. Introduction
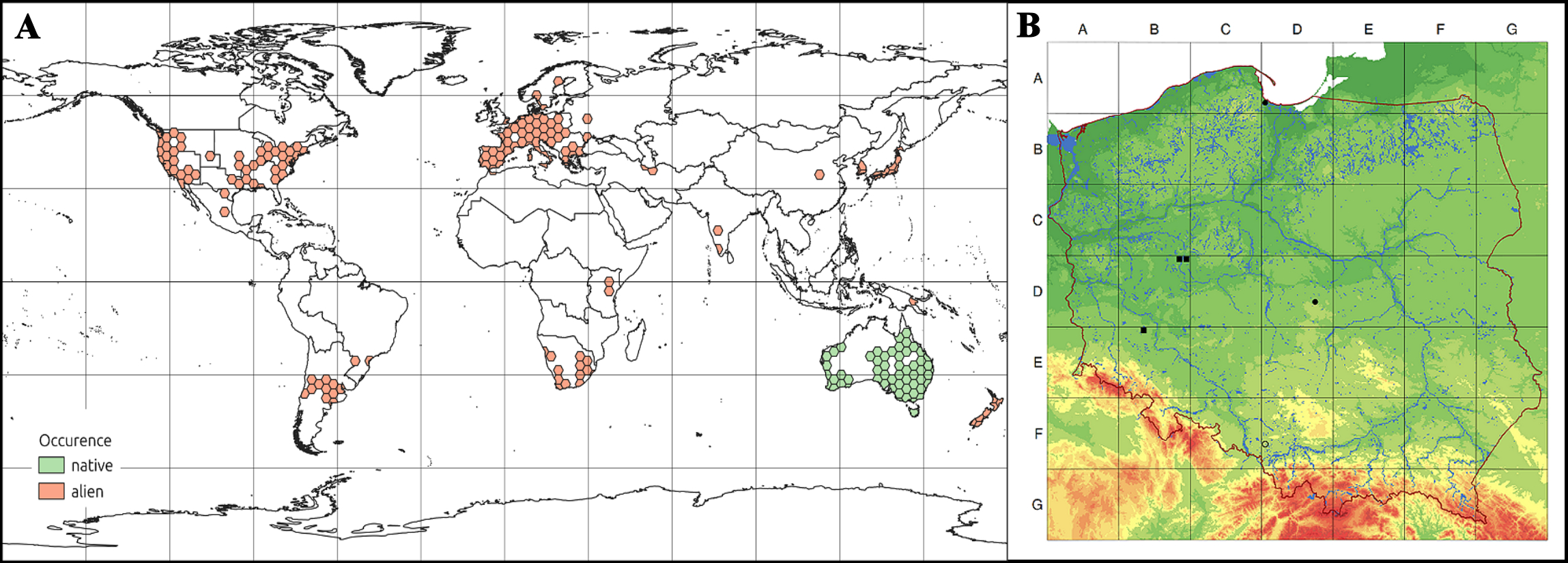
Dysphania pumilio (R.Br.) Mosyakin & Clemants (Chenopodium pumilio R.Br.; Amaranthaceae) is native and common in southern Australia and Tasmania, and has been unintentionally introduced to all continents but Antarctica (GBiF, 2024; POWO, 2024; Uotila et al., 2021; Figure 1A). This very vigorous annual species may be a troublesome weed.
Figure 1
Distribution maps of Dysphania pumilio (R.Br.) Mosyakin & Clemants. (A) general; (B) in Poland.
Within the genus, it is distinguished by vertical seeds, perianth segments rounded abaxially, tepals pallid, whitish to white, and free to near base, numerous flowers in inflorescences, inflorescences in the form of lateral cymes or glomerules up to 5 mm in diameter. The lens-like fruits develop in 4–5-lobed herbaceous, green, pale green, pale yellow, rarely slightly pink perigones, the lobes of the perigone do not fit tightly and dull fruits inside are visible; stems and leaves are sparsely pilulose with segmented hairs and sessile or stipitate glandular hairs; leaves petiolate with blade elliptic to ovate or rhomboid-ovate, 1–3 cm long, with sinuate margins (Clemants & Mosyakin, 2003; Dostálek et al., 1990; Rutkowski, 2022; Wilson, 1984; Figure 2A–C).
Figure 2
Morphology of Dysphania pumilio (R.Br.) Mosyakin & Clemants. (A) plants with erected stems; (B) top of the stem; (C) cluster of flowers, visible fruits and flowers in various stages of development; (D) dominating prostrate form on vegetable bed, general view; (E) the smallest flowering plants; (photo Marcin Bortniak, 2023.10.14).

Its primary habitats in the native range are difficult to assess. Probably, it occurs on periodically drying riverbanks and shores of reservoirs, and occasionally in saline soils. Human activity caused it to spread to secondary disturbed habitats; moreover, it commonly occurs as an agricultural weed (Wilson, 1984). According to the Australian HerbiGuide (http://www.herbiguide.com.au/Home.aspx), it is toxic to sheep, allelopathic, and relatively tolerant to low rates of glyphosate.
The species is thought to be introduced to Europe with Australian wool (Ascherson, 1896; as Chenopodium carinatum R.Br.). The first records were made in 1889 in Döhren in Germany (Aellen, 1979) and in 1890 in Nosislav in the current Czech Republic (Lhotská & Hejný, 1979). Since then it has spread and been reported in almost all European countries (POWO, 2024). In Germany, the Czech Republic, and Slovakia, it is considered to be a naturalized but not invasive species and is associated with plant communities of weeds and terophytes of the classes Stellarietea mediae Tx. et al. in Tx. 1950 (current Papaveretea rhoeadis S. Brullo et al. 2001) and Bidentetea tripartitae Tx.et al ex von Rochow 1951 (Dostálek et al., 1990; Oberdorfer, 2004; Pyšek et al., 2012).
In Poland, D. pumilio was very rare and classified as an ephemeral species (Rutkowski, 2022) or a locally established neophyte (Tokarska-Guzik et al., 2014). It was recorded only in three sites, all on roadsides or wastelands: (1) the port in Gdańsk (the first observation in 1974), where it survived for decades although its place of occurrence has changed (Misiewicz & Korczyński, 2003; Urbisz, 2011). In 2020, the species was also collected in the second site in Gdańsk-Wrzeszcz (GBIF, 2024). (2) Rybnik-Piaski; three individuals were noted in 1992 on roadsides; this site was destroyed (Urbisz, 2011; Witosławski, 2009). (3) Stryków, over a dozen individuals were observed in 2005; the species survived to 2008 when it was almost destroyed, and only two plants occurred in 2009 (Witosławski, 2009); in 2023, a photograph of the species from a site located about 6 km south of Stryków was added to iNaturalist (https://www.inaturalist.org/).
In the last years, the species was observed in two new locations. One site was found in Poznań in 2018, and the second was discovered in Rudna village (Lower Silesia, SW Poland) in 2023 (Figure 1B). For the first time in Poland, it occurred as a weed in cultivated areas.
We undertook investigations to (1) determine the sources of seeds or plants and the way of D. pumilio spreading, (2) determine the ability of the species to overwinter in Poland, (3) check the production and viability of seeds, and (4) assess the impact of the species on the existing synanthropic vegetation and its ability to penetrate other communities. Based on these studies, we attempted to determine the invasive potential of D. pumilio in Poland.
. Material and methods
The object of our research were two numerous populations of Dysphania pumilio.
Population in Poznań; Daleka Street, Poznań–Górczyn; 16.887678 E, 52.383153 N. It is associated with urban ornamental perennial and shrub beds. The first plants were observed in 2018, shortly after the renovation of the greenery and new plantings in the area. The species has been present there since then. Due to the likelihood that the species was transferred with the nursery material, the research covered ornamental plant nurseries in the vicinity of Poznań.
Population in Rudna; 16.2610 E, 51.5042 N. The species occurs in a cultivated vegetable garden. When it was found in 2023, it already occupied ca. 70 m2.
The determination of the species was based on Clemants & Mosyakin (2003), Dostálek et al. (1990), Rutkowski (2022), and Wilson (1984). Three voucher specimens of D. pumilio collected in Rudna were deposited in the Herbarium of the University of Wrocław.
Distribution maps of all the species occurring in Poland were made in the ATPOL grid of 10×10 km (Zając & Zając, 2019). To prepare a global distribution map, the species occurrence data was taken in January 2024 from GBIF, using the rgbif package (Chamberlain et al., 2023). Data sets were cleaned with the CoordinateCleaner package (Zizka et al., 2022) and mapped to a hexagon grid with a function to count the points (species occurrence) in the polygon. World distribution maps of the species were prepared in QGIS (http://qgis.osgeo.org).
The origin of the populations was determined on the basis of interviews with land owners and managers and research in adjacent areas.
To determine the species overwintering capacity in Poland, we checked the lowest winter temperatures recorded in both locations during the period of the presence of the populations. Due to the lack of a measuring station in the vicinity of Rudna, the temperature data is based on ERA5 hourly data on single levels from 1940 to present, Copernicus Climate Change Service (https://cds.climate.copernicus.eu/).
Since mechanical and chemical treatments had been used to limit the occurrence of D. pumilio at the site in Poznań, the research on the density and features of plants as well as the size of the seed bank and viability of the seeds was carried out only on the population from Rudna.
The density of individuals was counted on ten 0.5×0.5 m test plots. The population size was estimated based on the average density of plants in the test plots multiplied by the area inhabited by the population.
The length of the main shoot was taken as the plant size. This feature was measured for erected and prostrate plants from two spontaneous germination events: the main one in late spring and summer and the much rarer autumn one.
The number of inflorescences was calculated per 10 cm of the central part of shoot for 30 shoots of various lengths, and the result was averaged.
The number of flowers in 60 inflorescences was counted for glomerules along the entire length of five shoots.
The size of the seed bank was tested on 10 plots 0.1×0.1 m. Ten samples of deposited seeds were collected with a vacuum cleaner, and the seeds were separated and weighed. Then, 1 000 seeds were selected for each sample and weighed, and the number of seeds in a given sample was calculated based on their weight.
The viability of the seeds was tested on five samples of 100 seeds. The first part of matured seeds was collected from green plants in October 2023. One sample was sown immediately after harvesting, and two others were sown after 14 days of stratification at 5ºC (one sample – dry stratification, one in wet substratum). The second part of seeds was taken from the seed bank collected in February 2024. All samples were sown in a greenhouse on sterile substrate at room temperature.
Phytosociological relevés were prepared according to the Braun-Blanquet method (Westhoff & van der Maarel, 1978). The area occupied by D. pumilio and the distinctive patches of communities containing the species allowed only four phytosociological relevés. One plot was documented on 22.08.2022 in Poznań (relevé 1), and three relevés (2–4, respectively) were prepared 14.10.2023 in Rudna. Relevé 3 was prepared in the only plot with dominant D. pumilio, and relevés 3 and 4 were made in weed communities where D. pumilio appears with a small coverage. The nomenclature of species was adopted for POWO (https://powo.science.kew.org/). The nomenclature of syntaxa, affiliation, and diagnostic species are consistent with FloraVeg.EU (https://floraveg.eu/). The phytosociological relevés were included into the Polish Vegetation Database of the University of Wrocław (https://www.givd.info/ID/EU-PL-001).
The invasiveness potential of Dysphania pumilio in Poland was calculated according to the Weber & Gut method (Weber & Gut, 2004).
. Results
Population in Poznań. The first record: 22.05.2018, Poznań, Daleka St., ATPOL BD0856; approximately 100-m long part of the pavement; plants grew between paving stones on the driveways and at the edges of the shrub beds. The species has been present there every year, but the population rapidly decreased in 2019 after cleaning and chemical treatment and was restored in next the years. In 2023, the species was observed on ca. 3 m2. The pattern of its occurrence suggests that the seeds were transferred with the substratum used to renovate the plant beds. In 2022, the species was also noted here in balcony pots on the fourth floor of a building at 7 Daleka St. The seeds may have been transported with the substratum; however, we cannot exclude anemochory.
Moreover, on 12.08.2022, D. pumilio was also found in one ornamental plant nursery in the vicinity of Poznań, i.e. in the nursery of perennials in Dobrepole, Poznań-Spławie, ATPOL BD0995. A few individuals were growing in the cracks of concrete slabs between pots with ornamental plants. The mode and time of arrival has not been specified. We did not observe the species growing in the pots; however, transport of the seeds with the nursery stock cannot be excluded.
Population in Rudna. In 2023, Dysphania pumilio was found in a private garden (Figure 2D) in the southern part of the village, ATPOL BE0319. The soil there is sandy and dry and the garden is extensively cultivated. According to the information obtained from the owner, it probably appeared about 10 years ago, and was introduced with herb seeds used to start a crop in containers. The containers were partially buried in the ground and their total area was 1 m2. The soil used in cultivation came from this garden. After four years, the cultivation and containers were removed, and the soil with the seed bank remained in the garden. The species has been spreading in the garden for about six years. In 2023, the species was present in two areas: the first one covered about 60 m2 including the site of a former herb bed and its vicinity, while the second plot was smaller and covered about 10 m2. It was located about 20 m west of the primary site and the seeds were probably transferred on the tiller. The species spread in the cultivated part, did not penetrate the adjacent lawn, and did not spread to the field in the immediate vicinity of the garden.
The lowest winter temperatures noted in last six years in Poznań and in Rudna were –17.6 and –16°C, respectively (Table 1).
Table 1
Minimum winter temperature noted in Poznań and Rudna in years 2018–2023 in ºC.
| 2018 | 2019 | 2020 | 2021 | 2022 | 2023 | |
|---|---|---|---|---|---|---|
| Poznań | -14.6 | -8.2 | -6.1 | -17.6 | -9.3 | -9.9 |
| Rudna | -14.9 | -9.7 | -6.1 | -16.0 | -8.1 | -8.1 |
On the 10 tested plots in Rudna, from 1 to 43 plants of D. pumilio per 0.25 m2 were noted, 13.6 on average, which means ca. 54.5 plants per 1 m2. We estimated the entire population at about 3800 individuals. The area occupied by a single plant depended on its size and shape. The smallest and erected plants covered about 1 cm2, whereas large and prostrate or almost creeping individuals covered up to 0.5 m2. Erect and prostrate forms were observed in both locations. Prostrate to creeping plants dominated in trampled places, e.g. paths in the garden in Rudna and pavements in Poznań, but were also observed in non-trampled areas.
Morphological features
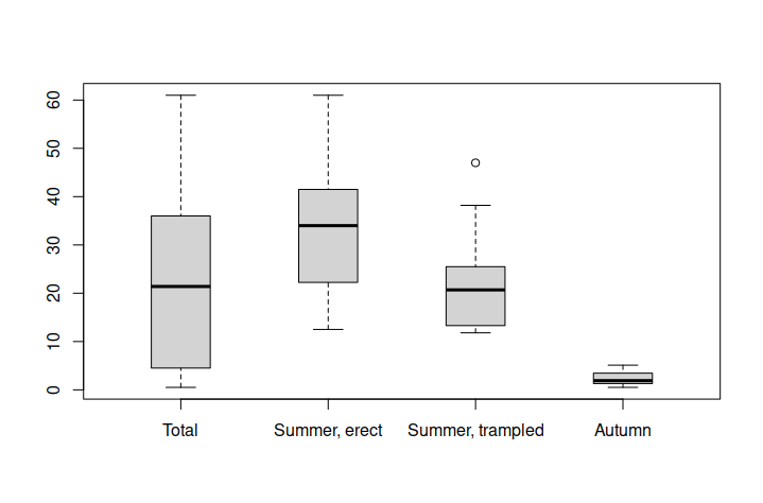
The sizes of the main shoots of mature plants varied greatly. Their high/length ranged from 0.5 (Figure 2E) to 61 cm, 21.4 on average (Figure 3). This difference is an effect of reaction to two factors: trampling and time of sowing/germination. Plants growing in summer were bigger, often numerously branched. Erect non-trampled plants reached sizes of 12.5 to 61 cm, 34 on average, whereas trampled plants were smaller, i.e. from 11.8 to 38.5 cm, 20.7 on average. Plants germinating in early autumn occurred on open soil between others, were less numerous, and did not exceed 5.5 cm (1.9 in average).
Figure 3
Size of the longest stem of Dysphania pumilio (R.Br.) Mosyakin & Clemants; general, plants germinated in spring erect and trampled, plants germinated in early autumn.
The density of inflorescences on a shoot increased from the base of the shoot to its top. In the lower part of the stem, the glomerules were rather sparsely located approximately 15 mm apart. At the top of the shoot, the distances were about 1 mm, and the glomerules looked like a continuous structure. On average, ca. 17 inflorescences developed per 10 cm of shoot.
The number of flowers in the inflorescence varied along the stem. The smallest glomerules with 1–3 flowers developed in the lower parts of the stems. The glomerules in the central and upper part consisted of 10–20 flowers, up to 41 in the upper part of the stem. On average, there were 18 flowers in one inflorescence. The flowers developed gradually. In the same inflorescence, there were achene-type fruits with mature seeds, mature flowers, and flower buds (Figure 2C).
Seed production and viability
The fruits fell mainly under the mother plant (Figure 4A). 1000 fruits weighed from 0.16 to 0.222 g., on average 0.1871. In the researched 10 × 10 cm plots, we found from 3623 to 20964 seeds, 12477 on average. The fruits fell in perigones (Figure 4B).
Figure 4
Fruitification of Dysphania pumilio (R.Br.) Mosyakin & Clemants. (A ) plant remnants with a layer of fallen fruits; (B) close-up of fruit layer; (photo M. Bortniak, 2024.02.03).

Seeds collected in October 2023 and sown immediately did not germinate at all. In February, three autumn samples of seeds were sown (1) without stratification and with stratification at 5ºC in dry (2) and wet (3) substratum. In total, 17%, 0%, and 75% of the seeds germinated, respectively. Seeds collected in February in the field germinated at the 37% rate. Germination occurred mainly within seven days, with no or slight changes over the next seven days (Table 2).
Table 2
Seed germination. (A) seeds collected in October 2023 from plants; (B) seeds collected in February 2024 from seed bank; (1) seeds sown after collecting; (2) seeds sown without stratification; (3) seeds after dry stratification; (4) seeds after wet stratification; 7, 14, 28 – days of observation period.
| A | B | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||||||
| Observation period | 7 | 14 | 28 | 7 | 14 | 28 | 7 | 14 | 28 |
| Germination in % | 0 | 16 | 17 | 0 | 75 | 75 | 35 | 37 | 37 |
Plant communities with Dysphania pumilio
In general, Dysphania pumilio is classified as a diagnostic species of the class Digitario sanguinalis-Eragrostietea minoris Mucina, Lososová et Šilc in Mucina et al. 2016 including thermophilous grass-rich anthropogenic vegetation native to the Mediterranean area (FloraVeg.EU 2024). In Poland, the species was found in two types of plant communities. In Poznań, most of the population was noted in an unclassified plant community. A poor thermophilic and trampled plot developed on the roadside and in the driveway. The plants grew between paving stones. In the plot, eight vascular species (including 4 neophytes), seven moss species, and one liverwort species were noted (Table 3, relevé 1).
In Rudna, D. pumilio occurred in a typical weed community of the class Papaveretea rhoeadis (= Stellarietea mediae; Table 3, 2–4). One relevé was prepared in an area with the dominance of D. pumilio and two relevés were made in an area where the species is in the expansion phase. In all the plots, only the herb layer occurred, and the number of noted vascular plants was 15, 30, and 33, respectively. Galinsoga parviflora was the dominating or co-dominating species in all the plots, with at least one significant coverage of Digitaria sanguinalis and Convolvulus arvensis.
Table 3
Plant communities with Dysphania pumilio (R.Br.) Mosyakin & Clemants. Phytosociological relevés (1) Poznań, Daleka St., 16.8876 E, 52.3831 N; (2)–(4) Rudna, Młynarska St., (2) 16.261060 E, 51.504241 N, (3) 16.2609271 E, 51.5042439 N, (4) 16.260553 E, 51.504283 N.
In all the plots, about half of the species were alien taxa, 7 (2 archaeophytes and 5 neophytes), 12 (6 archaeophytes and 6 neophytes), and 20 (14 archaeophytes and 6 neophytes), respectively. More than half of the species noted in the plots were represented by only a single plant (7/15, 16/30, and 21/33, respectively). Almost all the recorded taxa are terophytes, while perennial plants occurred sporadically and only as seedlings.
Competitiveness and potential invasiveness of Dysphania pumilio
Based on the phytosociological documentation, it can be concluded that D. pumilio is able to replace existing weed species. The process is rather slow, affects areas in the immediate vicinity of the place of introduction, and is effective mainly against other annual species. Dysphania pumilio has not penetrated dense communities formed by perennials.
To summarize our research, we attempted to determine the invasiveness potential of D. pumilio in Poland. According to the protocol developed by Weber & Gut (2004), the species received high scores in nine of 12 categories analyzed (Table 4). However, three categories of high importance have a value of zero. The lack of vegetative reproduction (category 8), being a small annual plant which means low competitiveness (category 10), and the lack of data on its occurrence in particularly sensitive ecosystems (category 11) caused the overall assessment of the species’ invasiveness to be relatively low. The invasiveness risk class of Dysphania pumilio was assessed as intermediate; nevertheless, the species requires further observation.
Table 4
Risk of potential invasiveness of Dysphania pumilio (R.Br.) Mosyakin & Clemants, assessment according to protocol by Weber & Gut (2004).
. Discussion
Dysphania pumilio is one of four species of the Dysphania genus noted in Poland. Unlike other species, it was never used medicinally and had no importance in traditional rituals (Dagni et al., 2022; Łuczaj et al., 2022). It was not introduced intentionally or cultivated.
It is a species with high ecological plasticity, easily colonizing a variety of disturbed habitats. Its tolerance of saline soils makes it able to inhabit roadsides and some urban habitats, which are difficult to access for most native species due to the presence of salt destroying seeds and seedlings.
The species seems to be expanding on a global scale; however, its distribution and especially new appearances seem not to be related to ongoing spontaneous expansion but to accidental transport by humans. New localities appear in almost all parts of the world, from where it continuous to spread or not. After 2000, it appeared in the Old World in some new and isolated sites, i.e., in China (Zhu & Zhu, 2006), in the western part of Iran (Rahiminejad et al., 2004), or the southern part of India (Kambhar et al., 2017). In Europe, the species has been present in some countries longer than 100 years (Germany, Czech Republic), although their climate conditions did not seem favorable. On the other hand, its sites in much more favorable habitats are rather new finds, e.g. in the Balkan Peninsula. In the last years, it has been reported as a new species in Albania, Greece, Kosovo, and Serbia (Barina et al., 2020; Bogosavljević & Zlatković, 2017; Vladimirov et al., 2013).
The species spreads only by seeds. Its primary habitats and the presence of a light perigone surrounding the fruit suggest hydrochory and/or anemochory as an original mode of spreading. Nowadays in Australia, the seeds are transported directly by epichory (mainly by sheep). Within the secondary range, the species is not dependent on one type of spreading. In Europe, autochory, chamaechory, endochory, hydrochory, and different forms of anthropochory have been observed (Lhotská & Hejný, 1979). Such numerous forms of dispersal indicate random rather than specialized vectors.
In Poland, the species is a relatively new incomer. The oldest and still existing site has been standing for 50 years (Witosławski, 2009). The other known localities have also existed for at least several years, and the species slowly spreads. The exception is the site in Rybnik; however, the species did not disappear there due to natural causes, but the population was destroyed by construction works (Urbisz, 2011).
The seed sources varied. They arrived to the port with imported articles in Gdańsk, with prepared garden substratum used for renovation of shrub beds in Poznań, and with a mix of herbal seeds in Rudna. The primary sources of the last two populations as well as the populations in Stryków and Rybnik are not known (Witosławski, 2009). The species is likely to spread along roads where seed can be transported directly by means of vehicles or picked up and carried by the wind.
The seeds germinate at light and high temperatures. Seedlings appear when the soil is sufficiently heated at the end of May or beginning of June (Lhotská & Hejný, 1979). According to vegetation phenology, it is full spring and early summer. Plants start bearing fruit quickly and continue until the first frost. Global warming has caused favorable thermal conditions to occur earlier and last longer, which extends the time of ongoing growth and fruiting. Cooling of seeds promotes the germination process. Lhotská & Hejný (1979) cooled seeds for 30 days at 10ºC, and we applied a temperature of 5ºC for 14 days; the seeds germinated with a high percentage in both experiments. Temperatures below 0ºC reduce the ability of seeds to germinate. The tested seeds from Rudna germinated at 75% when cooled, while the seeds collected in field after 8ºC frost germinated at 37%. However, the confirmed frost resistance of the seeds reached at least 15ºC below 0. This means that, in the current climatic conditions, winter temperature in Polish lowlands is not a factor that can destroy the entire seed pool.
Experiments on germination conducted in the Czech Republic in the 1970s showed that seed viability varied greatly in different populations, ranging from 0 to 82% (Lhotská & Hejný, 1979). We observed a similar range in one tested population, depending on the habitat conditions. Particularly interesting is the lack of germination immediately after harvest, which suggests that the seeds really quickly started the dormancy period. The seed viability of the population in Rudna is at least 75%, which was recorded in the sample subjected to cooling (Table 2).
The chances of spreading are increased by the large number of produced seeds. As we estimated in the population from Rudna, a medium-size plant with a total length of shoots ca. 200 cm (5 shoots 10 to 21 cm long and secondary short shoots 1–3 cm long) produces about 6000 seeds. With the confirmed field germination rates, it potentially gives at least 2300 new plants.
Dysphania pumilio can disperse by fruits in the perigone, naked fruits, fragments of plants with fruits, and entire plants with fruits. In the Czech Republic, 70% of observed populations were associated with dumps and heaps of refuse, trampled areas, and railway areas (Lhotská & Hejný, 1979). This suggests that the species disperses mostly by various kinds of anthropochory. Moreover, diaspores remain all winter on a dry plant. In Poland, in winter 2023/24, all the fruit fell in January, greatly reducing the chances of spreading. The seeds fell directly under the mother plants.
The population in Rudna occurred in a fenced and extensively cultivated garden. As we observed, the spread of the species was rather slow. Over six years, the occupied area increased from 1 m2 to 70 m2, and the plants moved away from the starting point by a maximum of 25 m. The highest density of plants was observed in the beds directly at the site of the species introduction. This all indicates the lack of an efficient transport vector. The population in Poznań is located on and near the city pavement and roadside, where the traffic of people and cars may be a transport factor itself or facilitate anemochory. In 2018, the population occupied an area of ca 100 m2 and was reduced to ca. 3 m2 in 2023. Plants appearing there were treated as weeds and destroyed mechanically and chemically. Despite this, the species managed to survive.
Seedlings of D. pumilio developed on bared substratum in open stands of initial stages of vegetation (Lhotská & Hejný, 1979). The species seems to be little competitive for existing living plants, but it effectively uses gaps in the plant cover. In the observed populations, the species appeared on substratum without natural or seminatural vegetation, i.e. without competitive perennial plants, and did not enter the extensive lawn in the garden. Greater resistance to drought and high temperatures may facilitate the species penetration into native communities disintegrated by climate changes. Interestingly, it was unable to colonize a cultivated field located nearby, although it was observed in such habitats in other countries.
Based on our research, we attempted to determine the invasiveness risk of D. pumilio as intermediate; however, its total score had the lowest value of this category (Weber & Gut, 2004). Moreover, we can conclude that, until an effective seed dispersal factor appears, the risk of intensive expansion of the species is rather low.
. Conclusions
The Australian species Dysphania pumilio is a neophyte locally established in Poland. The species is able to slowly but effectively replace other weeds in cultivated areas; however, it does not penetrate natural or seminatural communities. The frost resistance of its seeds is sufficient to survive winters in Poland. The plants produce a large number of highly viable seeds, but there is no factor that could effectively contribute to their spread. It seems more likely to spread along roads than as a weed in crops. The current risk of invasion is rather low, but the species requires further observation.
