, Juniperus sabina, Kobresia capilliformis, K. pamiroalaica, Leontopodium ochroleucum, Lonicera semenovii, Oxytropis gorbunovii, Parnassia laxmannii, Philonotis fontana, Phlomoides pratensis, Potentilla crantzii, Ptilagrostis malyschevii, Rhodiola heterodonta, Salix caesia, Sanguisorba officinalis, Scorzonera subacaulis, Stipa regeliana, Thalictrum alpinum, Veronica luetkeana, Viola thianschanica
11. Violo thianschanicae-Stipetum regelianae S. Nowak et al. 2024 ass. nov. hoc loco (Figure 5D)
Holotypus: Table S1, relevé number 175, holotypus hoc loco.
Diagnostic species: Astragalus lepsensis, Bryum pseudotriquetrum, Carex haematostoma, C. melanantha, Dasiphora parvifolia, Draba oreades, Gentiana tianschanica, Helictotrichon mongolicum, Kobresia capilliformis, K. pamiroalaica, Leontopodium ochroleucum, Oxytropis gorbunovii, Parnassia laxmannii, Phlomoides pratensis, Ptilagrostis malyschevii, Sanguisorba officinalis, Stipa regeliana, Thalictrum alpinum, Viola thianschanica
Constant species: Bistorta elliptica
Dominant species: Caragana jubata, Carex melanantha, Kobresia capilliformis, Kobresia persica, Salix caesia, Stipa regeliana
Floristic and habitat characteristics: Violo thianschanicae-Stipetum regelianae forms an alpine and subnival mats or short grasslands with a dominance of graminoids. This vegetation serves as pastures on gentle slopes of the Terskey Ala-Too range (the Jeti-Ögüz and Ak-Suu districts of the Issyk-Kul region) and near Kara-Unkur (the Kochkor district of the Naryn region). The plots were sampled at an elevation of 2,807–3,422 m a.s.l. (mean = 3,250 m; Figure 3A) with varying slope exposures in north-eastern Kyrgyzstan. The inclination of the slopes was low and ranged between 3° and 35° (mean = 18°; Figure 3B). The patches were relatively rich and harbor 16 to 27 species in a particular relevé (Figure 3C). The cover of the herb layer ranges from 45 to 95%, with a high mean of more than 80% (Figure 3F). The cover of the moss layer was rather low, occasionally reaching up to 25% (Figure 3G).
Mesic grazed alpine pastures on fertile soils
Order: Geranio saxatilis-Festucetalia alaicae S. Nowak et al. 2024 ord. nov. hoc loco
Nomenclatural type: Poo alpinae-Bistortion ellipticae S. Świerszcz et al. 2023 hoc loco
General remarks: This order comprises alpine short grasslands of the eastern Irano-Turanian region, including Middle and Central Asia. The communities of this order form dense stands of graminoids, mainly Carex spp., Festuca spp., Trisetum spp., and Poa spp., as well as short forbs, e.g. Erigeron spp., Gentiana spp., Potentilla spp., Ranunculus spp., Cerastium spp., and Primula spp. The grasslands serve as summer pastures for sheep and horses, rarely yaks.
Diagnostic species: Allium oreophilum, Androsace dasyphylla, Aspicilia fruticulosa, Carex alajica, C. orbicularis, Cerastium pusillum, Dichodon cerastoides, Draba alajica, D. altaica, Dracocephalum nutans, Erigeron allochrous, E. heterochaeta, E. tianschanicus, Eritrichium villosum, Festuca alaica, F. brachyphylla, Gentiana algida, G. kirilowii, G. squarrosa, Gentianella turkestanorum, Geranium collinum, Helictotrichon tianschanicum, Koeleria pyramidata, K. spicata, Lagotis ikonnikovii, Lomatocarpa albomarginata, Minuartia biflora, Omalotheca supina, Oxytropis globiflora, O. immersa, Papaver croceum, Poa attenuata, Potentilla flabellata, P. hololeuca, P. pamiroalaica, Primula algida, Pyrethrum karelinii, Ranunculus alberti, Schmalhausenia nidulans, Schulzia crinita, Stipa trichoides, Taraxacum kovalevskiae, Trollius komarovii
Constant species: Bistorta elliptica, Carex orbicularis, Dichodon cerastoides, Erigeron allochrous, Festuca alaica, Koeleria spicata, Leontopodium ochroleucum, Poa alpina, Poa litvinoviana, Schulzia crinita
Dominant species: Carex alajica, Eritrichium villosum, Festuca alaica, Oxytropis savellanica, Poa alpina, Poa attenuata, Potentilla crantzii, Stipa trichoides
Geographical range: Alpine belt of the northern and eastern ranges in the Irano-Turanian region (mainly Kyrgyzstan, northern Tajikistan, Kazakhstan, Xinjang in China).
Habitat characteristics: This vegetation occurs in mesic habitats in the alpine belts of Middle and Central Asian mountain ranges. These communities develop on gentle slopes or flattened areas with an inclination of approx. 0–30°, well moistened with mean annual precipitation between 500 and 1,500 mm. The main altitudinal distribution of these alpine grasslands in Middle Asia is between approx. 3,200 and 3,600 m a.s.l.
Alliance: Poo alpinae-Bistortion ellipticae S. Świerszcz et al. 2023
This vegetation covers typical alpine grasslands with a large proportion of graminoids and a considerable share of Euro-Siberian species. They are distributed mainly in the Tian Shan Mts., preferably on the northern slopes. They are used as pastures, especially for sheep, and less often for yaks, horses, and goats. Patches belonging to this alliance are generally characterized by high species richness and high cover of herbs and mosses (Figure 3C, Figure 3F–G) compared with the other two alliances.
Diagnostic species: Androsace dasyphylla, Carex alajica, C. orbicularis, Dichodon cerastoides, Draba altaica, Erigeron allochrous, Eritrichium villosum, Festuca alaica, Gentiana squarrosa, Koeleria spicata, Minuartia biflora, Omalotheca supina, Oxytropis immersa, Papaver croceum, Potentilla flabellata, P. hololeuca, Primula algida, Ranunculus alberti, Schulzia crinita
12. Papavero crocei-Ranunculetum popovii S. Nowak et al. 2024 ass. nov. hoc loco (Figure 6A)
Figure 6
Photographs of the mesic grazed alpine pastures on fertile soils: (A) Papavero crocei-Ranunculetum popovii; (B) Littledaleo alaicae-Stipetum trichoidis; (C) Bistorto ellipticae-Pedicularietum korolkowii; (D) Poo attenuatae-Callianthemetum alatavici; (E) Poo bucharicae-Oxytropidetum savellanicae; (F) Drabo alajicae-Omalothecetum supinae and (G) Trollius komarovii and Erigeron heterochaeta community. All photographs were taken by A. Nowak.

Holotypus: Table S1, relevé number 189, holotypus hoc loco.
Diagnostic species: Carex alajica, Cerastium pusillum, Draba altaica, Leontopodium ochroleucum, Minuartia biflora, Papaver croceum, Potentilla flabellata, Ranunculus popovii, Schmalhausenia nidulans, Schulzia crinita, Taraxacum strizhoviae, Aspicilia fruticulosa
Constant species: Erigeron allochrous, Festuca alaica, Gastrolychnis uralensis, Koeleria spicata, Oxytropis immersa, Poa alpina, Poa litvinoviana
Dominant species: Carex alajica, Festuca alaica, Kobresia capilliformis, Kobresia persica, Leontopodium ochroleucum, Oxytropis immersa, Potentilla virgata, Ranunculus popovii, Trollius dschungaricus
Floristic and habitat characteristics: Papavero crocei-Ranunculetum popovii forms patchy alpine grasslands used as pastures at altitudes from 3,119 to 3,477 m a.s.l. (Figure 3A) on flattened areas of hilltops and passes of the Terskey Ala-Too range south of the Issyk-kul Lake (NE Kyrgyzstan) and the Alai range (SW Kyrgyzstan), as well as on the gentle slopes of the Kyrgyz Ala-Too range in the North Tian-Shan (N Kyrgyzstan). It prefers south-eastern exposures and inconsiderable slope inclinations of approximately 2° to 15° (Figure 3B). Both, species richness and cover of the herbaceous layer are relatively high. From eight to 27 species were recorded in sampled plots (mean = 20; Figure 3C). The cover of the herb layer ranged from 40 to 95% (average above 70%; Figure 3F). Noteworthy is the presence of mosses with a cover of up to 10% (Figure 3G).
13. Littledaleo alaicae-Stipetum trichoidis A. Nowak et al. 2018 (Figure 6B)
Diagnostic species: Androsace dasyphylla, Dracocephalum paulsenii, Gentianella turkestanorum, Gypsophila cephalotes, Hedysarum minjanense, Helictotrichon tianschanicum, Koeleria pyramidata, Potentilla pamiroalaica, Stipa trichoides, Taraxacum kovalevskiae
Constant species: Festuca alaica, Poa litvinoviana
Dominant species: Dracocephalum paulsenii, Festuca alaica, Hedysarum minjanense, Oxytropis lehmannii, Stipa trichoides
Floristic and habitat characteristics: Littledaleo alaicae-Stipetum trichoidis is a typical alpine sparse grassland used as grazing land. Patches of this association have been recorded in Tajikistan on the gentle slopes of the Peter the First ranges in the vicinity of Koshtegirman and Mok at elevations between 3,165 and 3,297 m a.s.l. (Figure 3A) with a gentle inclination of slopes from 2° to 15° (mean = 5.5°; Figure 3B), most often with a north-western exposure. The cover of the herb layer ranges from 60 to 85%, with a mean of 70% (Figure 3F). The species richness is fairly low, ranging from six to 13 species per plot (average 10; Figure 3C).
14. Bistorto ellipticae-Pedicularietum korolkowii S. Nowak et al. 2024 ass. nov. hoc loco (Figure 6C)
Holotypus: Table S1, relevé number 232, holotypus hoc loco.
Diagnostic species: Bistorta elliptica, Carex orbicularis, Cerastium pusillum, Crepis oreades, Draba subamplexicaulis, Dracocephalum nutans, Erigeron allochrous, E. tianschanicus, Euphorbia alatavica, Gentiana squarrosa, Koeleria spicata, Minuartia biflora, Oxytropis immersa, Pedicularis korolkowii, Poa alpina, Potentilla hololeuca, Primula algida, Rumex acetosa, Schulzia crinita
Constant species: Festuca alaica, Geranium saxatile, Leontopodium ochroleucum
Dominant species: Alchemilla tianschanica, Carex alajica, Festuca alaica, Festuca amblyodes, Lagotis ikonnikovii, Leontopodium ochroleucum, Poa alpina, Potentilla flabellata, Schulzia crinita
Floristic and habitat characteristics: Bistorto ellipticae-Pedicularietum korolkowii is an alpine grassland that serves as summer pasture mainly for sheep. This association has been recorded on gentle slopes of the Kyrgyz Ala-Too range (Chüy and Naryn regions in northern Kyrgyzstan) at heights ranging from 3,145 to 3,392 m a.s.l. (mean = 3,268 m a.s.l.; Figure 3A) with varying inclinations from 2° to 30° (mean = 10°; Figure 3B). This vegetation grows on slopes with different exposures, mainly north-east. The cover of the herb layer ranges from 60 to 90%, with a mean close to 80% (Figure 3F). The species richness is the highest compared to the other vegetation types, ranging from 16 to 30 species per plot (mean = 22; Figure 3C).
15. Poo attenuatae-Callianthemetum alatavici S. Nowak et al. 2024 ass. nov. hoc loco (Figure 6D)
Holotypus: Table S1, relevé number 266, holotypus hoc loco.
Diagnostic species: Bistorta vivipara, Callianthemum alatavicum, Festuca brachyphylla, Gagea michaelis, Gentiana algida, G. kirilowii, Hedysarum kirghisorum, Oxytropis trichocalycina, Poa attenuata, Ranunculus alberti, Thalictrum alpinum
Constant species: Carex orbicularis, Dichodon cerastoides, Eritrichium villosum, Koeleria spicata, Leontopodium ochroleucum
Dominant species: Bistorta vivipara, Carex alajica, Chrysosplenium nudicaule, Erigeron allochrous, Eritrichium villosum, Festuca alatavica, Gentiana algida, Kobresia persica, Koeleria spicata, Lagotis ikonnikovii, Poa attenuata, Potentilla flabellata, Ranunculus alberti
Floristic and habitat characteristics: Poo attenuatae-Callianthemetum alatavici can be classified as alpine snow-bed vegetation. This association has been sampled in north-eastern Kyrgyzstan in the Jeti-Ögüz district of the Terskey Ala-Too range at an altitude of 3,128 to 3,896 m a.s.l. (mean = 3,750 m a.s.l.; Figure 3A). It grows on gentle slopes or flatlands with an average inclination of approx. 9° (Figure 3B), most often with a south aspect. Plots comprised nine to 24 species, with a mean of 18 (Figure 3C). The cover of the herb layer ranges between 45 and 100% (mean = 80%; Figure 3F). Mosses form a distinct layer reaching up to 20% (Figure 3G).
16. Poo bucharicae-Oxytropidetum savellanicae S. Nowak et al. 2024 ass. nov. hoc loco (Figure 6E)
Holotypus: Table S1, relevé number 278, holotypus hoc loco.
Diagnostic species: Aconogonon hissaricum, Dichodon cerastoides, Oxytropis savellanica, Poa bucharica, P. hissarica, Ranunculus rubrocalyx, R. rufosepalus, Rheum fedtschenkoi, Taraxacum raikoviae
Constant species: Festuca alaica
Dominant species: Dichodon cerastoides, Kobresia persica, Oxytropis savellanica, Potentilla crantzii, Potentilla flabellata, Potentilla grisea
Floristic and habitat characteristics: Poo bucharicae-Oxytropidetum savellanicae is an alpine grassland used as a pasture mainly for sheep. The association has been recorded on gentle slopes and flattened areas in the Zeravshan Mts. (near Ayni) and the Hissar Mts. (near Kalon). Plots of this phytocoenosis occupy mostly north-eastern slopes with an inclination of 2° to 30° (approx. 10° on average; Figure 3B) at altitudes from 3,399 to 3,650 m a.s.l. (mean = 3,550 m a.s.l.; Figure 3A). The number of plant species ranges from six to 13 per plot, with a mean of 10 (Figure 3C). The cover of the herb layer is high, ranging between 55 and 95 species (mean = 75; Figure 3F).
17. Drabo alajicae-Omalothecetum supinae S. Nowak et al. 2024 ass. nov. hoc loco (Figure 6F)
Holotypus: Table S1, relevé number 284, holotypus hoc loco.
Diagnostic species: Aconogonon coriarium, Androsace dasyphylla, Carex turkestanica, Draba alajica, Erigeron cabulicus, Geranium collinum, Lagotis korolkowii, Omalotheca supina, Polygonum vvedenskyi, Potentilla crantzii
Constant species: Eritrichium villosum, Festuca alaica, Poa litvinoviana
Dominant species: Androsace dasyphylla, Carex turkestanica, Eritrichium villosum, Kobresia persica, Omalotheca supina, Potentilla crantzii, Sibbaldia olgae
Floristic and habitat characteristics: Drabo alajicae-Omalothecetum supinae forms an alpine short grassland that has been spotted in Tajikistan in the Peter the First Mts. near Koshtegirman and Mok at an altitude of 3,197 to 3,292 m a.s.l. (Figure 3A). This vegetation prefers the north-eastern aspect with a gentle inclination from 2° to 20° (mean = 13°; Figure 3B). The cover of the herb layer varies from 45 to 90% (mean = 74%; Figure 3F), and the species richness is moderate, ranging from nine to 17 species per plot (average 12; Figure 3D). This vegetation is used as summer pasture for grazing sheep.
18. Community of Trollius komarovii and Erigeron heterochaeta (Figure 6G)
Diagnostic species: Erigeron heterochaeta, Oxytropis gorbunovii, Scrophularia griffithii, Trollius komarovii
Constant species: Aster alpinus var. serpentimontanus, Carex orbicularis, Geranium saxatile, Ranunculus alberti
Dominant species: Caragana jubata, Carex orbicularis, Eritrichium villosum, Salix caesia
Floristic and habitat characteristics: The community of Trollius komarovii-Erigeron heterochaeta can be defined as alpine wet grassland. Patches of this phytocoenosis were found in north-eastern Kyrgyzstan in the Ak-Suu and Jeti-Ögüz districts on flatlands and the gentle slopes of the Terskey Ala-Too range east and south of Lake Issyk-kul, at altitudes of 2,615 to 3,842 m a.s.l. (Figure 3A). This vegetation develops mainly on the north-western slopes with a slight inclination of five to 15° (mean = 11°, Figure 3B) or flatlands. The mean cover of herbs is 45% (ranging from 30% to 80%; Figure 3F), and the richness of vascular species raging from seven to 29 species per plot (mean = 15; Figure 3D).
. Discussion
. Comparison of Pamir-Alai and western Tian Shan high mountain grasslands to the surrounding vegetation
Alpine and subnival grasslands create distinct zonal vegetation between 2,500 and 4,500 m a.s.l. along all mountain ranges from Pamir Alai in the southwest to Sayan Mts. in the northeast. They are often dominated by Achillea bucharica, Aconitum rotundifolium, Agrostis canina, Anemone protracta, Aster serpentimontanus, Calamagrostis alajica, Eritrichium villosum, Gagea jaeschkei, G. leucantha, G. olgae, G. setifolia, Hedysarum cisdarvasicum, Lagotis ikonnikovii, Lloydia serotina, Pedicularis sarawschanica, P. verae, and Tulipa turkestanica (Afanasjev, 1956; Sidorenko, 1971; Swacha et al., 2023; Świerszcz et al., 2023). In the recent work on alpine grasslands in Middle Asia (Swacha et al., 2023; Świerszcz et al., 2023), 13 plant communities were distinguished within three alliances (Aconito rotundifolii-Potentilion pamiroalaicae, Stipo regelianae-Ptilagrostion malyschevii and Artemision dracunculi). They are mainly adjacent to forb steppes in lower alpine and subalpine locations or mountain desert steppes or cryophilous steppes in lower rainfall zones, e.g., the Eastern Pamir (A. Nowak et al., 2021, 2023). Several grassland communities have been recorded in the alpine belt of Altai Mts. of south-eastern Kazakhstan (Rubtsov, 1948). They were included in high-altitude vegetation and called ‘low-herb alpine grasslands’ following the Russian classification approach. These communities are characterized by the presence of species such as Bistorta vivipara, B. elliptica, Primula algida, Festuca kryloviana, Poa alpina, Geranium saxatile, Trisetum altaicum, T. spicatum, etc. In addition, Kobresia-forb swards (with Kobresia capilliformis, K. humilis, Alchemilla retropilosa and Leontopodium fedtschenkoanum) and rupiculous alpine vegetation on fine debris (with Dracocephalum imberbe, Saussurea glacialis, Rhodiola coccinea, Potentilla biflora, Oxygraphis glacialis and Trollius altaicus) have been reported from Altai Mts. (Dimeyeva et al., 2016; Rubtsov, 1948). The majority of these species also occur in western Middle Asia. In our opinion, these communities are very similar to the alpine and subnival vegetation types in Tajikistan and Kyrgyzstan. For example, communities dominated by Dracocephalum imberbe, Poa alpina, Geranium saxatile, or Festuca alaica. Therefore, we decided to merge the high-altitude grasslands of western Middle Asia (Pamir-Alai, W Tian Shan) and Central Asia (Altai) into one class; however, this requires further comparative studies to reach the final consistency. Such a pattern of distribution shows that the forb steppe vegetation (dominated by Irano-Turanian plants), approaches from the south-west into the mountains of Middle Asia up to the subalpine belt, while from the opposite direction, from the north-east, the temperate climate vegetation (Euro-Siberian) forming alpine and subalpine grasslands “overlays” the alpine and subnival belts.
However, towards the south, especially in the drier areas surrounding the eastern Pamir (e.g., Alaian, Zaalaian, Fergana, Shugnan Mts.), this general SW-NE pattern changes somewhat along the phytogeographical borderline between Middle and Central Asia. Rainfall values are decreasing, resulting in the fact that, despite very high altitudes, species of Irano-Turanian origin dominate. Characteristic ‘dry alpine grasslands’ are formed (e.g., Dracocephaletum imberbe or Nepeto pamiricae-Rhodioletum recticaulis), or cushion communities (e.g., Acantholimonetum diapensioidis) which have low biomass production and sparse cover. They are similar to the high alpine rupiculous vegetation of Alborz Mts. in terms of functional, life-form, and often taxonomic composition (Oxytropido persicae- Cousinietum multilobae; Noroozi et al., 2010). In this case, combining these vegetation types in one large unit at the class rank seems reasonable (Klein, 1982; Ukacheva, 1975). However, given that our dataset contains little data corresponding to low, patchy alpine grasslands composed mainly of Oxytropis immersa (high share only in Papavero crocei-Ranunculetum popovii), we decided not to follow Ukacheva (1975) and validate the unit of Oxytropeta immerse but utilize the class proposed by Noroozi et al. (2014) – Didymophyso aucheri-Dracocephaletea aucheri. Being typical Middle Asian subnival vegetation, these communities inhabit rupiculous and xerophytic habitats at approximately 3,500–4,500 m a.s.l. They are composed of many common taxa, particularly at the genus rank, with Astragalus, Alchemilla, Allium, Cousinia, Didymophysa, Nepeta, Oxytropis, Potentilla, Silene, and Veronica as the richest genera (Klein, 1982; Noroozi et al., 2020). In particular, the snow-bed vegetation (Taraxaco brevirostris-Polygonion serpyllacei) in northern Iran shares a considerable proportion of common taxa with Middle Asia with Festuca alaica, Oxytropis savellanica, Kobresia schoenoides, Oxyria digyna, and Gnaphalium supinum as the most prominent. In addition, the name-giving species of this group (Polygonum serpyllaceum and Taraxacum brevirostre) are common in the highlands of Pamir-Alai (A. Nowak et al., 2020). T. brevirostris is replaced or supplemented by closely related species, such as T. minutilobum, T. pseudobrevirostre, T. kovalevskiae, T. schugnanicum, and T. murgabicum, which make the physiognomy of the vegetation very similar. Although, with lower rainfall compared to NW Iran, especially in the Pamir, the snow-bed vegetation of the Pamir Alai is definitely more rupiculous in character, probably more similar to those from the drier subnival belt of the Zagros Mts. where communities with Polygonum serpyllaceum were also noticed (Noroozi et al., 2020). The same applies to the rupiculous habitats of the subnival belt, where among the most common taxa in the Alborz Mts. are those also known from Middle Asian highest elevations (e.g., Didymophysa, Draba, Dracocephalum, Erysimum, Lamium, Oxytropis, Paraquilegia; Noroozi et al., 2020).
The relationship between the subnival vegetation of Middle Asia and the class Rhodioletea described in Altai is challenging to resolve. It is defined as rupiculous high-altitude sparse vegetation on subnival screes with Rhodiola algida, R. coccinea, R. quadrifida, R. rosea, Chamaenerion latifolium, Gastrolychnis apetala, Poa alpigena, Saxifraga sibirica, S. macrocalyx, S. cernua, S. oppositifolia, Cerastium lithospermifolium, Primula nivalis, and Trisetum mongolicum (Rhodioletalia quadrifidae Hilbig, 2000; Korolyuk, 2001). The description included in this work matches the physiognomy of the vegetation and habitat conditions in many patches of high mountain communities on fine stony screes with a thin soil layer in Pamir-Alai and western Tian Shan. Particulary in Achoriphragmetum pinnatifidi, the frequency of R. coccinea is relatively high. Other species from this genus (R. recticaulis, R. gelida, R. heterodonta and R. pamiroalaica) are also included in our dataset, but rather contributing as non-dominant compositional elements. It is worth noting that the genus Rhodiola has its main center of distribution in the high mountains of Middle and Central Asia, Hengduan Mts., and the Qinghai-Tibetan Plateau (Zhang et al., 2014). It seems necessary to carry out further studies focussing on patches of vegetation with a high proportion of species of the genus Rhodiola in order to define its range in the western part of Middle Asia.
The close geographical proximity and landscape similarities with a large area of alpine and subnival belts means that the described vegetation has much in common with the high mountain communities of the northern Hindu Kush. It shares numerous alpine species with the Pamirian plateau, including Waldheimia tridactylites, Saussurea glacialis, Aulacospermum stylosum, Psychrogeton andryaloides, Saussurea gnaphalodes, Nepeta pamiriense, Leontopodium ochroleucum, Chorispora macropoda, Draba altaica, Alopecurus himalaicus, Pentaphylloides dryadanthoides, Potentilla gelida, and Acantholimon diapensioides (Breckle & Rafiqpoor, 2020).
In the Karakorum and the western Himalayan range, in Eastern Ladakh, the Kobresia royleana–Kobresia schoenoides matts, and Poa attenuata–Potentilla pamirica grasslands have been reported from elevations up to 5,700 m. (Dvorskỳ et al., 2011). They are similar to high mountain matts in physiognomic terms but also in terms of species richness, composition, and vegetation structure. Perhaps the only difference is that the matts of the western Himalayas are mainly grazed by yaks, while in the western Tian Shan they are mainly grazed by horses.
. Variability and internal relations of high alpine and subniwal vegetation of Middle Asia
The vegetation of the highest mountain altitudes in Middle Asia is very diverse and, to some extent, heterogeneous. This is caused by a diverse, harsh environment that is characterized by extremely high-temperature amplitudes and the lowest minimum temperature in winter up to −60 °C, and also by high solar radiation, particularly UV-B radiation that can be 100% more intense than in the valleys (see Leuschner & Ellenberg, 2017). Moreover, the daily frost, even in summer, scarce soils, low nutrient availability combined with solifluction, long-lasting and deep snow cover, slabs, avalanches, landslides, and strong winds make this area one of the most unfavorable for plant life. This extremely harsh environment is responsible for controlling and filtering the species that need to adapt to these extremes despite the type of substrate that is not so crucial (e.g., calcicole or calcifuge). Therefore, it is often difficult to distinguish between scree habitats, chasmophytic ledges, and desert steppes at very high elevations. Some associations can occupy two or even three different habitat types (e.g., Dracocephaletum imberbe), but have an optimal habitat only in one. This causes considerable difficulty in distinguishing between these plant communities. However, with certainty, the high mountain vegetation of Middle Asia can be divided into 3-4 main groups. The first is subnival rupiculous vegetation, including Violo acutifoliae-Potentilletum biflorae and Dracocephaletum imberbe in central Tian Shan, Carici stenocarpae-Androsacetum akbaitalensis, Achoriphragmetum pinnatifidi, Leiospora bellidifolia-Ajania trilobata comm., Nepetetum longibracteatae, and Nepeto pamiricae-Rhodioletum recticaulis in the Pamir and Alai ranges. Additionally, into this group was classified the Dracocephaletum nodulosi-Rosularietum borissovae from the lower alpine belt of the central Tian Shan and Oxyria digyna community, which occupies snow beds. Patches of this community are widespread in the Eastern Pamir and inhabit wet snow beds but also gentle slopes with a well-hydrated soil profile. Towards the drier sites, the Acantholimon diapensiodes community gives way to typical cushion-tragacanthic vegetation growing on underdeveloped soils with Acantholimon spp. or Oxytropis echidna. This cushion-tragacanthic vegetation often has a patchy physiognomy and occurs in a mosaic with cryophytic desert steppes formed by Stipa orientalis and S. glareosa.
In places with a more nutrient-rich habitat and slightly better-developed and hydrated soils, graminoid matts dominate on gentle slopes, passes, and hilltops. Generally, in Middle Asia, they are composed mainly of species of the genus Kobresia. They form dense and widespread matts with dominance of species such as S. regeliana as well as Ptilagrostis malyschevii and Stipa tremula (the two later species replace here P. dichotoma and S. purpurea that occur in Himalaya and Tibet; Nobis et al., 2022), that is present in our dataset and forms a distinct Violo tianschanicae-Stipetum regelianae.
Another group is formed by vegetation of slightly lower altitudes, which belong to typical alpine grasslands (Swacha et al., 2023). They include several distinct groups and occur mainly in the Tian Shan ranges. Some of these communities have sparse physiognomy of graminoid swards (Papavero crocei-Ranunculetum popovii, Littledaleo alaicae-Stipetum trichoidis, Poo bucharicae-Oxytropidetum savellanicae), whereas other form relatively dense carpets and resemble species rich, compacted short-grass vegetation of the alpine zone (Bistorto ellipticae-Pedicularietum korolkowii, Trollius komarovii-Erigeron heterochaeta community). Two communities in this group are associated with wetter habitats that occur adjacent to moraines‘ foot, flat snow beds, or sloping water outlets. These are Poo attenuatae-Callianthemetum alatavici and Drabo alajicae-Omalothecetum supinae. It cannot be ruled out that the syntaxonomic position of the latter two vegetation types will change in the future after collecting more data.
. Conclusions
Our study has expanded the knowledge of subnival snow bed, grassland, and rupiculous vegetation in the Pamir-Alai and western Tian Shan Mts. and contributed to the consistent hierarchical classification of vegetation in the region (A. Nowak et al., 2016a; Swacha et al., 2023; Świerszcz et al., 2023). Similarly to the vegetation of screes, cryophilous or forb steppes, or alpine grasslands (A. Nowak et al., 2016b, 2021; Świerszcz et al., 2023), the heterogeneity of this subnival vegetation is evident. The Central Tian Shan is strongly influenced by a temperate climate and Euro-Siberian flora (thus the solid relation for Altai vegetation), while the southernmost limits of the eastern Pamir are compositionally related to the Irano-Turanian region. An additional strong gradient that influences the diversity of subnival and alpine vegetation of Central and Middle Asia is the humidity, which decreases from the snow bed, moraines, through mountain slopes with moderate moisture content to rupiculous and almost semi-desert plateaus in the Eastern Pamir, which are arid. Grasslands are additionally influenced by different grazing intensities, which is a crucial factor throughout Middle Asia. It has the greatest impact on alpine grasslands and isis much less significanton low-productive vegetation at the highest altitudes.
This study summarizes our grassland surveys in Middle Asia; however, it would still be helpful to conduct further surveys, especially in neighboring regions such as the Hindu Kush, Kopet-dagh, and Altai Mts., to obtain a stable and internally consistent syntaxonomical system of grassland communities.
. Supplementary material
The following supplementary materials are available for this article:
Table S1. The analytic table of the high alpine and subnival vegetation of the Pamir-Alai and western Tian Shan Mts.
Table S2. Synoptic table of the high alpine and subnival vegetation of the Pamir-Alai and western Tian Shan Mts. The phi coefficient values (in superscript) in the table are multiplied by 100. Main values are species frequencies (in percentages).
Tables S1 and S2 are available in the repository Figshare: https://doi.org/10.6084/m9.figshare.22758647.v2
