. Introduction
The alpine belt, commonly defined as high-altitude areas above the climatic treeline, is characterized by a high micro-habitat variability and a wide range of environmental conditions. This results in a considerable species richness and vegetation diversity. Many global alpine regions are considered as biodiversity and endemism hotspots and are inhabited by substantial numbers of endemic species (Körner, 2003; Nagy et al., 2003; Spehn et al., 2010). Pamirian vegetation was found to be one of the most diverse and species rich at the global scale (Testolin et al., 2020). At the same time, alpine habitats, such as meadows, pastures, wind-swept swards, and steppes are ecosystems threatened by land use (Körner, 2003; Nagy & Grabherr, 2009) and anthropogenic climate change (Hughes, 2000; Körner, 2003; Spehn et al., 2010). Alpine biomes cover 3.56 Mkm2 or 2.64% of land outside Antarctica (Testolin et al., 2020). Despite temperature differences across latitude, these ecosystems converge below a threshold of ca. 6 °C mean annual temperature and are differentiated by seasonality and continentality. Alpine vegetation in the Pamir Mountains is important for household and pastoral economy throughout Middle Asia (Kerven et al., 2011).
Although rocks are not the most species rich among mountain habitats, the chasmophytic vegetation is considered very unique and interesting. Owing to the harsh and extreme conditions, such habitats could serve as suitable biotopes for many endemic and specialized plant species (e.g., Agakhanjanz & Breckle, 1995, 2002; Kazakis et al., 2007; Médail & Verlaque, 1997). The extreme uniqueness of petrophytic flora is reflected in many phytosociological surveys focused on rupiculous vegetation which have been conducted in mountainous areas of Europe and Asia. Vegetation of saxatile habitats has been continuously studied in the Carpathians (e.g., Sanda et al., 2008; Valachovič, 1995), the Alps (e.g., Pott, 1995), the Mediterranean (Deil et al., 2008; Dimopoulos et al., 1997; Pesaresi et al., 2014; Terzi & Jasprica, 2020; Terzi et al., 2018), Crimea (Ryff, 2016) and have been summarized in the work of Mucina et al. (2016). In Africa, few studies concerning rupicolous vegetation have been published (e.g., Deil & Hammoumi, 1997). In Asia, the Taurus (Eren et al., 2004; Hein et al., 1998; Parolly, 2015), Caucasus (Belonovskaia, 2012; Ermolaeva, 2007; Golub et al., 2009), ranges of northern Iran (Klein, 1987, 1988; Naqinezhad & Esmailpoor, 2017; Noroozi et al., 2014) and Altai (Ermakov et al., 2006) have been investigated. In recent years, the mountain ranges of Tajikistan have been extensively studied (Nowak et al., 2011, 2020).
Located between the continental Asian deserts in the west and east, the great mountain ranges of Tian Shan in the north and Kunlun, Hindu-Kush, and Karakorum in the east and south, respectively, and having all vertical belts from hot to permafrost deserts, Tajikistan offers an outstanding range of biotopes for plants and vegetation. Chasmophytic habitats in the country, probably because of their high degree of isolation and position within the borderland of different phytogeographical regions, are particularly diverse. They harbor more than 35% of the country’s vascular plant flora and act as a refuge for many narrowly distributed plant species. More than half of them inhabit scree and rocky habitats. Scree phytocoenoses also harbor the most numerous representation of exclusive endemics of this territory (Nowak et al., 2011; Nowak, Świerszcz, Nowak, & Nobis, 2021).
Research on rock vegetation in Tajikistan, despite recent advancements (Nobis et al., 2013; Nowak, Nobis, et al., 2014; Nowak, Nowak, et al., 2014a, 2014b, 2014c; Nowak, Świerszcz, Nowak, Nobis, et al., 2021), is still far from being completed. Owing to significant differences in habitat conditions (inclination, insolation, crevice size, altitudinal amplitude, soil availability) and species composition, two main groups of phytocoenoses have been distinguished within the chasmophytic vegetation representing the Campanuletalia incanescentis order in Middle Asia: Asperulo-Poion (with two suballiances, Campanulenion lehmannianae and Pentanemenion albertoregeliae) and Caricion koshevnikovii that include the communities of colline and montane belts in western Pamir-Alai (Nowak, Nobis, et al., 2014; Nowak, Nowak, et al., 2014a). Within the alpine petrophytic vegetation in tiny fissures and on solid rock faces in higher altitudes (alpine and subnival) a number of associations have been distinguished, e.g., Sergietum regelii, Scutellarietum rubromaculatae, Scutellarietum orbicularis, Campanuletum lehmannianae, Achoriphragmetum turkestanici, Andrachnetum fedtschenkoi, Nanorrhinetum ramosissimi, Scutellarietum baldshuanicae, Minuartio litwinowii-Phaeonychietum surculosi, and Silenetum samarcandensis. These were included in the Campanuletalia incanescentis order and the Asperulo albiflorae-Poion relaxae alliance (Nobis et al., 2013; Nowak, Nobis, et al., 2014). For habitats located at lower elevations in warm, Irano-Turanian climate, an additional eight associations were proposed: Scutellarietum hissaricae, Scutellarietum schugnanicae, Scutellarietum zaprjagaevii, Scutellarietum baldshuanicae, Tylospermetum lignosae, Dionysietum involucratae, Nanorrhinetum ramosissimi, and Campanuletum albertii (Nowak, Nobis, et al., 2014; Nowak, Nowak, et al., 2014a).
A geobotanical survey aimed at providing a detailed description of petrophytic vegetation was initiated in western Tajikistan in 2008 in both rock and scree habitats. Until now, the vegetation of these habitats in Pamir-Alai and Tian Shan has been partially studied within various elevational belts mainly in western Tajikistan (e.g., Nowak, Nobis, et al., 2014; Nowak, Nowak, et al., 2014a, 2014c; Nowak et al., 2016). However, the area of eastern Pamir and the relation of Pamir-Alai and Tian Shan colline and the classification, phytogeography, and ecology of montane vegetation are still insufficiently investigated.
Continuing the exploration of the chasmophytic habitats of Middle Asia, we present the diversity of rock vegetation in the eastern Pamirs and SW Tian Shan in the alpine belt (approx. 1,850–4,000 m a.s.l.). Climatically and phytogeographically, this area is fundamentally different from the Western Pamiro-Alai, and its alpine belt vegetation has not yet been explored. Therefore, the purpose of this paper is to identify the main types of chasmophytic vegetation inhabiting crevices, fissures, rock ledges, and clefts. To stabilize the classification and to complement the survey, we also included the southern outskirts of Pamir-Alai and Western Pamir areas in our research. Using data available from our phytosociological research, we have attempted to address the following questions: (i) What is the diversity of the rock vegetation of alpine belts in the Pamir and western Tian Shan? (ii) What is the relation of petrophytic vegetation pattern to environmental variables in the studied area? (iii) What is the relationship of alpine petrophytic vegetation recognized in the study area with that previously observed in other regions of Europe and Asia?
. Material and Methods
. Study Area
The vegetation survey was conducted in the eastern part of Middle Asia, i.e., the Pamir Mountains (eastern Tajikistan) and western Tian Shan Mountains (Kyrgyzstan) within an area of ca. 150,000 km2 (Figure 1). Owing to considerable phytogeographical differences between the Pamir-Alai and Tian Shan Mountains, scree vegetation of both regions (Alai, Transalaian, Alichurian, Shachdarian, Shugnan, Sarikol, Yazgulem, and Peter the First ranges in Pamir-Alai; and Trans-Ili Alatau, Kyungey Ala-Too, Terskey Ala-Too, Songkol, Fergana, Kyrgyz, and Chatkal mountains in Kyrgyzstan) were included in the study. The stony and bare character of the highland landscapes of eastern Middle Asia makes this territory particularly suitable for different types of chasmophytic communities. The vast rupiculous habitats of eastern Middle Asia extend across a long elevation gradient. The studied sites were located between 1,000 and 4,100 m a.s.l. (mean 2,700) and differ in terms of aspect, inclination, bedrock, and particle size.
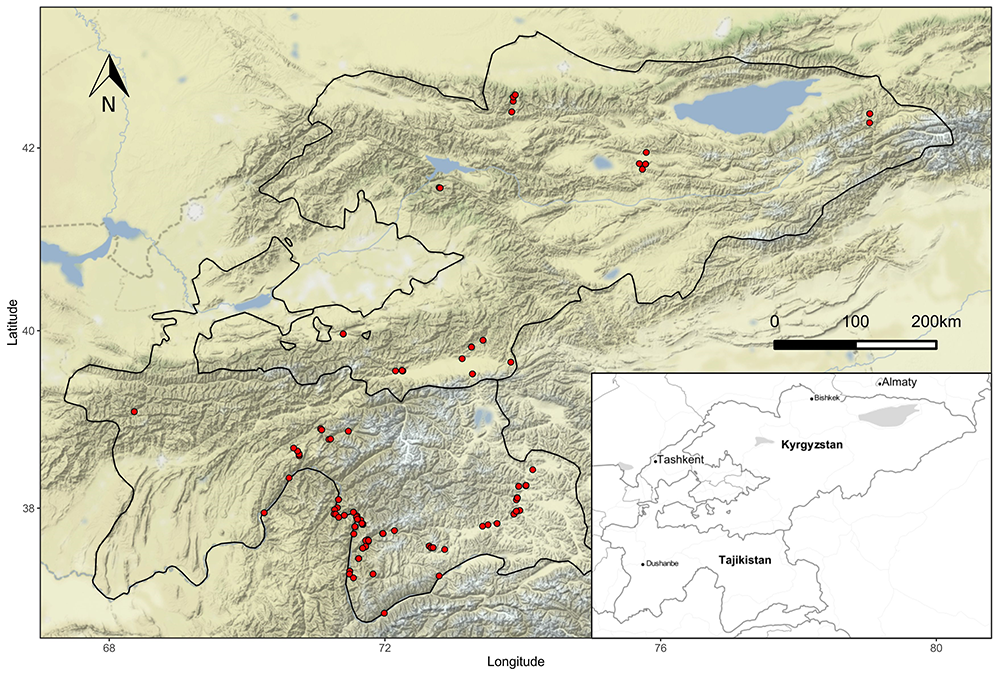
Variable orography and location between the main bioclimatic zones of the Irano-Turanian and Central Asiatic regions (with influences of Indo-Indochinese climate from the south and Euro-Siberian from the north) make the climatic conditions of the studied area difficult to characterize. The study area is located within a transition zone between the temperate and Irano-Turanian types of macrobioclimates. The first one is characterized by a summer precipitation peak and lower continentalism, whereas for the second, winter rainfall is typical (Djamali et al., 2012). There are four main types of climatic regions within the study area (Latypova, 1968; Safarov, 2003; Vladimirova, 1968):
The warm, continental, Irano-Turanian climate in the Fergana Basin. The surroundings of Jalalabad and Osh are characterized by winter precipitation with a peak of up to 80 mm in March and a yearly average of ca. 200–250 mm. The temperature reaches 20 °C in April and rises to an average of 34 °C in June–August. Precipitation is scarce during these months, and does not exceed 10 mm per month. Snow and frost occur from December to February with extreme values that can reach −27 °C; however, the average temperature does not decrease below −3 °C.
The warm, humid, continental climate in the Tian Shan and Pamir-Alai ranges. In the colline and montane zones of the ranges, the average temperature in June is around 22 °C and drops to 10 °C in the alpine belt. Annual precipitation here ranges from about 500 mm on the northern slopes to ca. 1,000 mm on the southern ones.
The cold semiarid climates of the Issik-Kul basin, central and western parts of the Alai Valley as well as foothills and plateaus at the colline, montane, and subalpine belts. These areas are clearly distinguished by low precipitation with an average ca. 200–400 mm per year. The distribution of the rainfall during the year is similar to that in the temperate climate, with a maximum in May–July (up to 70 mm). The temperatures exceed 20 °C only in summer with the annual average ca. 10 °C.
The cold desert climate of the easternmost sections of the Alai Valley and eastern Pamirian Plateau. Unlike the west Pamir or Tian Shan ranges, this area is distinguished by significant aridity with less than 100 mm mean annual precipitation. Only in May and August does the average monthly rainfall exceed 20 mm. The average yearly temperature slightly exceeds 0 °C with a minimum below −30 °C in January and February.
Numerous local anomalies caused by orography, wind circulation, and altitudinal differences occur within all zones mentioned above.
. Data Sampling and Data Analyses
For six successive vegetation seasons from 2013–2019, 237 relevés were collected in rock fissures, ledges, and clefts in the alpine belt of the Pamir and south-western Tian Shan mountains. The size of each vegetation plot was 1 m2. In each plot, all vascular plant and cryptogam species were recorded using the 7-degree cover-abundance scale of Braun-Blanquet (Westhoff & van der Maarel, 1973). We covered a broad range of habitats located across a wide altitudinal range, various size of rock debris, exposition, and inclination. Exposition was defined as the compass direction that the slope faced (N, S, E, W, NE, NW, SE, and SW) determined by using a Suunto compass and inclination was measured visually. For each plot, geographical coordinates and altitude were measured using a GPSMAP 60CSx device with an accuracy of ±10 m, and the WGS84 reference frame. In the tables, latitude and longitude are given in decimal scale. Rock type was determined by a professional geologist (see Acknowledgments). Rock samples were identified by analysis of lithology, pore geometry, mineralogical components, texture, permeability, hardness, and pH. For the statistical analyses, we used climatic data (mean annual temperature and sum of annual precipitation) that were extracted from the CHELSA database version 1.2 at resolution of 30 arc sec (ca. 1 km2; http://chelsa-climate.org/; Karger et al., 2017). The CHELSA dataset was used because it contains data averaged over the period 1979–2013 and it was not feasible for this study to measure these climatic variables in the field (e.g., using data loggers).
All collected data were stored in the Vegetation of Middle Asia database (Nowak et al., 2017) and analyzed in R (R Core Team, 2020) and JUICE software (Tichý, 2002). A modified TWlNSPAN analysis (Hill, 1979; Roleček et al., 2009) provided an initial idea of data structure and resolution. The cover-abundance scale was transformed using the 4-step interval scale with cut-off levels at 0%, 5%, and 25%. As the plots were selected fairly objectively, we downweighted rare species using the chord distance as a measure of cluster heterogeneity (Roleček et al., 2009). Taxa identified only at the genus level were omitted during the analysis. Diagnostic species were identified using the phi coefficient as a fidelity measure (Chytrý & Tichý, 2003). Group size was standardized, and the Fisher exact test (p < 0.05) was applied. Species with a phi coefficient higher than 0.20 were considered diagnostic for a particular cluster. Taxa diagnostic for alliances were defined as those with a phi coefficient ≥0.15 in at least two clusters within this alliance (with the exception of one cluster which is very distinct). Species with a frequency higher than 40% were defined as constant and those with a maximum cover value exceeding 20% were defined as the dominant species of an individual cluster (plant community). For translation of the TWINSPAN results into phytosociological associations, we chose the highest division that still yielded floristically well-characterized terminal clusters with their own diagnostic species (Dengler et al., 2005; Michl et al., 2010). These terminal clusters were considered as associations or plant communities depending on the geographical range, certainty of taxonomic status of the diagnostic species, and the International Code of Phytosociological Nomenclature (ICPN) recommendations. During the division, the habitat profile and authors’ field experience were used to find comprehensive and ecologically interpretable results of classification.
To check the floristic-sociological classification and to highlight the relationships between relevés and species, nonmetric multidimensional scaling (NMDS) was performed (no downweighting of rare species, response data were log-transformed; Figure 3). Species cover values of the 7-degree Braun-Blanquet scale were transformed to a percentage scale (r, +, 1, 2, 3, 4, 5 to 0.1, 1, 5, 15, 37.5, 62.5, and 87.5 respectively). The differences between groups in environmental factors (altitude, temperature, precipitation, and inclination) and vegetation variables (cover herb and moss layer, species richness, and Shanon diversity index) were assessed using Kruskal–Wallis rank sum test (function kruskal.test) with multiple comparison based on Dunn test using the function dunnTest in the FSA package (Ogle et al., 2018) in R.
A data set with the constancy degree of all species is given in Table S2. Only species reaching constancy not less than 20% in at least one column are shown. For newly described associations, the ICPN was adhered to (Theurillat et al., 2021). According to the fourth version of the Code (Theurillat et al., 2021), we assigned the relevé type with reference to the supplementary materials (Table S1). All mentioned syntaxa are arranged into a syntaxonomic overview at the beginning of the description in the Results section. As the most significant attributes of the habitat, we considered spatial structure and environmental characteristics, mainly the elevation, crevice size, and bedrock type.
Species nomenclature mainly followed The Plant List (http://www.theplantlist.org/) and in some cases Cherepanov (1995). Plant material collected during field studies was deposited in the Herbarium of Middle Asia Mountains, hosted in OPUN (Opole University, Poland) and in KRA (Jagiellonian University, Poland). Supplementary tables (Table S1, Table S2) and figures (Figure S1, Figure S2) are available in Figshare Digital Repository (Nowak, Świerszcz, Nowak, Plášek, et al., 2021).
. Results
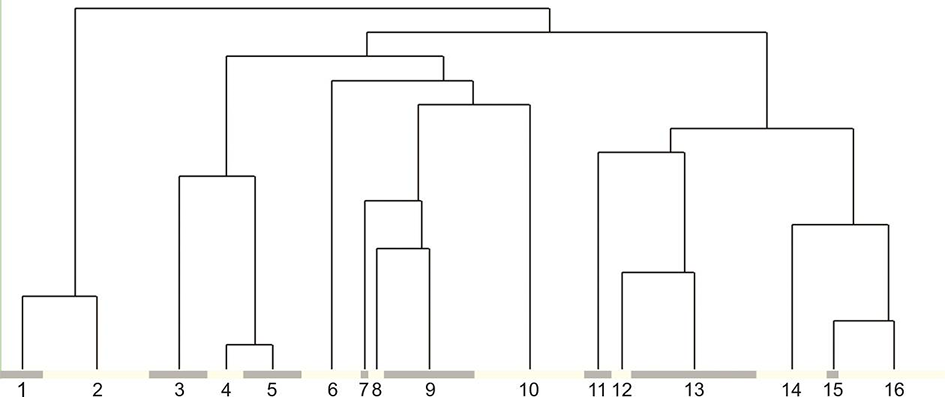
The TWINSPAN classification reveals five main groups of plant communities that can be distinguished in the alpine belt of the eastern Pamir-Alai and south-western Tian Shan (Figure 2; see synopsis of syntaxa). Group I comprises rocky plant communities from the Eritrichion pamirico-subjacquemontii alliance, distribution of which is determined mainly by harsh, semiarid climate of high eastern Pamir. Within Group II, the communities of petrophytic vegetation of rock ledges and clefts of highly elevated semiarid zone of the southern Pamir were gathered (Figure 2). Group III comprises the petrophytic vegetation of the alpine and upper montane belts of the subhumid zone in the Darvaz, Western Pamir, and south-western Tian Shan. The last two groups are included in one alliance, Asperulo albiflorae-Poion relaxae, as they exhibit similar species composition, preference to solid rock faces, and structural resemblance. However, Group IV shows considerable differences as it comprises plant communities that prefer more thermophilic habitats. It occurs in rock crevices in the alpine belt of the thermophilous south-western Pamir and was included in the Hippolytion darvasicae alliance. This vegetation often develops in shadowed sites, in deep fissures, or ledges with fairly high water availability. Still warm, but much drier are the habitats of Group V that is represented by three plant communities distributed in southeastern Pamir in the arid borderland close to Afghanistan and Vakhan Corridor.
Figure 2
Dendrogram illustrating the assignment of relevé groups identified by TWINSPAN to particular syntaxonomical units: 1 – community of Androsace villosa var. dasyphylla; 2 – Crepidifolio tenuifolii-Stipetum tianschanicae; 3 – community of Potentilla malacotricha and Waldheimia glabra; 4 – Parryetum schugnanae; 5 – Corydalideto bucharicae-Hippolytietum schugnanicae; 6 – Drabetum darvasicae; 7 – community of Erigeron vicarius and Ephedra regeliana; 8 – Paraquilegietum anemonoidis; 9 – Psychrogetonetum andryaloidetis; 10 – community of Campanula albertii; 11 – Silenetum kuhistanicae; 12 – community of Silene schugnanica; 13 – Scutellarietum schugnanicae; 14 – Acantholimonetum alexeenkoani; 15 – community of Rosularia alpestris; 16 – Inuletum schmalhausenii.
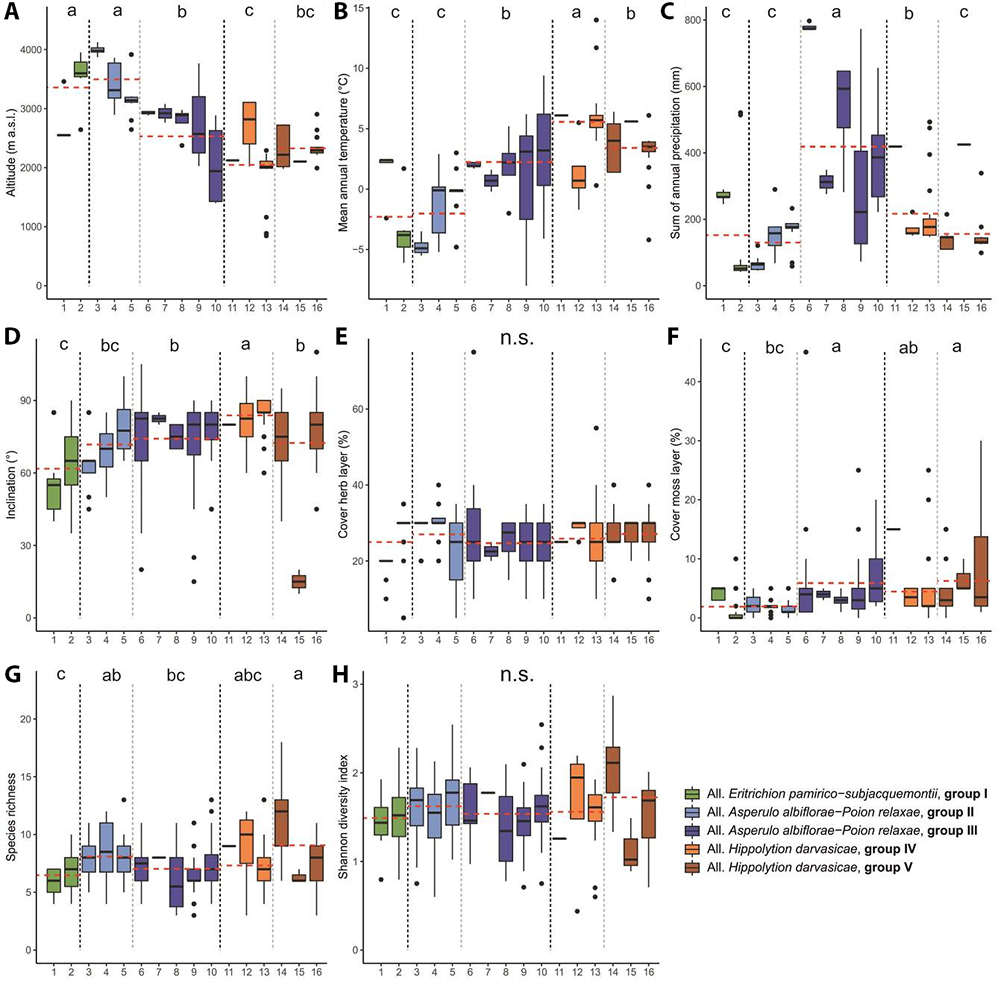
Similar results were obtained from NMDS analysis (Figure 3). Kruskal–Wallis test showed significant differences between analyzed groups according to altitude, mean annual temperature, precipitation, inclination, cover of moss layer, and species richness (Table 1). Mean values of these parameters with post hoc comparisons are presented on Figure 4. The analyses show three groups of samples with the additional two side small groups being internally fairly homogenous and determined mainly by climate and geography. However, within the heterogenous central group representing the Asperulo albiflorae-Poion relaxae alliance two subgroups may be distinguished. The upper one marked with dotted line represents the communities of the warmer zone of the Pamir that is to some extent similar to the samples from the left-side group (Clusters 10–13, Figure 3). The chasmophytic vegetation of Corydalideto bucharicae-Hippolytietum schugnanae and Parryetum schugnanae located in the central group occupy warmer habitats on sunny, southern slopes and walls (Figure 4B). Samples located in the lower part of the group correspond to communities of northern exposition like Psychrogetonetum andryaloidetis or at higher latitudes such as communities of Campanula albertii. The temperature gradient along the second axis of the NMDS applies to all groups shown in the graph. In the group of Pamirian chasmophytic vegetation (left side of the graph) it refers clearly to Inuletum schmalhausenii and Acantholimonetum alexeenkoani. However, it might explain to some extent the position (right part of the graph) of the Eastern Pamir highland group – Crepidifolio-Stipetum tianschanicae.Nevertheless, these Pamirian communities affiliated to the Eritrichion pamirico-subjacquemontii alliance are clearly distinct and reflect the profound difference in climatic conditions between Montano-Middle Asian (Afghano-Turanian) vs. Pamiro-Tibetan subregions (Kamelin, 2010). The first axis is undoubtedly related to altitudinal gradient. On the left-hand side of the graph (Figure 3), communities occurring in the upper montane and low alpine belts are centered, and on the right-hand side, there are samples of the highest, sometimes clearly exceeding 4,000 m a.s.l. sites.
Figure 3
NMDS ordination of chasmophytic vegetation in the study area. Note that in this analysis groups with small number of relevés classified as communities were omitted. Abbreviations: 1 – community of Androsace villosa var. dasyphylla; 2 – Crepidifolio tenuifolii-Stipetum tianschanicae; 3 – community of Potentilla malacotricha and Waldheimia glabra; 4 – Parryetum schugnanae; 5 – Corydalideto bucharicae-Hippolytietum schugnanicae; 6 – Drabetum darvasicae; 7 – Paraquilegietum anemonoidis; 8 – Psychrogetonetum andryaloidetis; 9 – community of Campanula albertii; 10 – Scutellarietum schugnanicae; 11 – Acantholimonetum alexeenkoani; 12 – community of Rosularia alpestris; 13 – Inuletum schmalhausenii.
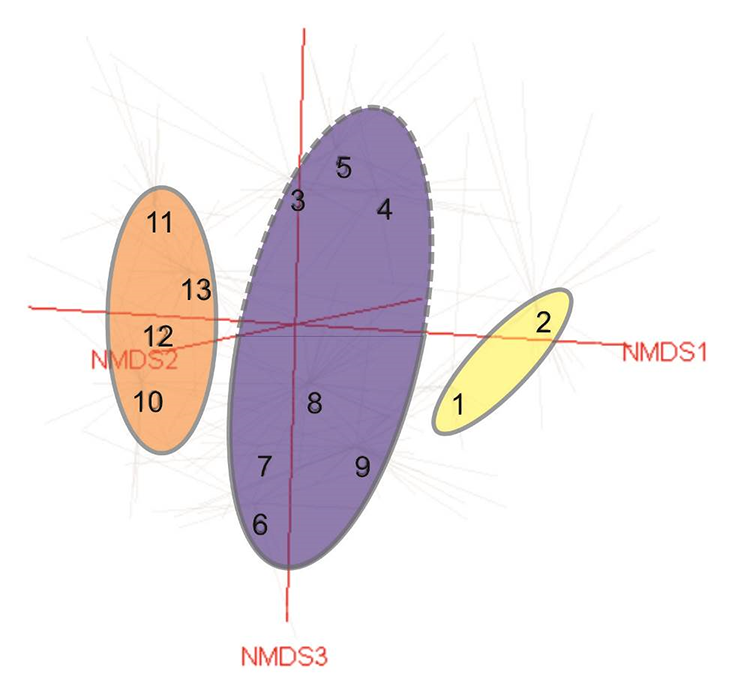
Table 1
Results of Kruskal–Wallis test for environmental and vegetation variables between identified syntaxonomical units of chasmophytic vegetation.
Figure 4
Boxplots showing median (line), quartiles, outliers and the range of (A) altitude, (B) mean annual temperature, (C) sum of annual precipitation, (D) inclination, (E) cover of herb layer, (F) cover of moss layer, (G) species richness, (H) Shannon diversity index for particular syntaxonomical units. Red line indicates mean values of vegetation groups. The values of χ 2 and p for statistical tests for vegetation groups are shown in Table 1. Different letters indicate significant differences among the vegetation groups. The abbreviations of the syntaxonomical units are explained in synopsis and Figure 1.
The total number of taxa recorded in the whole data set is 311, with 38 exceeding 5% of constancy. The most frequently observed species in the study area were Poa relaxa (134), Artemisia rutifolia (97), Pyrethrum pyrethroides (65), Hippolytia darvasica (59), Poa litvinoviana (59), Carex koshevnikovii (57), Stipa glareosa (44), Poa bulbosa (42), Paraquilegia anemonoides (37), Campanula incanescens (35), and Silene microphylla (34). From this group of species, only Poa litvinoviana prefers high alpine swards whereas Poa bulbosa frequently contributes to steppe or ruderal vegetation. The other species are confined to the typical chasmophytic habitats of the Pamir-Alai. Due to the close proximity and habitat similarity, the rock communities of Pamir-Alai frequently consist of scree species, including, e.g.: Bunium badachschanicum, Callipeltis cucullaris, Eremopoa persica, Scariola orientalis, Silene brahuica, or Veronica capillipes. These plants comprise ca. 10% of all the taxa in the data set. Additionally, high altitude semidesert plants often co-occur with chasmophytic species with Crepis flexuosa, Festuca tianschanica, and Krascheninnikovia ceratoides as the most frequent. Moreover, particularly in the eastern Pamir in Tajikistan and Alai Valley in Kyrgyzstan, grassland species, typical for both cryophilous steppes (Stipa glareosa) and montane steppes (e.g., Arenaria serpyllifolia, Bromus oxyodon) can be found. Representatives of other habitats, such as xerophytic shrubs (e.g., Cerasus verrucosa), dwarf Artemisia steppes (e.g., Artemisia korshinskyi) or alpine swards, and cushion vegetation (e.g., Acantholimon diapensioides or Nepeta pamiriensis) were rather irrelevant. Hence, as was found for chasmophytic vegetation in western Pamir-Alai (Nowak et al., 2014a, 2015), the substantial compositional structure of the Pamir and western Tian Shan petrophytic phytocoenoses is homogenous and unique for this kind of habitat.
. Synopsis of Syntaxa
Based on this study, we propose the following classification of vegetation of rock crevices of the alpine belt in the eastern Pamir Alai and south-western Tian Shan Mts:
Class: Asplenietea trichomanis (Br.-Bl. in Meier et Br.-Bl. 1934) Oberdorfer 1977
Order: Campanuletalia incanescentis M. Nobis, A. Nowak et A. Nobis 2013
Alliance Eritrichion pamirico-subjacquemontii nom. prov.
Group I – Rock ledges in highly elevated semiarid zone
1. Community of Androsace villosa var. dasyphylla (Cluster 1) – NMDS 1
2. Crepidifolio tenuifolii-Stipetum tianschanicae Nowak et al. hoc loco (Cluster 2) – NMDS 2
Alliance: Asperulo albiflorae-Poion relaxae M. Nobis, A. Nowak et A. Nobis 2013
Group II – Petrophytic vegetation of rock ledges and clefts of highly elevated moderately humid zone of the southern Pamir
3. Community of Potentilla malacotricha and Waldheimia glabra (Cluster 3) – NMDS 3
4. Parryetum schugnanae Nowak et al. hoc loco (Cluster 4) – NMDS 4
5. Corydalideto bucharicae-Hippolytietum schugnanicae Nowak et al. hoc loco (Cluster 5) – NMDS 5
Group III – The chasmophytic vegetation of the alpine and upper montane belts of the Darvaz, Western Pamir and SW Tian Shan subhumid zone
6. Drabetum darvasicae Nowak et al. hoc loco (Cluster 6) – NMDS 6
7. Community of Erigeron vicarius and Ephedra regeliana (Cluster 7)
8. Paraquilegietum anemonoidis Nowak et al. 2014 (Cluster 8) – NMDS 7
9. Psychrogetonetum andryaloidetis Nowak et al. 2015 (Cluster 9) – NMDS 8
10. Community of Campanula albertii (Cluster 10) – NMDS 9
Alliance: Hippolytion darvasicae Nowak et al. all. nova hoc loco
Group IV – The chasmophytic vegetation of rock crevices in the alpine belt of the thermophilous south-western Pamir
11. Silenetum kuhistanicae Nowak et al. 2014 (Cluster 11)
12. Community of Silene schugnanica (Cluster 12)
13. Scutellarietum schugnanicae Nowak et al. 2014 (Cluster 13) – NMDS 10
Group V – The chasmophytic vegetation of rock ledges in the alpine belt of the south-eastern Pamir
. Description of Petrophytic Vegetation of Alpine Belt of the Pamir and South-Western Tian Shan in Middle Asia
Phytocoenoses of this vegetation occupy the rocky outcrops, shelves, ledges, crevices and fissures and occasionally screes in Central and Middle Asian regions with Irano-Turanian climate with strong continental influence.
. The High Alpine Rock Ledges Vegetation in Subarid Zone of the Pamir
Eritrichion pamirico-subjacquemontiinom. prov.
Diagnostic species:Erigeron vicarius, Eritrichium pamiricum, E. subjacquemontii, Lonicera humilis, Roegneria schugnana, Rubia tibetica.
Floristic and habitat characteristics: This type of high alpine vegetation is characteristic of the harsh conditions in subarid zone of the Eastern Pamir, but can also be found on the highest, windswept peaks in West Pamir. Communities of Eritrichion pamirico-subjacquemontii inhabit the clefts and shadowed ledges of rocks in Rushan, Yazgulem, Ishkashim, Vakhan, Alichur, Muzkol, and Schugnan ranges. It is mainly noted in high alpine elevations between (3,200–)3,500–4,700(–5,500) m a.s.l. It is species poor vegetation of very short blooming period of plants in July–August and extremely continental climate with yearly temperature amplitude between −40 in winter and +50 in summer.
Group I – Rock ledges in highly elevated semiarid zone
1. Community of Androsace villosa var. dasyphylla (Cluster 1)
Diagnostic species:Androsace villosa var. dasyphylla, Asperula oppositifolia, Helianthemum songaricum, Pentanema albertoregelia, Stipa orientalis.
Constant species:Androsace lehmanniana, Stipa orientalis.
Dominant species:Androsace lehmanniana, Arabis kamelinii, Artemisia rutifolia, Pentanema albertoregelia, Stipa orientalis.
Floristic and habitat characteristics:Androsace villosa var. dasyphylla (= Androsace dasyphylla) is scattered across the alpine chasmophytic habitst in Pamir-Alai and Tian Shan ranges. In Tajikistan, it is mostly distributed in high altitudes of Eastern Pamir, and occasionally also in Hissar and Kuraminian Mts (Chukavina, 1984). The plant prefers rock ledges and clefts; however, it also grows on solid rock faces or on fine-gravel screes. During the study the community was found in the Pamirian alpine belt (Figure S1) between 2,500 and 3,500 m a.s.l. (mean approx. 2,650; Figure 4A), mainly on granite or limestone rock ledges and clefts with accumulated soil. The community inhabits mainly southern and south-western expositions with relatively gentle inclinations (40°–85°, mean approx. 55°; Figure 4D). As in similar high altitude petrophytic vegetation it characterizes by scarce vascular plant cover ranging between 10% and 20% (close to 19% on average; Figure 4E) and is species poor (between four and seven species per plot, and six taxa on average; Figure 4G). The moss layer is not abundant reaching up to 5% per plot (Figure 4F). It includes fairly homogenic patches in terms of species composition that reflects the Central Asian climatic influences by the contribution of, e.g., Stipa orientalis or Festuca tianschanica. Because the community is closely related to high altitude wind-swept swards and the main diagnostic species is widely distributed in Central Asia, we leave the community rankles at this stage of the research.
2. Crepidifolio tenuifolii-Stipetum tianschanicae Nowak et al. hoc loco (Cluster 2; Figure 5A)
Type relevé:Table S1, ordinal plot number 13, holotypus hoc loco.
Diagnostic species:Ajania gracilis, Allium tianschanicum, Angelica ternata, Crepidifolium tenuifolium, Crepis flexuosa, Dracocephalum stamineum, Krasheninnikovia ceratoides, Roegneria schrenkiana, Stipa tianschanica.
Constant species: -
Dominant species:Ajania gracilis, Allium tianschanicum, Angelica ternata, Crepidifolium tenuifolium, Dracocephalum stamineum, Eritrichium subjaquemontii, Stipa tianschanica.
Floristic and habitat characteristics: Both name giving plants (Crepidifolium tenuifolium and Stipa tianschanica) are widely distributed in Central Asia in semiarid chasmophytic habitats (Nobis, 2014; Nobis et al., 2020; Rasulova, 1991). Crepidifolium tenuifolium (= Youngia diversifolia) additionally spread across the southern Siberian ranges. In Tajikistan, the main distribution range of this community includes Eastern Pamir with some scattered locations also in the Western Pamir (Figure S1). It grows mainly on large ledges and crumbling rocks in dry, deserted slopes of Alichur, Schugnan and Sari-kol Mts (Rasulova, 1991). Probably, it can thrive also on ephemeral alpine brook bars and steep screes. The patches of the association have been recorded fairly often within the range of altitudes from 2,600 to 4,000 m a.s.l. (average 3,650; Figure 4A), mainly on limestone substrates. This association inhabits relatively steeply sloping rocks (35°–85°) with the mean inclination of approx. 70° (Figure 4D). It was found mainly on eastern and northern expositions. The association is characterized by a sparse herb cover with the mean close to 30% and is almost devoid of moss contribution (mean approx. 1%; Figure 4E). The species richness within documented patches was low, with the average seven species and maximum of 10 plants per plot (Figure 4G). In comparison to other communities, the Crepidifolio tenuifolii-Stipetum tianschanicae inhabits the driest habitats with low nutrient content.
Figure 5
Photographs of the chasmophytic vegetation belonging to the: (A) Crepidifolio tenuifolii-Stipetum tianschanicae in the North Alichurian Range in the Eastern Pamir, (B) community of Potentilla malacotricha and Waldheimia glabra in the rock outcrops to the north of Jelondy, (C) Parryetum schugnanae on the limestone rocks near Jelondy, (D) Corydalideto bucharicae-Hippolytietum schugnanicae on the solid limestone rock faces in the Khuf River Valley in the Western Pamir, (E) Drabetum darvasicae on the limestone rock ledges close to Khoburobod Pass in Darvaz Range, (F) Psychrogetonetum andryaloidetis in the limestone rock clefts in the Obikhingou River valley close to Tavildara. All pictures were taken by A. Nowak.

. The Petrophytic and Chasmophytic Vegetation of the Alpine and Subnival Zone
Group II – Petrophytic vegetation of rock ledges and clefts of highly elevated moderately humid zone of the Pamir
3. Community of Potentilla malacotricha and Waldheimia glabra (Cluster 3; Figure 5B)
Diagnostic species:Acantholimon lycopodioides, Draba altaica, Festuca alaica, Potentilla malacotricha, Rhodiola coccinea, Roegneria schugnana, Waldheimia glabra.
Constant species: -
Dominant species:Acantholimon diapensioides, Acantholimon lycopodioides, Paraquilegia anemonoides, Potentilla malacotricha, Rhodiola coccinea, Stipa caucasica glareosa, Waldheimia glabra.
Floristic and habitat characteristics: This vegetation of rock ledges was noted in the highest, subnival elevations of subhumid zone, exclusively in Eastern Pamir (Figure S1). Due to wide range of the diagnostic taxa (at least Waldheimia glabra), restricted range of the sampled plots and some taxonomic ambiguity regarding the Potentilla malacotricha (unaccepted name according to The Plant List) we left this community rankless. Additional sampling will be indispensable along with taxonomic research on Potentilla agrimonioides group to finally defined this vegetation. Despite these uncertainties and the close relation to alpine swards (e.g., Nepeta pamiriensis communities) it is fairly distinct community in the subnival belt. The communities prefer eroded ledges, and large clefts with a fairly high content of organic matter and water availability. Plots noticed in the Pamir were found between 3,850 to 4,150 m a.s.l. (mean approx. 4,000; Figure 4A). Plots of this community inhabit sloping walls and screes with the mean inclination value approx. 65°, ranging from 45° to 85° (Figure 4D). The phytocoenoses occur generally on eastern exposition (Figure S2). Its total cover of herb layer is moderate. It ranges between 20% and 30% (mean ca. 28%; Figure 4E). It is species poor community with five to 11 species per plot (mean ca. seven; Figure 4G). The moss layer is scarce and has the average value close to 2% (Figure 4F). Because of the close vicinity of the alpine swards, the species typical for wind swept vegetation contribute to this chasmophytic community with Acantholimon diapensioides as the most abundant.
4. Parryetum schugnanae Nowak et al. hoc loco (Cluster 4; Figure 5C)
Type relevé:Table S1, ordinal plot number 55, holotypus hoc loco.
Diagnostic species:Parrya schugnana, Potentilla moorcroftii, Pyrethrum pyrethroides, Semenovia heterodonta.
Constant species: Parrya schugnana.
Dominant species:Acantholimon diapensioides, Arabis kamelinii, Artemisia rutifolia, Hippolytia schugnanica, Macrotomia euchroma, Parrya schugnana, Potentilla moorcroftii, Pyrethrum pyrethroides, Semenovia heterodonta.
Floristic and habitat characteristics:Parrya schugnana is an endemic species with the core area of distribution in the alpine belt of western Pamirian ranges (particularly Schugnan, Vanch, and Rushan Mts; Ovchinnikov, 1978). It inhabits large lefts and ledges on limestone and schist outcrops. It was noted in cracks and ledges at the elevation between 3,100 and 3,850 m a.s.l. (mean 3,400; Figure 4A). The plots were found on southern or south-western exposition and were located on steep rock wall (50°–80°, mean ca. 70°; Figure 4D). However, plots under the overhanging rock balconies were also observed. The total cover of vascular plants was between 20% and 40%, mean approx. 30% (Figure 4E). The moss layer has negligible contribution to this vegetation (Figure 4F). The association is moderately rich in species and composed of four to 11 with the mean of eight (Figure 4G). Parryetum schugnanae is typical rock wall chasmophytic community in eastern Pamir-Alai.
5. Corydalideto bucharicae-Hippolytietum schugnanicae Nowak et al. hoc loco (Cluster 5; Figure 5D)
Type relevé:Table S1, ordinal plot number 74, holotypus hoc loco.
Diagnostic species:Corydalis bucharica, Hippolytia schugnanica, Piptatherum sogdianum, Scrophularia pamirica.
Constant species:Hippolytia schugnanica.
Dominant species:Acantholimon diapensioides, Corydalis bucharica, Erigeron bellidiformis, Festuca alaica, Hippolytia schugnanica, Macrotomia euchroma, Parrya schugnana, Scutellaria shugnanica, Thalictrum isopyroides.
Floristic and habitat characteristics: This association has similar extent of occurrence as the described above. It prefers schist, granite, and mudstone rocks in southern Pamir (Figure S1). Both name giving taxa are considered to be endemic to Tajikistan-Afghanistan borderland within the western Pamir. Corydalis bucharica extends more westward and reaches the montane belt of south-western Pamir-Alai (Mikhailova & Sochivko, 2011). In comparison to other Pamirian chasmophytic plant associations, Corydalideto bucharicae-Hippolytio schugnanicae inhabits warmer, well insolated sites. Due to strong sunlight, in the late summer, the phytocoenosis almost dries up and disappears. It was noted on rock faces with crevices and cracks, sometimes also small ledges in the alpine belt of Schugnan and Rushan ranges. The phytocoenoses develop in alpine belt with mean elevation value of approx. 3,100 m a.s.l. (2,650–3,200 m a.s.l.; Figure 4A). The community inhabits preferably steep rock walls with mean inclination of approx. 80° (Figure 4D), sometimes also under the overhanging rocks. The patches of Corydalideto bucharicae-Hippolytietum schugnanicae were noted mainly on western and south-western expositions. Only 5–13 taxa were noted within the patches and the mean species number per plot was eight (Figure 4G). The vegetation abundance is moderate with the average vascular plant layer cover of about 25% (Figure 4E). Mosses have inconsiderable share and cover up to 5% of the surface (Figure 4F).
Group III – The chasmophytic vegetation of the alpine and upper montane belts of the Darvaz and Western Pamir subhumid zone
6. Drabetum darvasicae Nowak et al. hoc loco (Cluster 6; Figure 5E)
Type relevé:Table S1, ordinal plot number 86, holotypus hoc loco.
Diagnostic species:Cystopteris fragilis, Draba darvasica, Erigeron brachyspermus, Hymenolaena nana, Hyssopus seravschanicus, Inula glauca, Rhodiola heterodonta, Sedum ewersii.
Constant species:Draba darvasica.
Dominant species:Carex koshevnikovii, Draba darvasica, Erigeron brachyspermus, Hymenolaena nana, Hyssopus seravschanicus, Inula glauca, Parrya runcinata, Rhodiola heterodonta, Sedum ewersii.
Floristic and habitat characteristics: The Darvaz and Peter the First Mountains standing between the high Pamir and western Pamir-Alai, get much higher precipitation (Figure 4C) if compared to more arid lands eastwards. This is one of the reasons for the extremely high endemism of these mountains, particularly the Darvaz range. One of the exclusive plants of this zone is Draba darvasica known from few sites, including the area of Khoburobot Pass (Figure S1). It inhabits the limestone rock outcrops in the belt of high alpine pastures. The association prefers crevices in solid faces and small ledges with insignificant soil amount. The phytocoenosis develops in alpine belt with mean elevation of approx. 2,900 m a.s.l. (from 2,850 to 2,950 m a.s.l.; Figure 4A). The association was recorded on steep rock walls with mean inclination of approx. 70° (Figure 4D) and prefers south-western and southern expositions (Figure S2). The mean value of herb layer cover is about 30% ranging from 15% to 75% (Figure 4E). Mosses occur sporadically with the mean cover value of ca. 7% (Figure 4F). Only in single plot dominated by Sedum ewersii its share reaches 45%. The vegetation is moderately rich in species and composed of six to 11 species, approx. seven per plot (Figure 4G).
7. Community of Erigeron vicarius and Ephedra regeliana (Cluster 7)
Diagnostic species:Bromus oxyodon, Ephedra regeliana, Eremopyrum bonapartis, E. distans, Erigeron vicarius, Lophantus virescens, Macrotomia cana, Onosma gmelinii.
Constant species:Artemisia rutifolia, Erigeron vicarius.
Dominant species:Artemisia rutifolia, Erigeron vicarius.
Floristic and habitat characteristics: This high alpine chasmophytic community is insufficiently documented and has to be surveyed in wider range. We found only two plots in limestone rock ledges in the Trans-Alai Mts. Due to the small sample number and the limited range of sampling this community must be leaved rankless. Considering the ranges of both main diagnostic taxa, it can be expected that this type of vegetation occurs in the mountains of Central Asia, including Tian Shan in eastern China (Xinjiang Province). The collected data is insufficient and should be supplemented in future.
8. Paraquilegietum anemonoidis Nowak et al. 2014 (Cluster 8)
Diagnostic species:Draba lanceolata, Gagea olgae, Paraquilegia anemonoides, Taphrospermum altaicum, Viola allochroa.
Constant species:Paraquilegia anemonoides.
Dominant species:Paraquilegia anemonoides, Rhamnus minuta, Viola allochroa.
Floristic and habitat characteristics: This typically chasmophytic association of solid rock faces was firstly described from the western Pamir-Alai (Nowak, Nowak, et al., 2014a). As the diagnostic species has wide geographical range, it was expected that the range of the community will increase when the survey will be continued. During the investigation of the alpine belt of Eastern and Western Tian Shan we found several patches representing Paraquilegietum anemonoidis on limestone walls in Alai Mts in Kyrgyzstan (Figure S1). The patches have been found at altitudes from 2,400 to 3,000 m a.s.l. (Figure 4A) on various exposition (Figure S2). As in western Pamir-Alai the community inhabits fairly solid rock faces with almost vertical inclinations (mean approx. 80°; Figure 4D). It prefers coherent rocks with small-sized crevices with inconsiderable amount of organic matter. The association is characterized by a moderate herb cover value of approx. 25% (between 15% and 30%; Figure 4E). Sampled plots have medium number of species, between three and 11 (mean value approx. six per relevé; Figure 4G). The moss layer is negligible. It does not exceed 5% (Figure 4F).
9. Psychrogetonetum andryaloidetis Nowak et al. hoc loco (Cluster 9; Figure 5F)
Type relevé:Table S1, ordinal plot number 96, holotypus hoc loco.
Diagnostic species:Psychrogeton andryaloides.
Constant species:Psychrogeton andryaloides.
Dominant species:Acantholimon diapensioides, Artemisia rutifolia, Asperula oppositifolia, Carex koshevnikovii, Inula schmalhausenii, Parrya schugnana, Psychrogeton andryaloides, Silene microphylla.
Floristic and habitat characteristics: This association is one of the most spread across the eastern Pamir-Alai, particularly in Western Pamir and Peter the First Range (Figure S1). Because the main diagnostic species, Psychrogeton andryaloides, is broadly distributed plant in alpine and subnival belts of Hindukush (Kinzikaeva, 1988), Kunlun and the Himalayas, one can expect the occurrence of this association also in these ranges. The patches of Psychrogetonetum andryaloidetiswere found in a number of sites, mainly in the Eastern Tajikistanian geobotanical region on limestone, marl and schists. This association prefers fissured, cracked, solid rocks. It inhabits large crevices and clefts with inconsiderable amount of soil substrate. Its phytocoenoses develop on moderate elevations in upper montane and alpine belts, between 2,000 and 3,000 m a.s.l. (mean approx. 2,750; Figure 4A). The community develops on walls of various steepness, from 15° to 90°. The mean inclination value for the sampled plots was approx. 70° (Figure 4D). The association prefers well exposed to the sunlight south-western slopes (Figure S2). The cover of herb layer is moderate. It ranges between 10% and 35% (mean approx. 25%; Figure 4E). The moss layer, particularly in lower elevations is fairly well developed and reaches up to 25%, however, the mean is close to 4% (Figure 4F). The species richness is rather low with three to 11 species per plot (mean 4, Figure 4G).
10. Community of Campanula albertii (Cluster 10)
Diagnostic species:Allium oreoprasum, Campanula albertii, Dasiphora parviflora, Parrya pinnatifida, Peucedanum transiliense, Rhinactinidia limoniifolia, Silene lithophila, Stipa gracilis.
Constant species: -
Dominant species:Artemisia rutifolia, Asperula laevis, Campanula albertii, Dasiphora parviflora, Erigeron bellidiformis, Parrya pinnatifida, Rhinactinidia limoniifolia, Silene lithophila.
Floristic and habitat characteristics: The plots concentrated in a group number 10 differ from the other samples by a clearly lower altitudinal range (Figure 4A). This is a very heterogeneous group that includes plots from the lowest locations of the alpine belt. They were dominated by several species like Silene lithophila, Campanula albertii, Parrya pinnatifida, or Rhinactinidia limoniifolia that have the main occupation range in the montane belt. They were presented in detail in our separate paper devoted mostly to this vertical zone of the eastern Pamir-Alai and Western Tian Shan (Nowak et al., 2021). To find better consistency with other chasmophytic vegetation and show the relations between different saxatile communities, we decided to keep this cluster in our data set.
. The Chasmophytic Vegetation of the Alpine Belt of the Pamir
Hippolytion darvasicaeNowak et al. all. nova
Nomenclatural type:Inuletum schmalhausenii Nowak et al. hoc loco.
Diagnostic species:Arabis karategina, Hippolytia darvasica, Inula schmalhausenii, I. schischkinii, Ribes heterotrichum, Silenemicrophylla, S. schugnanica, Piptatherum vicarium.
Floristic and habitat characteristics: This type of chasmophytic vegetation is a vicariant one to Western Pamir-Alai phytocoenoses of Asperulo albiflorae-Poion relaxae. It is distributed mainly in the southern, open to south valleys of the Pamir, mainly in the southern sections of the Vanch, Rushan, Yazgulem, Ishkashim, Vakhan, and Schugnan ranges. However, it also occurs in the warmest sites in the south Darvaz slopes and possibly the Peter the First Mts. Communities of Hippolytion darvasicae inhabit mainly alpine elevations between (1,400–)1,700 and 2,800(–3,700) m a.s.l. In comparison to the vicariant communities from western Pamiro-Alaian they have slightly higher average species number (ca. eight) per plot and occur in lower locations, 2,200 m a.s.l. on average.
Group IV – The chasmophytic vegetation of the rock crevices in the alpine belt of the thermophilous south-western Pamir
11. Silenetum kuhistanicae Nowak et al. 2014 (Cluster 11)
Diagnostic species:Alyssum campestre, Eremurus comosus, Melica jacquemontii, Polygonum polycnemoides, Silene kuhistanica, Thalictrum isopyroides.
Constant species:Silene kuhistanica.
Dominant species:Silene kuhistanica.
Floristic and habitat characteristics: This chasmophytic community was reported firstly from the core occurrence area in western Pamir-Alai (Nowak, Nowak, et al., 2014c). In eastern Pamir-Alai, Silene kuhistanica occurs sporadically scattered in Darvaz and Peter the First ranges (Figure S1). We found only one plot representing this association. It was located on the distributional limits of the main diagnostic species. It inhabits the limestone steep wall (80° inclination; Figure 4D) at ca. 2,100 m a.s.l. (Figure 4A). The exposition of the rock was south-western (Figure S2) and the vegetation cover 25% (Figure 4E). Within the plot, nine species were noted (Figure 4G).
12. Community of Silene schugnanica (Cluster 12)
Diagnostic species:Campanula incanescens, Cheilanthes persica, Corydalis gortschakovii, Kudryaschewia allotricha, Parietaria judaica, Silene schugnanica.
Constant species:Silene schugnanica.
Dominant species:Silene schugnanica.
Floristic and habitat characteristics:Silene schugnianica occupies southern slopes of Darvaz Mts, Peter the First and Western Pamir ranges. It is endemic to Tajikistan, inhabiting the base parts of the limestone walls, screes, and sometimes also ruderal sites along roads. We found only four plots from one site with this species, fairly distinct from other vegetation and thus separated by the Twinspan algorithm. It was found mostly on the southern, steep (80° inclination; Figure 4D) slope of Darvaz Mts (Figure S1). The plot consists of nine species on average (Figure 4G) and total herbaceous cover was up to 30% (Figure 4E).
13. Scutellarietum schugnanicae Nowak et al. 2014 (Cluster 13; Figure 6A)
Diagnostic species:Carex koshevnikovii, Cephalopodum badachschanicum, Potentilla evestita, Scutellaria shugnanica.
Constant species:Carex koshevnikovii, Scutellaria shugnanica.
Dominant species:Asperula laevis, Carex koshevnikovii, Cephalopodum badachschanicum, Hippolytia darvasica, Potentilla evestita, Scutellaria shugnanica.
Floristic and habitat characteristics: This association is one of the most spread along the Panj River Valley (Figure S1), particularly frequent in Western Pamirian ranges (Schugnan, Vanch and Rushan Mts; Kochkareva, 1986). The main diagnostic species is endemic with a narrow range restricted to southern Tajikistan. The altitudinal amplitude of the association is fairly wide extending from colline elevations (ca. 1,300 m a.s.l.) to high alpine belt (ca. 3,600 m a.s.l.; Figure 4A). As in montane belt, this association prefers crevices and clefts on different rock substrates, mainly granite, schist, dolomite, or conglomerate rocks (see Nowak, Nowak, et al. 2014b). It was found at the lowest alpine and montane elevations with the mean of approx. 1,950 m. The community develops on steep rock walls with mean inclination of approx. 85° (Figure 4D) on southern or south-western expositions (Figure S2). The total cover of herb layer is about 25% on average, and it ranges from 10% to 40% (Figure 4E). Mosses enter the plots quite rarely and have the average cover value of ca. 4% (Figure 4F). The montane character of this association is reflected by the prominent contribution of Carex koshevnikovii, Dianthus darvazicus, or Scrophularia scabiosiifolia.
Figure 6
Photographs of the chasmophytic vegetation belonging to the: (A) Scutellarietum schugnanicae on the limestone rocks in the Panj River Valley neat Qualaikhum, (B) Acantholimonetum alexeenkoani on the limestone rock clefts near Ishkashim in the Southern Pamir, (C) community of Rosularia alpestris on the limestone ledges in the Alai Range near Osh, (D) Inuletum schmalhausenii on the limestone rock clefts in the Ghunt River Valley close to Khorogh. All pictures were taken by A. Nowak.

Group V – The chasmophytic vegetation of the rock ledges in the alpine belt of the south-eastern Pamir
14. Acantholimonetum alexeenkoani Nowak et al. hoc loco (Cluster 14; Figure 6B)
Type relevé:Table S1, ordinal plot number 192, holotypus hoc loco.
Diagnostic species:Acantholimon alexeenkoanum, Astragalus longistipitatus, Hippolytia darvasica, Minuartia litvinovii, Piptatherum alpestre, Rosa huntica, Rosularia glabra, Tanacetopsis mucronate.
Constant species:Acantholimon alexeenkoanum, Hippolytia darvasica.
Dominant species:Acantholimon alexeenkoanum, Carex koshevnikovii, Cephalopodum badachschanicum, Hippolytia darvasica, Minuartia litvinovii, Tanacetopsis mucronate.
Floristic and habitat characteristics: This association is found within the range of name giving species Acantholimon alexeenkoanum which is endemic to Pamir (Figure S1). It is distributed in fairly dry valleys of Shakhdara, Wakhan, and Ishkashim ranges. It was found at the elevations from ca. 2,000 to 2,750 m a.s.l. (mean ca. 2,350). Its patches develop on limestone clefts and ledges and prefer sloping walls with different inclination from 55% to 90% (Figure 4D), sporadically also under overhanging galleries. The recorded phytocoenoses were usually exposed to the west (Figure S2). The community patches were fairly rich in species with six to 18 taxa per plot (mean 11; Figure 4G) and considerable plant cover ranging from 15% to 40% (mean ca. 25%; Figure 4E).
15. Community of Rosularia alpestris (Cluster 15; Figure 6C)
Diagnostic species:Clypeola jonthlaspi, Ferula kokanica, Rosularia alpestris, Silene brahuica, Stellaria alsinoides, Ziziphora bungeana.
Constant species:Rosularia alpestris.
Dominant species:Bromus tectorum, Rosularia alpestris.
Floristic and habitat characteristics:Rosularia alpestris is a rare plant species in Middle Asia, scattered from Western Pamir-Alai to Eastern Tian Shan. The plots with this species were found in few locations in western section of the Alai Range (Figure S1). They probably belong to the boulder and rock ledges vegetation that still needs to be surveyed and analyzed in Middle Asia. Physiognomically and ecologically it is supposed to be similar to palaearctic group of Alysso-Sedion with different Rosularia spp., Sedum spp., and additionally Clypeola jothlaspi and Alyssum spp. At the present stage of the research, we leave the community rankless as we have scarce data from restricted area. We found plots of the community on limestone rock ledges at the elevation of about 2,100 m a.s.l. (Figure 4A). It inhabits gently sloping rock plates (10°–20° with the average of ca. 15°; Figure 4D) with prevailing northern expositions (Figure S2). The total cover of herb layer is fairly abundant and ranges between 20% and 30% (Figure 4E). The presence of moss layer was noted with the mean cover value of ca. 7% (Figure 4F).
16. Inuletum schmalhausenii Nowak et al. hoc loco (Cluster 16; Figure 6D)
Type relevé:Table S1, ordinal plot number 221, holotypus hoc loco.
Diagnostic species:Hippolytia darvasica, Inula schischkinii, Inula schmalhausenii, Silene microphylla.
Constant species:Hippolytia darvasica, Silene microphylla.
Dominant species:Hippolytia darvasica, Inula schischkinii, Inula schmalhausenii, Poa relaxa, Silene microphylla.
Floristic and habitat characteristics:Inula schmalhausenii (along with I. schischkinii) is typically West Pamirian plant that spread in Rushan, Yazgulem, Gunt, Shakhdara, and Bartang valleys. This very distinct association inhabits different rock substrates but prefers granite ledges and clefts. Only plots with Silene microphylla (= S. guntensis) have preferences to solid rock faces (Figure S1). This vegetation inhabits alpine belt at the elevation of ca. 2,000–2,900 m a.s.l. (mean ca. 2,350; Figure 4A). The phytocoenoses develop on slopes with very different inclinations (ranging from 45° to 110°) with the mean of approx. 80° (Figure 4D). Inuletum schmalhausenii inhabits slopes with various expositions with some preference to north-western and western (Figure S2). The mean total cover of herb for the association is close to 30%, ranging from 10% to 40% (Figure 4E). The association is moderately rich in species, having on average eight taxa per plot (from three to 11; Figure 4G).
. Discussion
The mountains of Central Asia have been identified as one of the world’s biodiversity hotspots (Brooks et al., 2006; Critical Ecosystem Partnership Fund, 2017; Mittermeier et al., 2011), and as one of the 11 most important focal points of scientific studies and conservation efforts (Giam et al., 2010). In every phytogeographical approach (e.g., Goncharov, 1937; Kamelin, 2010; Takhtajan, 1986), the region of Middle Asia and in particular Tajikistan is entangled by boundaries of many subunits. In particular, the eastern part of Tajikistan and south-western Tian Shan is separated from the western part by a very important border of subregions: Central Asian subregion vs. Western Asian or Western Himalayan Province vs. Central Tian Shan Province (Takhtajan, 1986) or Ancient Mediterranean vs. Eastern Asian (Oriental) subregions or Montano-Middle Asian (Afghano-Turanian) vs. Pamiro-Tibetan vs. Western Himalayan (Kamelin, 2010, 2017). It is worth mentioning that despite the harsh conditions in alpine belt of the Middle Asian mountains, approx. 700 typical oreophilous taxa were listed for the region (Kamelin, 2010). All of these circumstances, combined with the unusual richness of a rock species pool, produce a stunningly diverse vegetation in petrophytic habitats in Middle Asia. Additionally, the global analysis of the species diversity of alpine ecosystems detects the highest scores in Middle Asia. The occurrence of such high diversity in the region located in temperate latitudes is explained by the historical abundance of alpine ecosystems, with a predominant effect of regional idiosyncrasies on community species richness (Testolin et al., 2020). Other mountainous regions such as the Mediterranean Region (Deil & Hammoumi, 1997; Dimopoulos et al., 1997; Galán de Mera et al., 2000; Parolly, 1998) or western or South Africa (Nagy & Grabherr, 2009) are not as diverse due to the lower ecosystem heterogeneity, different geological history, and lower isolation.
The strong phytogeographic distinction and clear geobotanical regionalization are not fully expressed by the pattern of rock vegetation in the eastern Pamir. It is caused by the fact that some species previously considered diagnostic for the established communities in Western Pamir-Alai seem to have a wider range towards east and north. An example is Poa relaxa, which is found on solid rocks from 1,200 to 4,000 m a.s.l. almost in the whole Middle Asia. This species is considered to be diagnostic for Asperulo-Poion relaxae which is typical for solid rock faces of Western Pamir-Alai (Nobis et al., 2013; Nowak, Nowak, et al., 2014a). In a case of species-poor plots, a single taxon can cause some vagueness in syntaxonomical classification. However, if one looks at the whole set of diagnostic species for Poion relaxae (i.e., Artemisia rutifolia, Asperula albiflora, A. czukavinae, Lophanthus virescens), the explanation why the western Pamiro-Alai vegetation is distinguished from the Pamirian one by the comparative algorithm seems more obvious. Neither Asperula albiflora or A. czukavinae (only A. fedtschenkoi, A. leavis, and A. oppositifolia are present), nor Lophanthus virescens or Stipa zeravshanica contribute to the Eastern Pamirian vegetation. Only Poa relaxa and Artemisia rutifolia occur in this area, however, with clearly lower constancies. Additionally, it is worth noting that in the Pamirs, the species that form their own communities in western Pamir-Alai, e.g., Sergia regelii, Campanula lehmanniana, Scutellaria megalodonta, S. orbicularis, S. rubromaculata, and many others, are absent (Nowak, Nowak, et al., 2014a). Therefore, the inclusion of Clusters 3, 4, 5, 6, 7, and 8 from the NMDS diagram into the Asperulo-Poion alliance ought to be considered as temporary. Further research in the Eastern Tajikistan and Western Pamir should be continued in order to fix the final arrangement of these communities. Currently, their position is based on the presence of two fairly common taxa in Middle Asia, i.e., Poa relaxa and Artemisia rutifolia.
Another problem with the internal relationships between the chasmophytic communities of the alpine belt is that some of the associations that are almost exclusively restricted to East Pamir (e.g., Potentilla malacotricha-Waldheimia glabra) were clustered together with typically West-Pamirian associations (e.g., Parryetum schugnanae or Corydalideto-Hippolytietum). This is due scarce sampling and the fact that plots representing Potentilla malacotricha-Waldheimia glabra community have been found in close proximity to the Ghunt Valley. Here, the influences of the warmer air masses from the south are significant and therefore the species compositions reflect more thermophilous characteristics. Other examples where nonphytogeographic factors could play an important role are the communities of the Western Pamirs. These were quite heavily scattered by the Twinspan algorithm, and their ranges overlap to some extent. An example is the group consisting of the two associations Corydalideto-Hippolytietum and Parryetum schugnanae, which are apparently separated from Acantholimonetum alexeenkoanae, Inuletum schmalhausenii, and Scutellarietum schugnanicae associations even though they occur in the same area. The latter group has somewhat lower elevational amplitude, but this pattern is puzzling and vague. It depicts how the highly complex relief of the Pamir Mountains, and the neighborhood of the warmer Hindu-Kush climate, the Kashgar desert zone, the Western Irano-Turanian influences, and the wetter Tian Shan alpine climate intersect here to create extremely complex conditions influencing the pattern of petrophytic vegetation.
Due to the extremely high endemism of the petrophytic flora of the eastern Pamiro-Alai and SW Tian Shan (Nowak et al., 2011, 2020), it is difficult to find strong compositional relationships with the rock vegetation of the neighboring areas. This is all the more difficult as there is very little research in the Karakorum, Hindu-Kush, and Western Himalayas areas in the chasmophytic habitats. Only some reports from Altai and southern Mongolia could serve as a reference, at least to the Tian Shan communities (Hilbig, 2000; Zibzeev & Igai, 2019). For the rocky vegetation of Altai-Sayan system, the class Rhodioletea quadrifidae Hilbig 2000 was coined (Hilbig, 2000). However, the species composition differs significantly. Only a few species are shared between Middle Asia and the borderland between southern Siberia and Central Asia. These include, e.g., Potentilla biflora and Sibbaldia tetrandra. In Central Tian Shan, these taxa occur separately on granite massifs at the highest elevations in the subnival belt. Hence, it is hardly comparable to the association of Potentillo biflorae-Sibbaldietum tetrandrae Zibzeev et Igai 2019 (Zibzeev & Igai, 2019). In Altai, this vegetation inhabits rather fine gravel scree habitats at much lower elevation in the alpine belt. Thus, this type of Cryopetrophyton oreoasiaticum sensu Kamelin (2005) has considerably different ecological characteristics and probably occurs in Middle Asia in the highest, subnival belt in more humid zones on northern expositions in Central and Western Tian Shan. With the exception of the two abovementioned species, no other taxa are shared in our data sets (e.g., Cerastium pusillum, Lagotis integrifolia, Melandrium apetalum, Rhodiola quadrifida, Saxifruga cernua, Smelowskia alba, or S. calycina).
A considerable number of alpine chasmophytic communities of Middle Asia are narrowly distributed and consist of a large proportion of endemic species to particular geobotanic subregions of Middle Asia. The community of Inuletum schmalhausenii is restricted to few valleys in the Western Pamir, the Parryetum schugnanae and Scutellarietum schugnanicae localities around Khoburobot Pass in Darvaz Range, the Psychrogetonetum andryaloidetis has apparently the highest frequency in Peter the First and Darvaz Mountains (mainly Obikhingou River Valley), and Acantholimonetum alexeenkoani prefers warmer and semiarid zones in southern Pamir. When analyzing the causes of the high diversity of the vegetation of warmer and lower belts in the Pamir in Middle and Central Asia, it is important to understand the remarkable endemism of the flora of this territory, which reaches approx. 35% (Nowak et al., 2011). There are crucial causes for this phenomenon remaining as an “island” character of a separated massifs serving as evolutionary radiation centers (Deil, 1999; Whittaker & Fernandez-Palacios, 2007). Additionally, when the changing climate blocked the westerlies and enhanced aridification during the Oligocene–Miocene in Middle Asia (Bougeois et al., 2018; Caves et al., 2016), a number of adaptations to rupiculous habitats were developed, such as new strategies for settlement, reproduction, and mitigation of drought stress (Kontopanou & Panitsa, 2020). This process has triggered an effective radiation and the chasmophytic habitats became a cradle of endemism in the high mountains of Middle Asia. Finally, the glacial climate strongly influenced the variability of endemism in Tajikistan (Raduła et al., 2021). Therefore, the species pool of the rocky habitat is particularly rich in Middle Asia and thus we can expect remarkable diversity among the plant communities of the petrophytic habitats. This situation is mirrored to some extent in the opposite site of the Qinghai–Tibet platform in Hengduan Mountains that are also defined as temperate biodiversity hotspots. In this area, rapid in situ speciation of resident lineages about 8 million years ago due to alpine orogeny were detected (Xing & Ree, 2017).
Towards the east and north, the rate of endemism decreases, as, particularly at higher elevations, environmental conditions are harsher and allow far fewer species to thrive (e.g., Kamelin, 2005). Additionally, in Central Asia, in Tian Shan ranges the precipitation profile is clearly temperate and the overall sum of rainfall is higher than in Pamir-Alai. This promotes the larger extent of species and vegetation types that are distributed across vast areas from the Eastern Pamir to southern Siberia (e.g., communities with Potentilla biflora, Saxifraga sibirica, Leontopodium fedtschenkoanum, or Dasiphora parvifolia). It is also worth mentioning that these taxa occupy not only typical rock faces but also fine gravelly screes and large rock ledges in severely eroded walls. This situation is apparent in the subnival belt of the Eastern Pamir in Tajikistan, where at the elevation of approx. 5,000 m a.s.l., it is hardly possible to distinguish the rock and scree vegetation from each other. It comprises Nepeta longibracteata, Erigeron pamiricus, Didymophysa fedtschenkoana, and other chasmophytes. These highly situated and, at the same time, most difficult to access rock plant communities still require more detailed studies. Although the vascular plant flora in the subnival belt includes only ca. 200 species, they form a very distinct group, especially when compared to the typical Irano-Turanian flora of Middle Asia in montane and alpine belts. The most cryophilic habitats of the Pamirs have strong phylogeographic relations with the Altai, Sayan, and other mountain ranges of the northern part of Central Asia and Siberia.
. Supplementary Material
The supplementary material for this article is available in Figshare Digital Repository (https://doi.org/10.6084/m9.figshare.16460400.v1):
Table S1. Analytic table of chasmophytic vegetation in the alpine belt of the eastern Pamir-Alai and western Tian Shan Mountains.
Table S2. Synoptic table of chasmophytic vegetation in alpine belt of the eastern Pamir-Alai and western Tian Shan Mountains.
Figure S1. Distribution maps of the chasmophytic vegetation in alpine belt of the eastern Pamir-Alai and western Tian Shan Mountains.
Figure S2. The exposition preferences of the researched plant communities.
