. Introduction
There are more than 200 species of the genus Valeriana among worldwide flora. They are mainly distributed in the Andes of South America, throughout Eurasia, and in the temperate zone of North America (Takhtajan, 2009). There are 10 species in the latest nomenclatural checklist for the flora of Ukraine (Mosyakin & Fedoronchuk, 1999). Species of the genus Valeriana L. are known for their medicinal properties; biologically active substances have been found in seven species among the flora of Ukraine (Minarchenko, 2005). To discover reliable characteristics for the identification of such species, it is important to increase the amount of information on the micromorphology of their fruits.
Morphological characteristics of fruits are known to be species-specific for many members of the subfamily Valerianoideae and the order Dipsacales (Al-dabbagh & Saeed, 2020; Tsarenko et al., 2019; Tsarenko, Bulakh, et al., 2020; Tsarenko, Tsymbaliuk, et al., 2020; Tsymbaliuk et al., 2018; Zaĭtseva, 2006). In numerous scientific papers as well as various floristic reports, manuals, etc., the morphological features of the fruits of Valeriana spp. are minimized and only the general characteristics of their shape and morphometric data are provided (Gorbunov, 2002, 2014; Katina, 1987; Vakulenko et al., 2016). Moreover, the micromorphological structure of the fruit surfaces have not been well studied. It is important to study these features to understand the issues of phylogeny and taxonomy of certain species as well as the family or the order of Dipsacales as a whole. The aim of this work was to conduct a detailed micromorphological characterization of the surfaces of fruits of Valeriana spp., which are important in pharmacology, in order to identify additional features that are relevant for species recognition.
. Material and Methods
All fruits were obtained from herbarium specimens of the National Herbarium of M. G. Kholodny Institute of Botany, National Academy of Sciences of Ukraine (KW). To obtain data on the general fruit appearance, they were examined using a light binocular stereoscopic microscope (MBS-9; Micromed).
The micromorphological features of the fruits were studied using scanning electron microscopy (SEM) (JSM-6060; JEOL). The samples were sprayed with a layer of gold according to the standard method used. Fruit descriptions were recorded using the terminology generalized in a number of papers (Barthlott, 1981; Plisko, 2000; Ziman et al., 2004).
Photographs of the fruits and the microstructure of their surfaces were taken at ×30 to ×4,000 magnifications. Measurements of the fruit elements were performed using the AxioVision Rel. 4.8 (Carl Zeiss) software. The sample citation herein is based on the original text of the labels (Table S1).
. Results
The fruits of Valeriana spp. are quite similar in appearance. They are somewhat flattened, especially on the adaxial side, mostly elongated-ovoid or ovoid (Figure 1A, Figure 2A, Figure 3A, Figure 4A,B, Figure 5A,B, Figure 6A, Figure 7A,B, Figure 8A, Figure 9A,B, Figure 10A,B). The apex of the fruit is elongated, and the base is rounded and slightly expanded, with a barely visible depression in the middle and a basal fruit raphe. The calyx is at the apex of the fruit, which transforms into a white feathery pappus-like structure, with 8–16 long thick leathery awns and numerous white long soft hair-like expansions (Figure 3A, Figure 4C, Figure 6A). These awns are basically fused together in the form of a membranous crown. In immature fruits, the awns with hairs are wrapped down and unfold only after ripening. The fruits have a border on the edge on both sides that is composed of parenchymal tissue with a noticeable vascular bundle (lateral ribs).
On the adaxial side there is one longitudinal vein in the center, and on the abaxial side there are three (one central and two lateral) longitudinal veins with vascular bundles, which look like ribs from the outside. Some species have a pubescent fruit surface. The hairs are simple, unbranched, unicellular, and thin-walled. Detailed descriptions of the fruits are provided below. The species were categorized according to the accepted system of the genus (Gorbunov, 2014).
Subgenus Valeriana
Section Tuberosae (Hoeck) Grub.
Valeriana tuberosa L. (Figure 1)
The epidermal cells have various shapes – oblong, with convex outer periclinal cell walls, located along the longitudinal fruit axis; on the inter-rib zones, rounded, angular with convex periclinal cell walls covered with a characteristically textured cuticle; and rounded with linear, warty, and striate ornamentation. Some epidermal cells have papillary elongated formations and papillae up to 30–40 µm high, with similar cuticular formations (Figure 1C,D). The anticlinal walls are straight or wavy.
Figure 1
Fruit of Valeriana tuberosa (SEM): General view (A); fragments of the fruit surface (B–D); stomium (E); fragment of the hair surface (F).

The microsculpture of the fruit surface is of the small tuberculate type (Figure 1B,C). The fruits are pubescent on both sides (except for the ribs) with simple ribbon-like slightly curved hairs, rounded at the apex and up to 0.4 mm long. The surface has cuticular formations similar to those on the surface of epidermal cells. On the inter-rib zones there are stomata surrounded by five or six radially located lateral cells, the cuticular sculpture of which is mostly of the striate type, and the stomatal apertures are parallel to the longitudinal axis of the fruits. The cells around the stomata are covered with single unequally divergent wax formations (Figure 1E). The awns are pubescent, with long ribbon-like, often twisted, tortuous hairs, and similar cuticular formations (Figure 1F).
Section Rhizophorae Grub.
Series Montanae Grub.
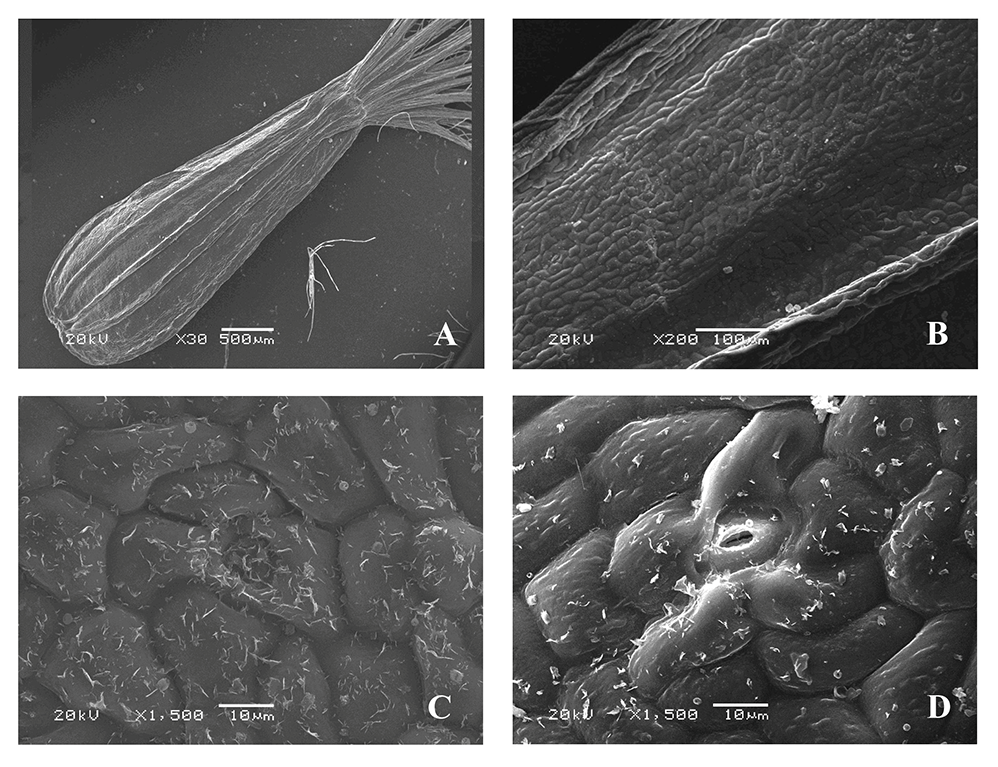
Valeriana tripteris L. (V. transsilvanica) (Figure 2)
The microsculpture of the fruit surface is of the small tuberculate type (Figure 2B–D). The boundaries between the cells are clear. On the ribs, the epidermal cells are located along the longitudinal axis of the fruits in clear rows, with convex outer periclinal walls. On the inter-rib zones, they are rounded, angular, with straight or slightly curved anticlinal and convex periclinal cell walls having rounded and linear warty cuticular ornamentation (Figure 2C,D).
The stomata are located only on the inter-rib zones, surrounded by four or five radially located lateral cells (sometimes the boundaries between the lateral cells are not clear); the shape and sculpture of the surface are similar to those of other epidermal cells. The stomatal apertures are parallel to the longitudinal axis of the fruit (Figure 2C,D). The epidermal cells are covered with epicuticular wax in the form of unequally divergent platelets. There is no pubescence on both sides of the fruits. Awns are pubescent, with long twisted soft ribbon-like hairs and rounded and linear warty cuticular formations.
Figure 2
Fruit of Valeriana tripteris (SEM): General view (A); fragment of the fruit surface (B); stomium (C,D).
Series Dioicae Grub.
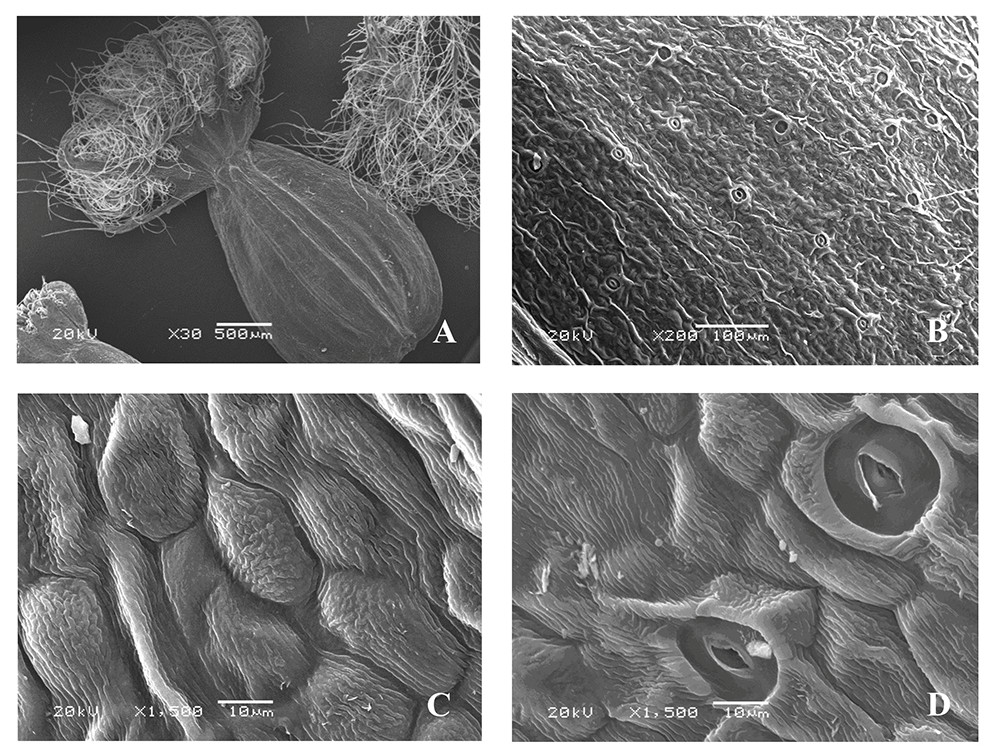
Valeriana simplicifolia (Rchb.) Kabath (Figure 3)
The microsculpture of the fruit surface is of the small tuberculate-crested type (Figure 3B). The boundaries between the cells are clear. On the ribs, the epidermal cells are located along the longitudinal axis, oblong, and smooth, with convex outer periclinal walls and straight, sometimes slightly curved, anticlinal walls. On the inter-rib zones, they are without a clear orientation, elongated, closer to the border, mostly angular, rounded with convex outer periclinal walls, which in some cells are compressed on the sides and form “ridges” slightly raised above the surface of other cells of epidermis, mainly with striate, and some cells with small tuberculate cuticular sculpture. The stomata are located only on the inter-rib zones, surrounded by four or five radially arranged cells. The shape and cuticular sculpture are similar to those of other epidermal cells, and the stomata are mostly parallel to the longitudinal axis of the fruit (Figure 3C,D). The awns are pubescent, with long twisted soft ribbon-like hairs, and surfaces having cuticular formations similar to those on the surface of epidermal cells.
Figure 3
Fruit of Valeriana simplicifolia (SEM): General view (A); fragments of the fruit surface (B,C); stomium (D).
Series Oficinalis Grub.
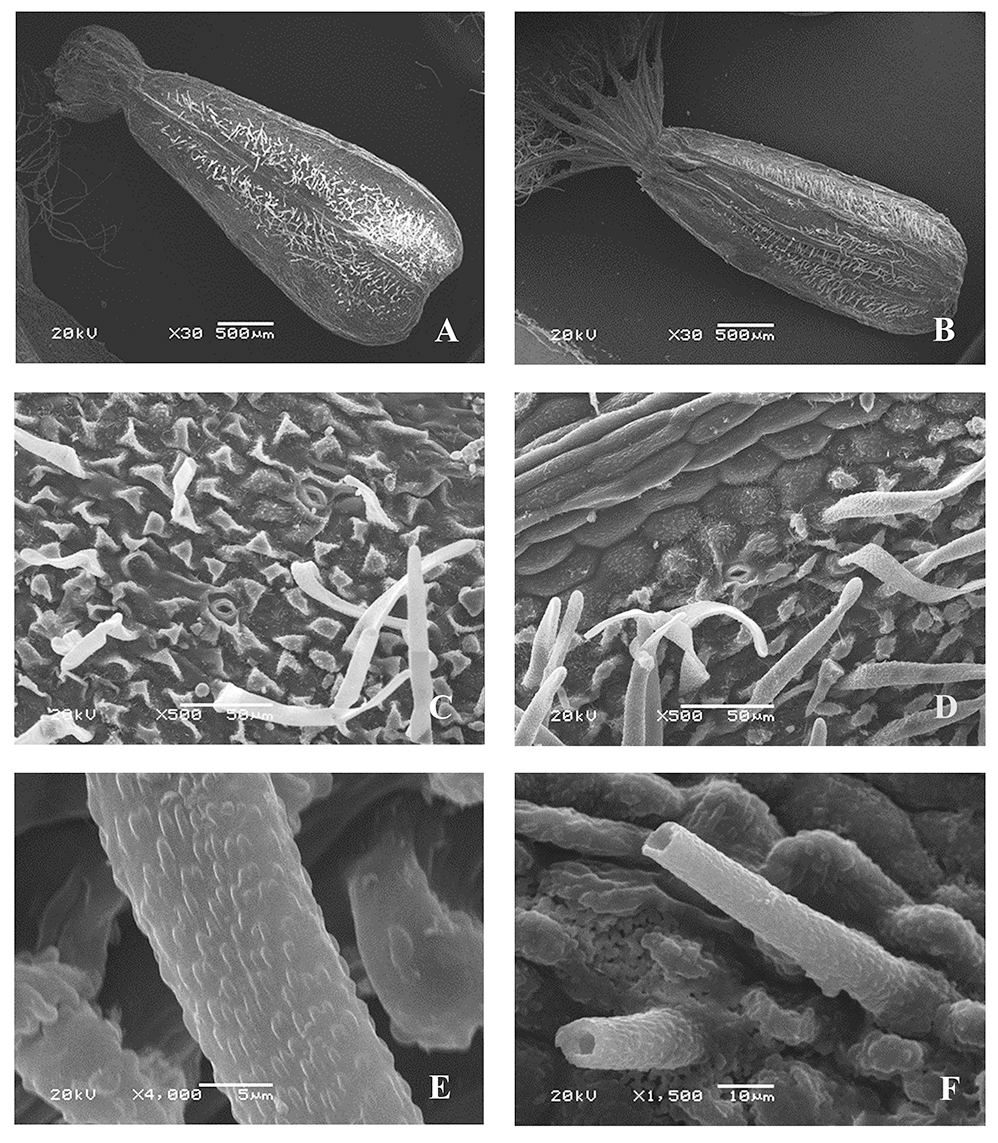
Valeriana wolgensis Kazak. [V. officinalis var. nitida (Kreyer) Rostanski] (Figure 4)
The microsculpture of the fruit surface is of the small tuberculate type (Figure 4D–F). The boundaries between the cells are clear. On the ribs, the epidermal cells are arranged in rows along the longitudinal axis (Figure 4E), and oblong, with convex outer periclinal walls. On the inter-rib zones, they are without a clear orientation or are oriented as on the ribs, rounded, angular, with convex outer periclinal walls and papillary protrusions in the center, and with rounded and linear warty cuticular ornamentation, or in some zones, with striate cuticular formations.
Figure 4
Fruit of Valeriana wolgensis (SEM): General view, adaxial side (A); general view, abaxial side (B); pappus-like calyx (C); fragments of the fruit surface (D–F).

The anticlinal cell walls are straight or slightly curved. Epicuticular wax is present only in some parts of the surface in the form of uneven irregularly divergent platelets. The fruits are pubescent on both sides (except ribs) with simple rigid tubular narrow-conical or ribbon-shaped slightly curved hairs, rounded at the apex, up to 0.2 mm long, and with rounded and linear warty cuticular ornamentation (Figure 4F). The stomata are solitary only in the inter-rib zones, surrounded by four or five lateral cells (sometimes the boundaries between the surrounding cells are indistinct), and the stomatal apertures are parallel to the longitudinal axis of the fruits, the shape and cuticular sculpture of which are similar to those of other epidermal cells (Figure 4E). The awns are pubescent, with long twisted soft ribbon-like hairs, and cuticular sculpture are similar to that of other epidermal cells.
Valeriana officinalis var. nitida (Kreyer) Rostanski (Figure 5)
The micromorphological characteristics of the fruit surfaces of samples defined as V. officinalis var. nitida are identical to those of V. wolgensis, particularly the cuticular ornamentation on the outer periclinal walls (Figure 5C,D) and on the surface of the hairs (Figure 5E,F).
Figure 5
Fruit of Valeriana officinalis var. nitida (Kreyer) Rostanski (SEM): General view, adaxial side (A); general view, abaxial side (B); fragments of the fruit surface (C,D); hairs (E,F).
Valeriana officinalis L. s. str. [J. C. V. exaltata Mikan (V. officinalis var. exaltata J. C. Mikan)] (Figure 6)
The microsculpture of the fruit surface is of the small tuberculate type (Figure 6B,C). The boundaries between the cells are clear. On the ribs, epidermal cells are arranged in rows along the longitudinal axis, oblong, and almost smooth, with convex outer periclinal walls. On the inter-rib zones, they are without a clear orientation, elongated, or rounded, angular, with convex outer periclinal walls, and with rounded and linear warty or striate cuticular formations (Figure 6B,C). In some areas the epidermal cells have papillary protrusions of the periclinal cell walls, the surfaces of which are similar to those of cuticular formations. The anticlinal cell walls are straight or slightly curved. Epicuticular wax is present only in some parts of the surface in the form of unequally divergent platelets. The surface of the fruit is pubescent (on the adaxial side between the ribs, and sometimes on the abaxial side), with simple soft hairs (up to 0.2 mm long), and rounded at the apex.
Figure 6
Fruit of Valeriana officinalis L. (SEM): General view (A); fragments of the fruit surface (B,C); stomium (D).
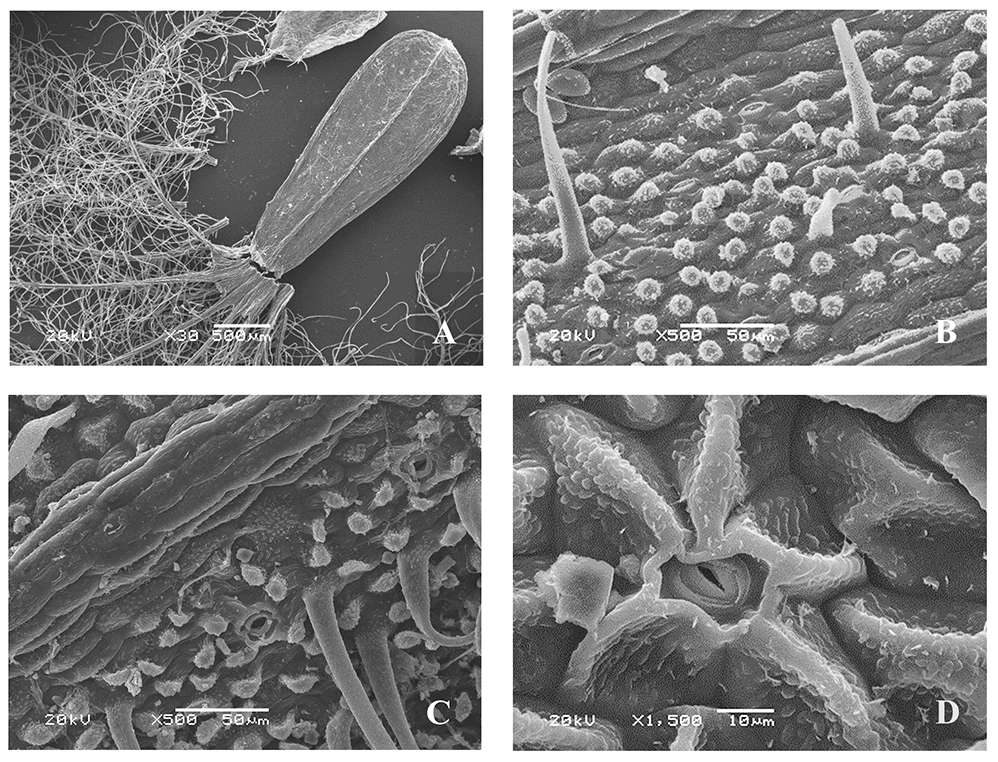
On the surface of the fruit (inter-rib zones), the stomata are oriented along the longitudinal axis, surrounded mainly by five (sometimes only one or two) radially located lateral cells, the shape and cuticular sculpture of which are similar to those of other epidermal cells (Figure 6D). The awns are pubescent, with long twisted soft ribbon-like hairs, and rounded and linear warty cuticular formations.
Valeriana grossheimii Worosch. (Figure 7)
The microsculpture of the fruit surface is of the small tuberculate type (Figure 7C). The boundaries between the cells are clear. On the ribs, the epidermal cells are arranged in rows along the longitudinal axis of the fruit, oblong, and smooth, with convex outer periclinal walls. On the inter-rib zones, they are without clear orientation, elongated, or rounded, angular, with convex or concave outer periclinal and with clear straight, rounded or slightly curved anticlinal cell walls. Some epidermal cells have papillary protrusions of the outer periclinal walls or are replaced by papillae up to 30–40 µm long, with rounded and linear warty and striate cuticular formations (Figure 7C). On the surface of the fruit (inter-rib zones) there are occasional stomata, which are surrounded by five lateral cells that are similar to other cells of the epidermal cuticular formations, and the stomatal apertures are parallel to the longitudinal axis of the fruit (Figure 7D). Epicuticular wax is present only in some parts of the surface in the form of unequally divergent platelets. On the adaxial side, the fruits are pubescent, with hairs rounded at the apex and up to 0.2 mm long. The surface of the hairs is covered with rounded warty cuticular formations. The awns are pubescent, with long twisted soft ribbon-like hairs, and similar cuticular formations.
Figure 7
Fruits of V. grossheimii Worosch. (SEM): General view, adaxial side (A); general view, abaxial side (B); fragment of the fruit surface (C); stomium (D).

Valeriana sambucifolia J. C. Mikan (Figure 8)
The microsculpture of the fruit surface is of the small tuberculate type (Figure 8B,C). The boundaries between the cells are clear. On the ribs, the epidermal cells are arranged in rows along the longitudinal axis, oblong, and smooth, with convex outer periclinal and clear, straight or slightly curved anticlinal walls. On the inter-rib zones, the cells are also oriented along the longitudinal axis of the fruits, and some cells are rounded, angular, with convex outer periclinal walls having rounded and linear warty cuticular formations.
Figure 8
Fruit of Valeriana sambucifolia J. C. Mikan (SEM): General view (A); fragment of the fruit surface (B,C); stomium (D).

On the inter-rib zones, the stomata are oriented along the longitudinal axis of the fruit. The stomata are surrounded by two–five lateral cells, located along the longitudinal axis of the fruit, and do not differ in shape and orientation from other epidermal cells (Figure 8D). Epicuticular wax is present only in some parts of the surface as unequally divergent platelets of wax. The awns are pubescent, with long twisted soft ribbon-like hairs, and rounded and linear warty cuticular formations.
Valeriana stolonifera Czern. (Figure 9)
The microsculpture of the fruit surface is of the small tuberculate type (Figure 9C,E). The boundaries between the cells are clear. On the ribs, the epidermal cells are arranged in rows along the longitudinal axis, oblong, and smooth, with convex outer periclinal walls; on the inter-rib zones, they are mostly without a clear orientation, mostly rounded, angular, or elongated in places, with convex outer periclinal walls, indistinct or in certain places clear anticlinal walls, and in some areas, papillary formations and papillae up to 30–40 µm long having rounded and linear warty cuticular formations (Figure 9F).
Figure 9
Fruits of Valeriana stolonifera Czern. (SEM): General view, adaxial side (A); general view, abaxial side (B); fragments of the fruit surface (C–F).
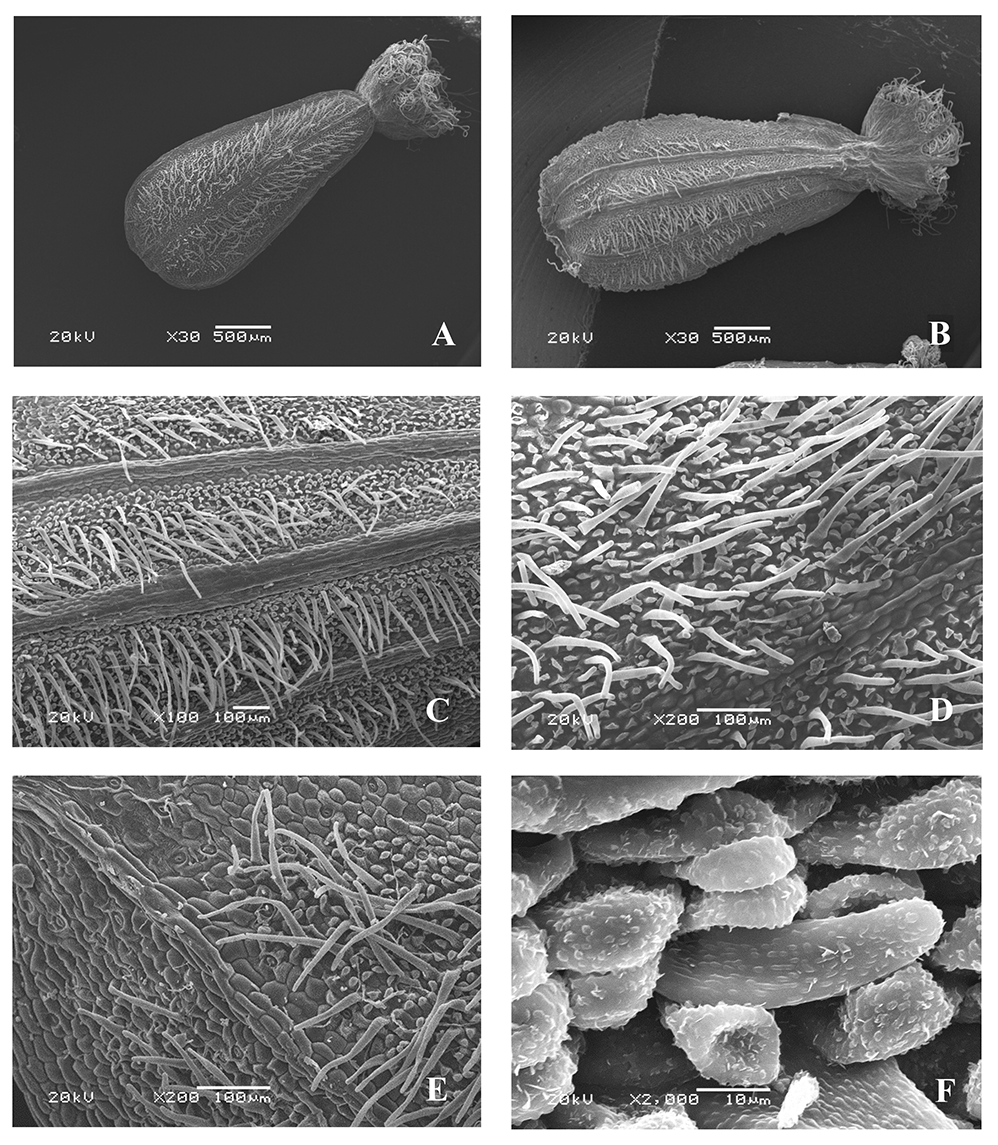
Epicuticular wax is present only in some parts of the surface as unequally divergent platelets of wax. On the adaxial and abaxial sides, the fruits are pubescent only between the ribs, with simple rigid narrow-conical or ribbon-like hairs, up to 0.2 mm long, tubular at the base, and rounded at the apex, with similar cuticular formations (Figure 9D,F). On the inter-rib zones, the stomata are surrounded mainly by five (sometimes two or three) lateral cells, the shape and cuticular sculpture of which are similar to those of other epidermal cells, and the stomatal apertures are parallel to the longitudinal axis of the fruit (Figure 9E). The awns are pubescent, with long twisted soft ribbon-like hairs, and warty cuticles.
Valeriana rossica P. Smirn. (Figure 10)
The microsculpture of the fruit surface is of the small tuberculate type (Figure 10C,D). The boundaries between the cells are clear. On the ribs, the epidermal cells are oblong, with convex outer periclinal walls, smooth, without warty sculpture, and arranged in rows along the longitudinal axis; on the inter-rib zones, they have different shapes – angular, rounded, or elongated, in certain places with straight or slightly curved anticlinal and convex periclinal walls, and with rounded and linear warty cuticular formations.
Figure 10
Fruits of Valeriana rossica P. Smirn. (SEM): General view, adaxial side (A); general view, abaxial side (B); fragments of the fruit (C); stomium (D).

On the adaxial side, the surface (except for the ribs and the border) is pubescent, with simple soft ribbon-like hairs (0.09–0.3 mm long), tubular at the base, rounded at the top, and with warty cuticular sculptures; on the abaxial side, it is not pubescent. The awns are pubescent, with long ribbon-like hairs. On the inter-rib zones, the stomata, oriented along the longitudinal axis of the fruits, are surrounded mainly by five (sometimes two or three) lateral cells, the shape and cuticular sculpture of which are similar to those of other epidermal cells (Figure 10D).
. Discussion
The morphological features of fruits are important for species identification (Mehebub et al., 2019). Considering the micromorphological features of the fruit surfaces of species of the genus Valeriana, we could distinguish three groups of species (which do not correspond to supraspecific taxa): 1 – those with glabrous fruits (V. tripteris, V. simplicifolia, and V. sambucifolia); 2 – those with fruits pubescent only on the adaxial side (V. grossheimii and V. rossica); and 3 – those with fruits pubescent on both sides (V. tuberosa, V. wolgensis, V. stolonifera, and V. officinalis).
SEM analysis showed that within these groups of species, the microstructure of the fruit surface is similar in terms of the shape of epidermal cells, the presence and shape of cuticular formations on the surface of the outer periclinal walls of epidermal cells and on the surface of hairs, the type and shape of hairs (in pubescent species), and the shape of epicuticular waxy formations, which are present mainly on hairless surfaces; however, there are also some differences between them (Table 1).
Table 1
Main differences in microstructure of fruit surfaces of studied Valeriana spp.
Valeriana wolgensis, V. stolonifera, and V. officinalis have a similar surface microstructure of fruits and belong to the collective polymorphous species V. officinalis s. l. (Gorbunov, 2002). These species are also similar in morphology of vegetative and generative organs; hence, it is understandable that a slight species differentiation manifests at the level of micromorphology on the fruit surface.
However, Karcz (1996), who studied the micromorphological features of the fruit surface of V. officinalis (the author considered the species in a broad sense), found some differences between two varieties (V. officinalis var. officinalis and V. officinalis var. nitida). The micromorphological differences were noted for the structure of cuticular formations on the surface of epidermal cells and the surface of hairs, as well as the presence of epicuticular wax. Karcz (1996) did not rule out that these characteristics are phenotypic, resulting from the conditions prevailing in their habitat. We analyzed samples of V. officinalis fruits that were taken from herbarium material (KW) (Table S1) and identified as V. officinalis var. nitida, V. officinalis s. str., and V. wolgensis (which we consider synonymous with V. officinalis var. nitida). There were no differences between the studied samples. Phenotypic variability can probably manifest in the presence and localization of pubescence of fruits of some species. The samples of V. grossheimii studied by us had pubescence of fruits only on the adaxial side; however, Gorbunov (2002) indicated pubescence for this species only on the abaxial side. Fruit samples of V. officinalis s. str. were pubescent on both sides, and according to the literature, the fruits can also be glabrous or pubescent on one side only (Grubov, 1958). However, to identify the species, it is necessary to consider the whole complex of features of the fruit surface microstructure, except the type of wax formation on the epidermal cell surface, which cannot be used as an additional diagnostic feature, because it is similar in all the studied species of the genus. At the supraspecific level, the revealed features of the fruit surface somewhat overlap and can be used to identify series and sections only as additional features.
Thus, an investigation on the microstructure of the fruit surfaces of Valeriana spp. was conducted, and the diversity in the cuticular sculpture of the outer periclinal cell walls of the epidermis was shown. In turn, the micromorphological features were identified, and these can be used as additional features for the identification of species and supraspecific taxa.
However, our data on the microstructure of the fruit surface for some Valeriana spp., samples of which were collected from the territory of Ukraine, differ slightly from those reported by other scientific sources. This study revealed detailed micromorphological characteristics of the fruit surfaces of all the studied species, and they may be useful for further comparative morphological studies of fruits taken from other aerial parts of the species. These will help identify stable micromorphological characteristics of fruits that are resistant to geographical and environmental factors.
