. Introduction
Chlorophyll (Chl) is an important pigment for plant photosynthesis (Green & Durnford, 1996). The dissociation of Chl is a necessary step for chloroplast degradation and provides nutrients for new leaves or developing organs (Shin et al., 2020). The decrease in Chl contents would increase the redness parameter of maple leaves (Primka & Smith, 2019). The color-break in grape berry was the visual manifestation of the developmentally regulated transition and often involves the massive breakdown of Chl (Giovanelli & Brenna, 2007; Kamffer et al., 2010). The yellowing of cigar leaves during the airing process of the flue-cured cigar was caused by the regulation of chlorophyllase 1 (CLH1) protein on the synthesis and decomposition of Chl, delaying the degradation of lutein and accelerating the decomposition of Chl (Wu et al., 2020). The difference in contents of Chl, and anthocyanin might be the main reason for the different leaf color performance of triangular maple (Rong et al., 2011). The ratio of anthocyanin and its various pigments can also lead to the color change of red-leaf camellia, and the red-green value a* could be used as a representative parameter to describe the leaf color change of red-leaf camellia (Pan et al., 2020). Carotenoids are mainly synthesized in chloroplasts and chromoplasts and widely participate in photosynthesis and pigment deposition of plants, thus regulating plant growth and development (Sandmann, 2021). The most direct factor for the change in the leaf color of begonia was the change in the ratio of the total chlorophyll to carotenoid (TC/Car) (Walibai et al., 2017). Therefore, the degradation of chlorophyll has a great impact on the change in leaf color.
Chl degradation includes two types of reactions: enzymatic degradation and photo-oxidation (Hendry et al., 1987). Generally, Chl degradation was regulated by varieties of enzymes (Schelbert et al., 2009; Shemer et al., 2008), which was firstly a depletion reaction under the catalysis of chlorophyllase (CLH), and secondly pheophorbide a (Pheide a) is formed by the removal of magnesium atoms by the action of Mg-dechelatase (MDCase) in the absence of plant-based chlorophyll a (Chl a) (Hörtensteiner, 2006). However, pheophytin pheophorbide hydrolase (PPH) could only specifically catalyze pheophytin a and b, and could not directly act on Chl a and Chl b (Schelbert et al., 2009). Therefore, Chl b was converted to Chl a by Chl b reductase, which was encoded by non-yellow coloring 1 (NYC1) and NYC1-like (NOL), and by 7-hydroxymethyl Chl a reductase (HCAR) (Horie et al., 2009; Kusaba et al., 2007; Meguro et al., 2011), Phein a was then converted to Pheide a by PPH. The porphyrin ring of Pheide a was cleaved by pheophorbide an oxygenase (PAO). The difference between the two pathways of Chl degradation lay in the sequence of pheophytin (Gómez-Lobato et al., 2014). During this process, the Chl degradation products showed different color changes. The color of Phein a produced by Chl a via the action of MDCase was olive green, but the color of Chlide a produced by the action of CLH is bright green. Therefore, the different pathways of Chl degradation might also be the main reason for the difference in the color of plant leaves.
Magnesium (Mg), as the central atom of the Chlporphyrin ring, affects a series of physiological and biochemical processes such as Chl synthesis and carbohydrate partitioning in plants (Beale, 1999). Chl degradation is affected by mounts of environmental factors such as fertilization (Pranckietiene et al., 2020), light (Mes et al., 2008), pH (Ono et al., 2019), temperature (Van Loey et al., 1998). Due to magnesium being an important component of chlorophyll, these environmental factors are likely to affect the Mg content in plant leaves, especially the application of Mg fertilizers. Foliar spraying of 0.27% Mg fertilizer had the best effect on improving the pigment content of wine grape Cabernet Sauvignon leaves (Gu et al., 2021). An appropriate amount of Mg application in Mg-deficient soils could significantly increase the contents of Mg and Chl in leaves and increase the net photosynthetic rate (Ma et al., 2018). Low Mg contents would hinder key enzyme activities and cytochrome synthesis in plant metabolism (Yin et al., 2009). Mg regulates the energy distribution between photosystem I (PS I) and photosystem II (PS II) of chloroplasts, effectively protects Chl structure (Tikkanen et al., 2014), improves Chl’s tolerance to strong light, and promotes the absorption and utilization of light energy (Farhat et al., 2015). Both excess and deficiency of Mg can cause abnormal chloroplast structure and shape, which affects the function of the chloroplast light reaction center, resulting in a decrease in Chl content and abnormal yellowing of leaf external morphology (Hermans et al., 2004). Therefore, the effect of Mg on the degradation of Chl in plants is particularly important. Leaf color is related to the endoplasmic pigment of plant leaves, and the degradation rate of Chl and Car had a direct impact on the appearance quality of plant leaves (Falcone et al., 2012).
Cigars were a special kind of tobacco products (Zhang et al., 2014). The main function of cigar wrappers was to improve the aesthetics of cigars (Gao et al., 2019), and their color and uniformity were the most important indicators for evaluating the quality of cigars (Ding et al., 2011). However, there are few reports on the effect of magnesium on leaf color in chlorophyll degradation. Therefore, this study conducted field experiments to study the changes of MgO fertilizer on the chromaticity values, pigment contents, and Chl-degrading enzyme activities of middle leaves of cigar tobacco plants, to determine the color formation mechanisms of cigar tobacco leaves, and the relationship between the degradation pathway of Chl by Mg and the formation of leaf color.
. Material and methods
. Field experiment
The field experiment was carried out in Cuiba Town, Enshi City, Hubei Province (30°28′8″N, 109°47′54″E) in 2021. Fields with moderate and uniform soil fertility and easy irrigation were selected. The basic meteorological conditions of the field test: the annual average rainfall is 1535 mm, the altitude is 600 m, and the annual sunshine time is 1300 h. Soil physical and chemical properties: pH 6.23, organic matter 24.93 g/kg, total nitrogen 1.55 mg/kg, alkaline hydrolyzable nitrogen 127.04 mg/kg, total phosphorus 1.19 mg/kg, available phosphorus 36.6 mg/kg, available potassium 306.2 mg/kg. The field management was carried out in accordance with the local technical specifications for high-quality cigar tobacco leaves production.
Cigar CX26 tobacco leaves (Folium cigarum) were used as the test variety, trials were set up with three treatments. Taking no fertilizer as control (CK), nitrogen - phosphate - potash fertilizers (NPK) and NPK fertilizers + 75 kg/hm2 MgO fertilizer (NKP+Mg) were set as different fertilizer treatments. NPK fertilizers were applied with a special compound fertilizer for tobacco at 119.4 kg N/hm2, 59.7 kg P2O5/hm2, and 358.2 kg K2O/hm2, respectively. The tobacco seedlings were cultivated in the special nursery of the base for two months. Transplant the well-raised cigar tobacco plants into the field, and apply MgO fertilizer at the tobacco plant cluster stage to ensure that the amount of MgO fertilizer applied to the cigar tobacco plants in each plot was consistent. Cigar tobacco plants went through four stages in the field growth period: seedling returning stage, root extension stage, vigorous growth stage, and mature stage, lasting for 80 days. They would be picked uniformly when the tobacco leaves in the middle were mature.
. Airing cigar tobacco leaves
The middle tobacco leaves from each treatment were placed in a special adjustable airing room for wrappers for tobacco airing. The same airing room was filled at one time, and the airing technology was the same to ensure that the temperature and humidity in the airing room were basically the same during the airing process. The temperature and humidity controllers were used to control the temperature and humidity changes in the airing room. The curing process of tobacco leaves successively went through four stages: withering stage (0–2 d), yellowing stage (2–6 d), browning stage (6–12 d), and dry gluten stage (12–30 d). During the withering period, ensure that the ambient humidity of the airing room was 95%, the yellowing period was 90%, the browning period was 85%, and the dry gluten period was 80%. The humidity was detected and recorded by the humidity recorder. Samples were taken during the airing process, and the leaves were collected at 0, 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30 d, and taken back to the laboratory to measure the chroma of the tobacco leaves immediately. The collected leaves were frozen with liquid nitrogen and stored in an ultra-low temperature refrigerator at −80 °C for the determination of pigment and enzyme activity (Pre-airing: 0, 2, 4, 6, 8, 10 d).
. Determination method
The Mg contents in leaves were measured following the method of Wang et al. (2021), and the water contents were measured refer to Qin et al. (2022). The chromaticity values (luminance value L*, red-green value a*, yellow-blue value b*, and total color difference value E) of the leaves were measured with an colorimeter following the reference of León et al. (2007). Enzyme activities (MDCase, CLH, PPH) determination refers to the method of Chen et al. (2012), Yang et al. (2009), Suzuki et al. (2005), respectively. The activities of pheophorbide a oxygenase (PAO) were determined according to the product manual of the ELISA kit. The absorbance of Chl a, b, and Car was determined by UV spectrophotometer at wavelengths of 665 nm (D665), 649 nm (D649), and 470 nm (D470), respectively. Calculated according to the following equations. CChl a, CChl b, CCar, and CTC represent the concentration of Chl a, Chl b, Car, and TC respectively.
. Results
. The Mg contents of cigar tobacco leaves
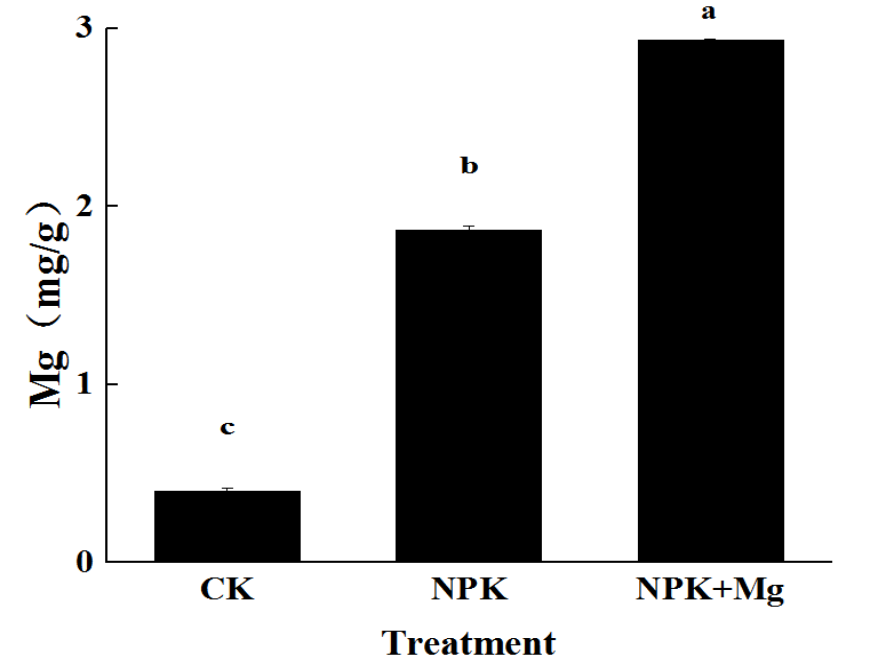
The Mg contents of cigar tobacco leaves treated with CK, NPK, and NPK+Mg were 0.4, 1.9, and 2.9 mg/g, respectively. The Mg contents of cigar tobacco leaves treated with Mg fertilizer were higher than that of other treatments (Figure 1), and the leaf color was greener (Supplementary Figure S1).
Figure 1
Mg contents of cigar tobacco leaves during harvest, no fertilizer (CK), nitrogen - phosphate - potash fertilizers (NPK), and NPK fertilizers + 75 kg/hm2 MgO fertilizer (NKP+Mg). The values are means of four replicates ± standard error (SE); the different letters indicate there are significant differences between the treatments at p < 0.05 level according to Duncan’s multiple range test.
. The chroma values of cigar tobacco leaves during airing
The brightness value L* (Figure 2A), the yellow-blue value b* (Figure 2C), and the total color difference value ΔE (Figure 2D) showed a trend of first increasing (0–6 days), then decreasing (6–8 days, the color changed from yellow to reddish-brown, the brightness decreased, the red became deeper, and the yellow became lighter), and then stabilizing (>8 days), and all reached the maximum on the sixth day, with the average values of 61.68, 41.44, and 28.06 respectively. The red-green value a* gradually increased with the airing process and then became stable, and each treatment reached the maximum on the 10th day; the values of treatments were 4.56, 9.46, 9.15, respectively (Figure 2B, Supplementary Figure S2). After airing for eight days, the color gradually changed from reddish-brown to tan on the basis of the original reddish-brown, the brightness continued to decrease, the red was deepener, and the yellow was lighter (Supplementary Figure S2).
Figure 2
The leaf chromaticity of cigar tobacco leaves during the airing period. (A) luminance value L*; (B) red-green value a*; (C) yellow-blue value b*; (D) total color difference value ΔE. CK - red lines (no fertilizer), NPK - blue lines (NPK fertilizer), NPK+Mg - green lines (NPK+MgO fertilizer). Treatment is measured in airing days.
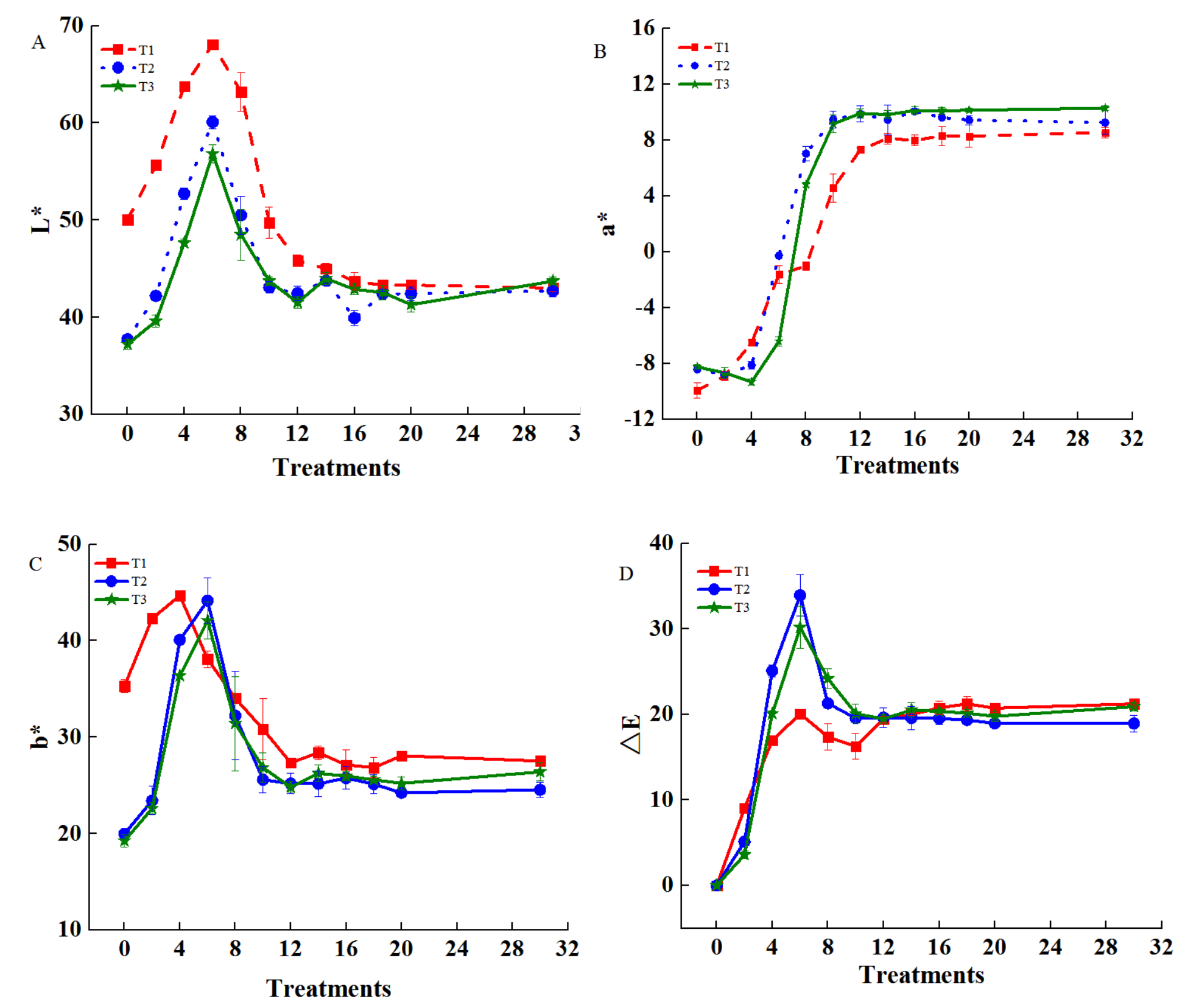
The total color difference ΔE could characterize the overall change in the color of tobacco leaves. When ΔE > 2, a significant change in color could be observed. As shown in Figure 2D, ΔE reached the maximum on the sixth day and remained stable after 10 days. These indicated that the color changes of cigar leaves in this study were mainly concentrated in the first 10 days (pre-airing). During this period, the L* and b* of the fertilizer treatment (NPK, NPK+Mg) were significantly lower than those of the no fertilizer (CK), and the a* and ΔE values were opposite to those of the CK, which indicated that fertilizer treatments would reduce the yellowness of the cigar tobacco leaves, which was consistent with the apparent color of the cigar tobacco leaves in airing (Supplementary Figure S2). The L*, a*, and b* values of NPK+Mg treatment were significantly lower than NPK treatment, and the L* and b* of CK were significantly higher than that of other treatments, but a* and E were significantly lower than that of fertilization treatments. Those results indicated that CK treatment could significantly improve the brightness and yellowness of tobacco leaves. Applying Mg fertilizer would reduce the brightness and redness of cigar tobacco leaves to a certain extent, and the color of tobacco leaves was brown, making the apparent color of cigar leaves darker (Supplementary Figure S2).
. The pigment contents of cigar tobacco leaves during the airing
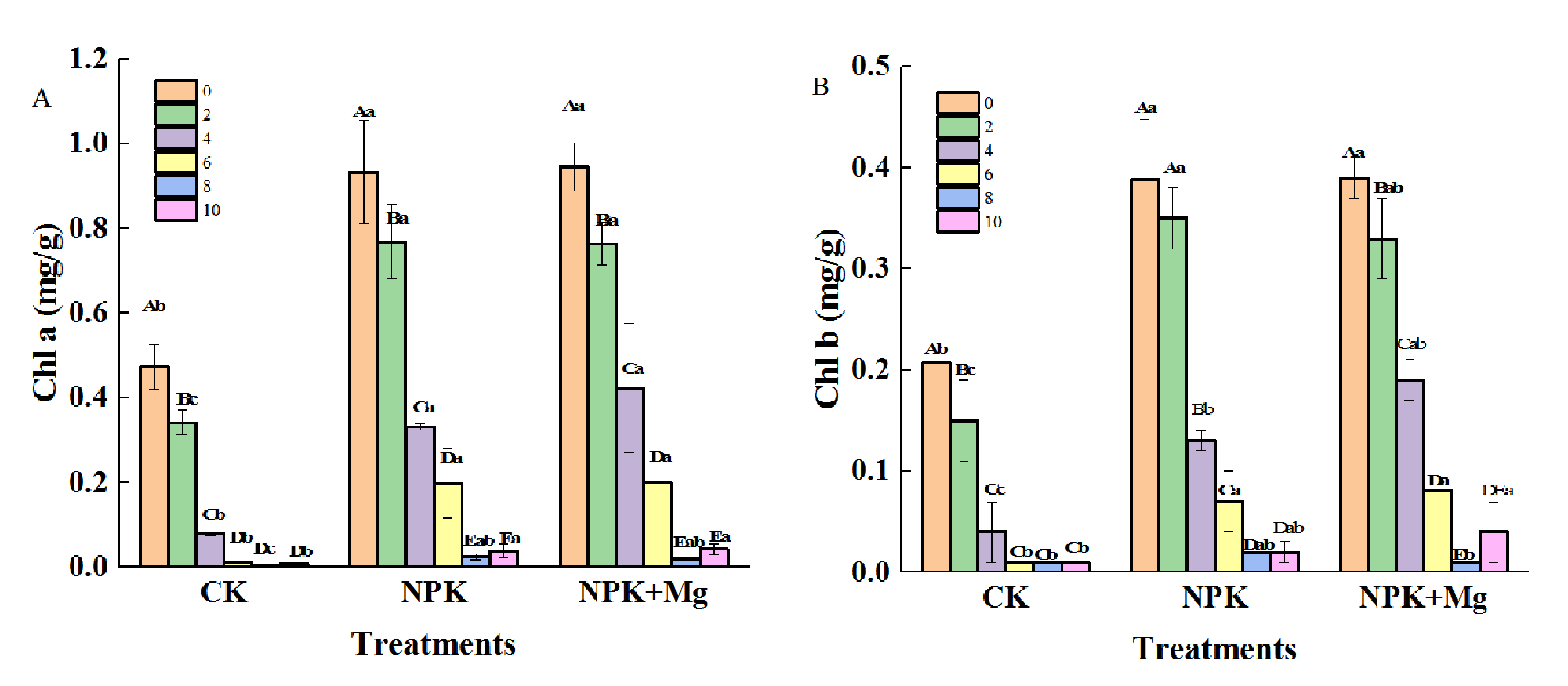
As shown in Figure 3, Chl a, Chl b, and TC contents in cigar tobacco leaves all showed a rapid decline with the advancement of the early airing process, and the content reached the lowest on the eighth day, with the average values of 0.015, 0.010 and 0.025 mg/g, respectively (Figure 3A,B,D). The Chl a, Chl b, and TC contents in NPK and NPK+Mg treatments were significantly higher than that in CK treatment, which indicated that applying fertilizer treatments could significantly increase the pigment contents of cigar tobacco leaves.
Figure 3
Pigment contents in cigar tobacco leaves during airing period. (A) Chl a (Chlorophyll a); (B) Chl b (Chlorophyll b); (C) Car (Carotenoid); (D) TC (Total Chlorophyll); (E) TC/Car (Total Chlorophyll/Carotenoid). Different capital letters on the bar chart represent the significant difference with the airing time, and different lowercase letters represent the significant difference between treatments at the same time according to Duncan’s multiple range test at the p < 0.05 level.
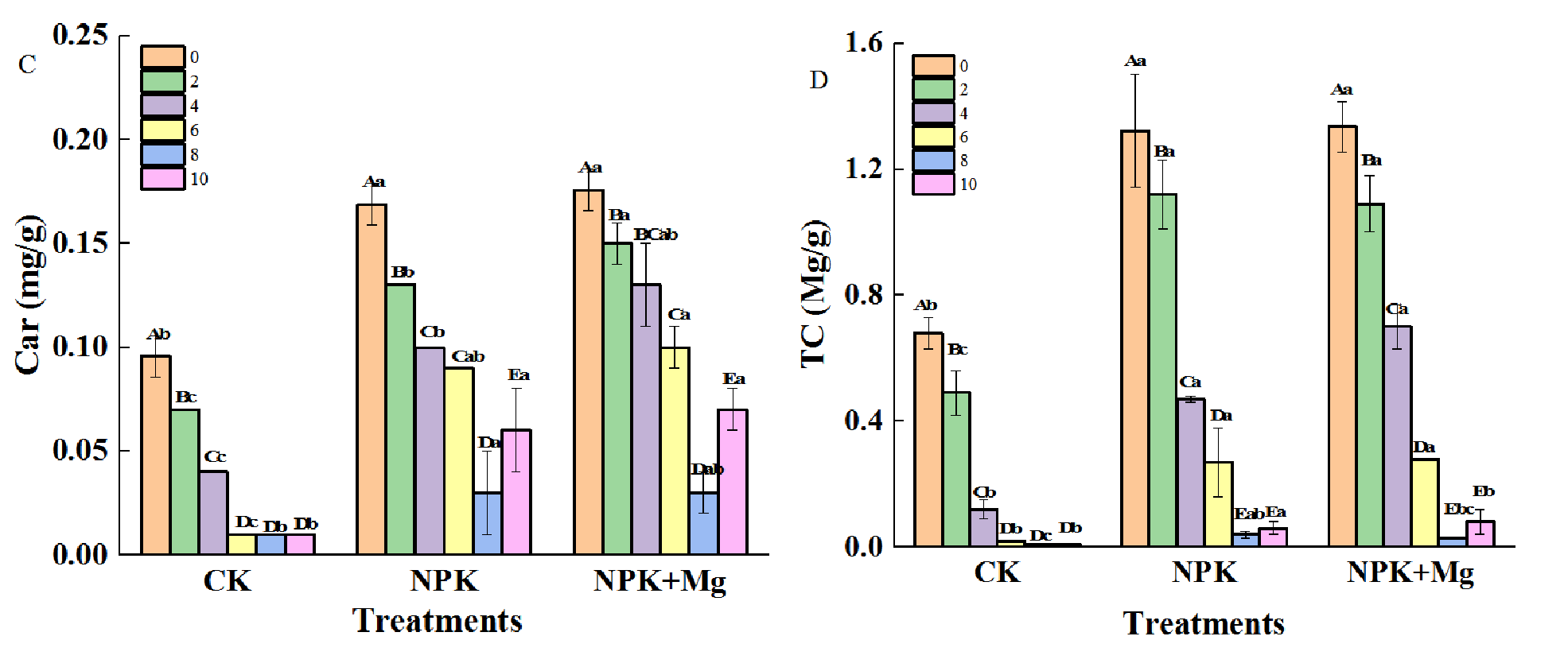
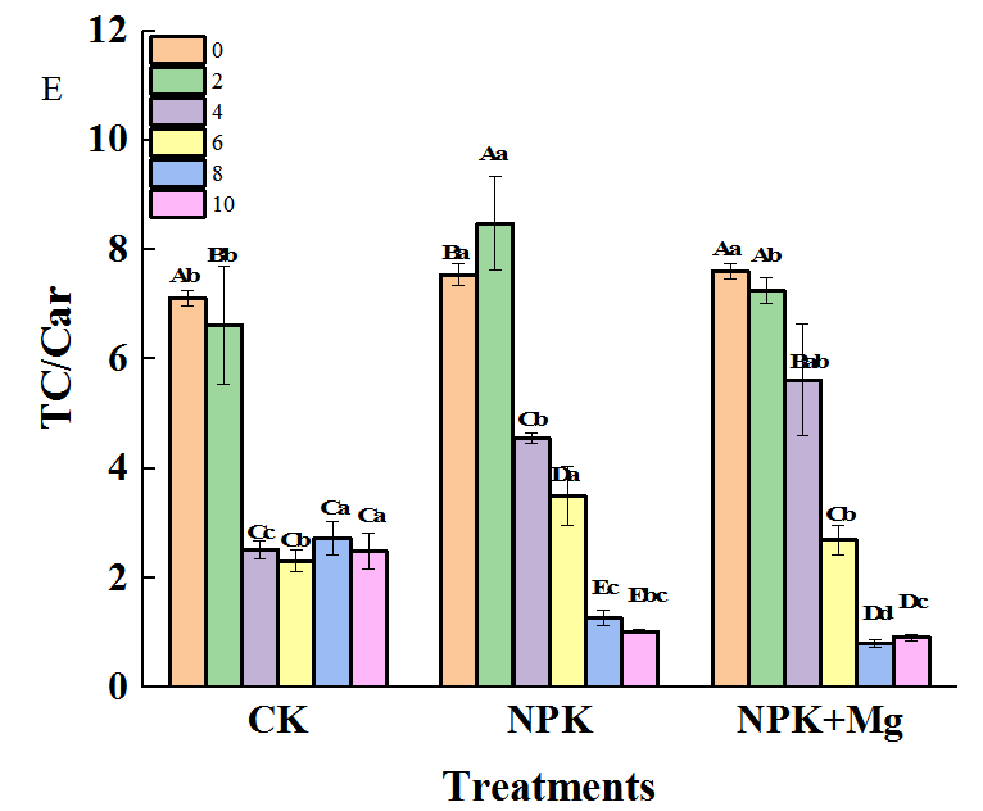
The TC/Car would directly affect the change of cigar tobacco leaf color. During the airing process, the changing trend of the TC/Car ratio of NPK treatments was different from that of the other two treatments. The NPK treatments showed a trend of first increasing and then decreasing and reached the maximum on the second day; at this time, the level of TC/Car in NPK treatment was significantly higher than that of other treatments (Figure 3E), indicating that the Chl degradation of the cigar tobacco leaves in NPK (without Mg fertilizer) treatment was insufficient in the first two days (withering stage).
However, the TC/Car in the Mg fertilizer treatment reached a significant level in 2–8 d, which indicated that applying Mg fertilizer would be beneficial to promote the degradation of Chl in cigar tobacco leaves in the early stage of airing, which in turn would be beneficial to the process of yellowing of tobacco leaves.
. The enzyme activities of cigar tobacco leaves during airing
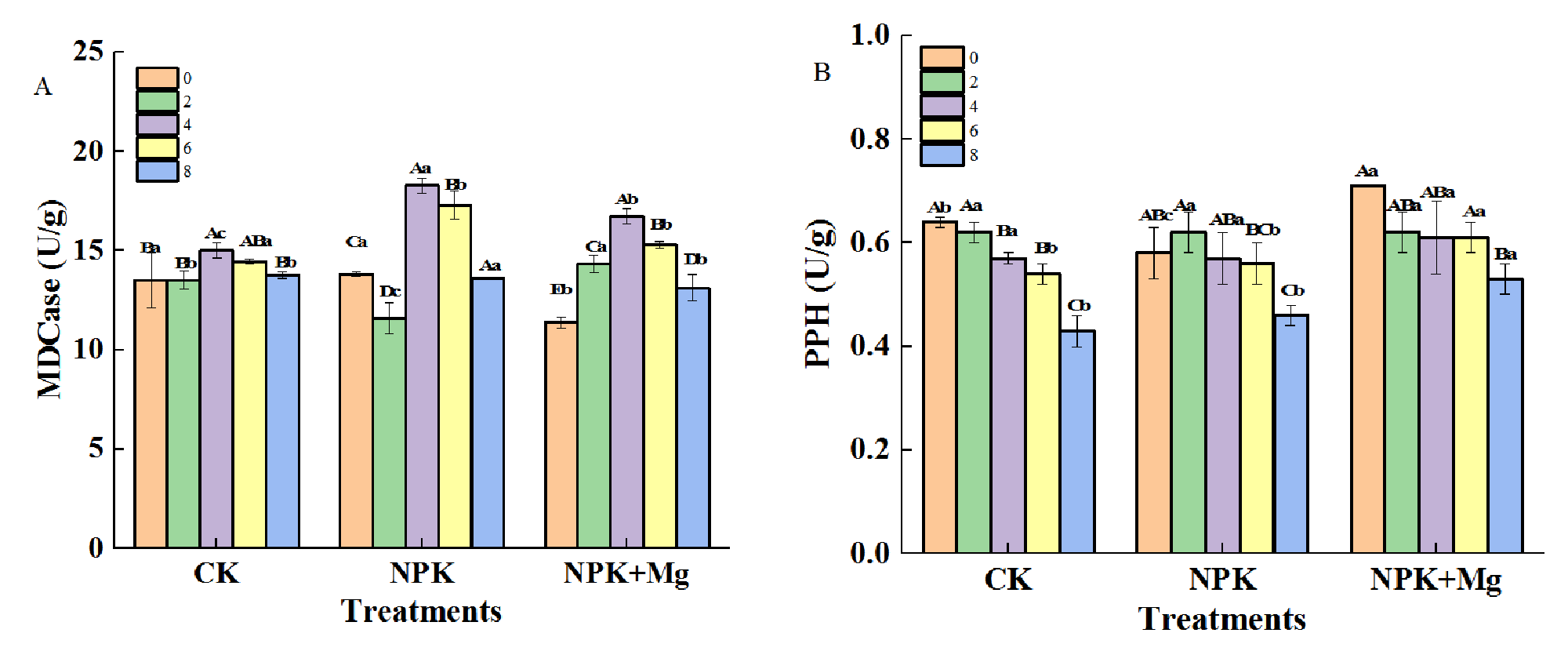
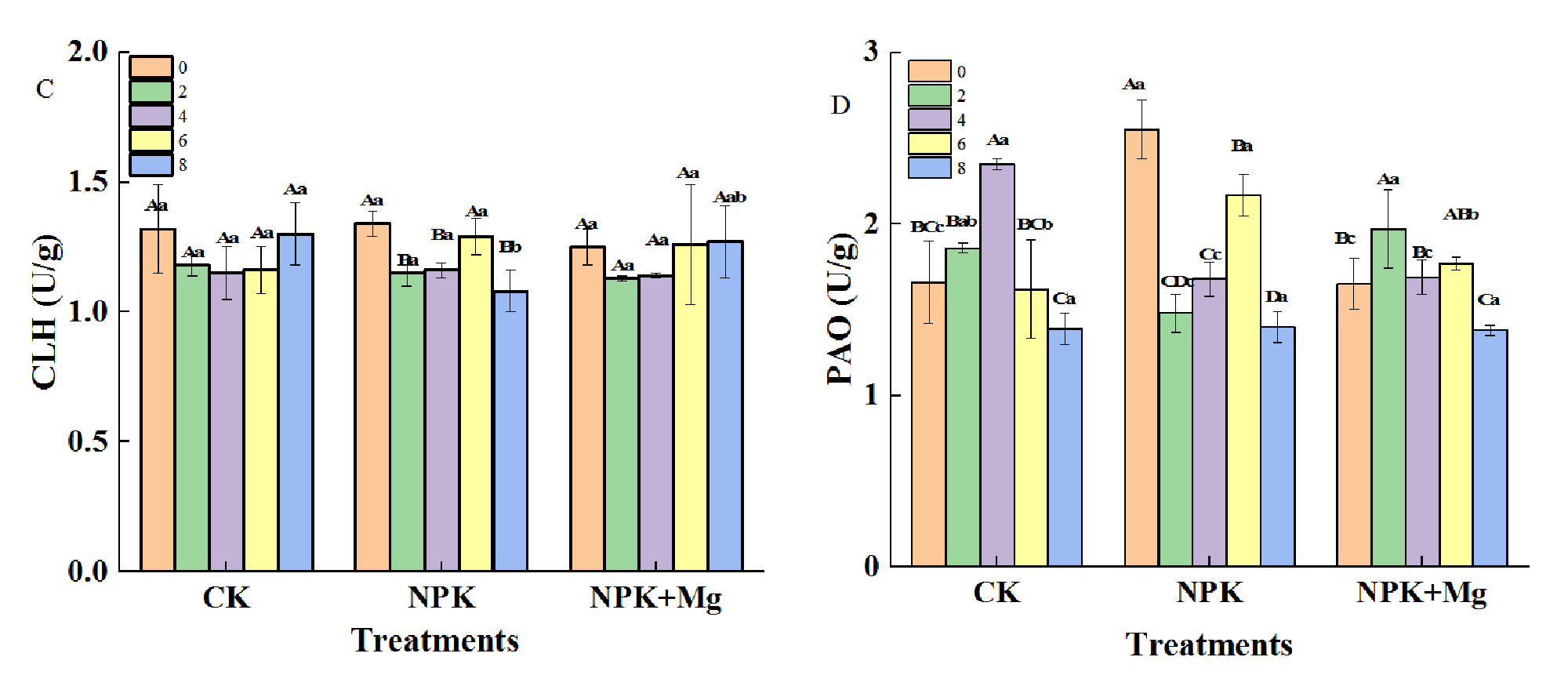
The activities of MDCase enzyme in CK and NPK+Mg treatments first increased and then decreased during airing, and the enzyme activities reached the highest value on the fourth day at 15.0 and 18.9 U/g, respectively (Figure 4A). On the contrary, CLH activities were not significantly different among different treatments (Figure 4C). The PPH activities showed a decreasing trend during the airing process, and the PPH activities of the NKP+Mg treatment were significantly higher than that of the other treatments on the 10th day (Figure 4B). The changing trend of PAO activities was different among treatments in the early stage of airing (Figure 4D). The CK and NKP+Mg treatments first increased and then decreased, while the NPK treatment was the opposite. The absence of Mg fertilizer might inhibit the activity of PAO and hinder the degradation of Chl by accelerating leaf water loss rates in the pre-airing stage (Supplementary Figure S3), and Mg mainly affected the PPH pathway of Chl degradation in cigar tobacco leaves to regulate the change of leaf color.
Figure 4
Enzyme activities in cigar tobacco leaves during airing period. Enzyme activities were measured for 0, 2, 4, 6 and 8 d for each treatment. (A) Mg-dechelatase (MDCase); (B) pheophytin pheophorbide hydrolase (PPH); (C) chlorophyllase (CLH); (D) pheophorbide a oxygenase (PAO). Different capital letters on the bar chart represent the significant difference with the airing time, and different lowercase letters represent the significant difference between treatments at the same time according to Duncan’s multiple range test at the p < 0.05 level.
. Chrominance correlation analysis
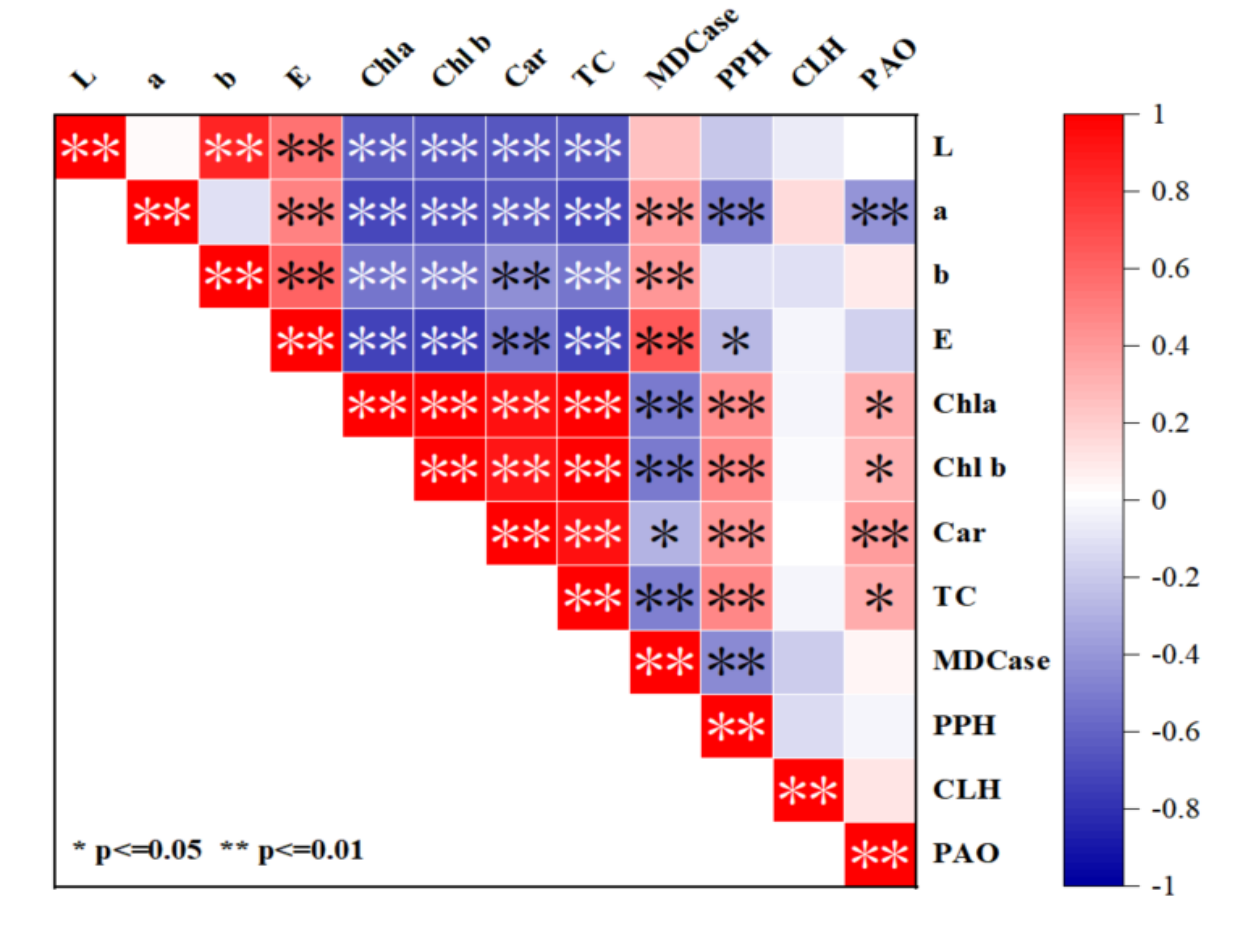
The chromaticity values (L*, a*, b*, ΔE) and pigments (Chl a, Chl b, TC and Car) were all extremely significantly negatively correlated. The red-green value a* was extremely significantly positively correlated with CLH but extremely significantly negatively correlated with PPH and PAO (Figure 5). The total color difference value ΔE was significantly positively correlated with MDCase and PAO, but extremely significantly negatively correlated with PPH. Chl a, Chl b, TC and Car were significantly positively correlated with PPH and PAO, but were not significantly correlated with CLH and MDCase. Further analysis showed that this process was accompanied by the degradation of TC and Car and then directly affected the change of leaf color.
Figure 5
Correlation analysis of cigar tobacco leaves color and internal chemical components during the airing process. Red represents positive correlation, blue represents negative correlation, and the darker the color, the greater the positive correlation. * represents significant correlation, ** represents extremely significant correlation.
. Discusion
The leaf color was largely affected by its pigment decomposition and transformation and specific physiological metabolism (León et al., 2007). This paper investigated the degradation of pigments (Chl a, Chl b, Car, TC) and changes of enzyme activities (MDCase, CLH, PPH, PAO) also compared the differences in the mechanism of color change under the conditions of Mg fertilizer application or not by elucidating the mechanism of the leaf color from green to yellow to brown in the pre-airing period of cigar CX26 tobacco leaves. According to the chromaticity values of this study, the discoloration of cigar tobacco leaves was mainly concentrated in the first 10 days. Further analysis of the correlation between pigment degradation and leaf color changes showed that the chromaticity values (L*, a*, b*, ΔE) were significantly negatively correlated with the content of pigments (Chl a, Chl b, TC, Car) in cigar tobacco leaves at the pre-airing stage. TC/Car gradually decreased in the early stage of airing, which might be due to the large degradation rate of Chl and the low degradation rate of Car in the pre-airing stage, which caused the change of the pigment ratio in the leaf tissue, resulting in the reduction of the relative content of TC. This led to a change in the leaf color of cigar tobacco leaves in the pre-airing stage, which was consistent with the research results of Wang et al. (2012) and Zhang et al. (2014). This indicated that the degradation of chlorophyll content during airing might be a key factor in the color change of tobacco leaves after conditioning (Lu et al., 2019). In this experiment, only applying NPK fertilizer increased TC/Car in the first two days, which may be because no Mg fertilizer reduced the activity of PAO, which would lead to the inhibition of the decomposition of chlorophyllin macrocycle, thus inhibiting the degradation of chlorophyll. The untimely and insufficient degradation of chlorophyll would affect the formation of leaf color and produce phototoxicity in plant cells. The untimely and insufficient Chl degradation would affect the formation of leaf color and cause phototoxicity to plant cells (Pruzinska et al., 2003). In NPK+Mg treatment, TC/Car showed a gradual downward trend, which indicated that applying Mg fertilizer would be beneficial to promote the degradation of Chl in cigar tobacco leaves in the pre-airing stage, which would be beneficial to the yellowing process of cigar tobacco leaves. Chl degradation under the action of SGR gene expression was the main cause of leaf color changes (Ono et al., 2019). At the same time, PPH expression contributed to the formation of grape berry color (Lu et al., 2022). Chl-degrading peroxidase (Chl-POD) and pheophorbidase (Pheidase) were the key enzymes to promote the decomposition of Chl during the airing process of prickly ash (Chen et al., 2012). In addition, rapid airing could inhibit the activity of enzymes and block the Chl degradation pathway (Chen et al., 2012; Pruzinska et al., 2005; Aiamla-or et al., 2010; Zou et al., 2012). Harpaz-Saad et al. (2007) believed that CLH deplantation reaction played a major role in the degradation of Chl in plants and crops such as zucchini and cigar, while PPH deplantation reaction in crops such as Arabidopsis (Schelbert et al., 2009), rice (Morita et al., 2009), tomato (Guyer et al., 2014). Chl content was significantly positively correlated with CLH (Zou et al., 2012). The results of this study showed that Chl content was not significantly correlated with CLH and MDCase, but was positively correlated with PPH and PAO. This study believed that the change of cigar tobacco leaves’ color was mainly regulated by Chl degradation of PPH.
The activity of MDCase was the highest on the sixth day of post-harvest storage of lychee fruit (Almela et al., 2000). The activity of MDCase was the highest on the ninth day, and its activity first increased and then decreased with the extension of refrigeration time (Zou et al., 2012). This is consistent with the trend of MDCase activity increasing first and then decreasing with the progress of the airing process of CX26 cigars in this study. However, the MDCase activity in this study was the highest on the fourth day, indicating MDCase may also play a role in the regulation of cigar tobacco leaves Chl degradation. Therefore, this study believed that MDCase first demagnesium, and then PPH deplantation were the main degradation pathways of Chl degradation in cigar CX26 tobacco leaves, which was different from the results of Harpaz-Saad et al. (2007). Wiwart et al. (2009) used color images to analyze the color response of leaves of different legumes to N, P, K, and Mg deficiency, which showed that the special sensitivity of plants to Mg deficiency caused substantial changes in pea leaf color. Spraying Mg fertilizer on lychee leaves can make the fruit turn red and solve the problem of evergreen (Gao et al., 2015). The brightness and total color difference of the tea leaves treated with Mg fertilizer were higher than those of the control treatment, and the tea yield was significantly improved (Farhat et al., 2015). The results of this study showed that the application of Mg fertilizer could reduce the brightness and redness of leaves to a certain extent. The color of cigar tobacco leaves was brown, and the apparent color of cigar tobacco leaves was dark and shiny.
. Conclusions
The application of Mg fertilizer reduced the brightness value L* and yellowness value b* of tobacco leaves and promoted the degradation of Chl in the yellowing and browning stages of cigar tobacco leaves. Mg mainly regulated the change of leaf color by affecting the Chl degrading PPH deplantation pathway of cigar CX26 tobacco leaves. In addition, applying Mg fertilizer would reduce the brightness and redness of leaves to a certain extent. The color of cigar tobacco leaves was brown, and the apparent color of cigar tobacco leaves was darker.
. Supplementary material
The following supplementary material is available for this article:
Figure S1. The color status of cigar tobacco leaves during harvesting.The tobacco leaves in the figure are the leaf state of CK, NPK and NPK+Mg fertilization treatment when they are ripe for harvest. The leaves of CK treatment are yellow, the leaves of NPK treatment are green, and the leaves of NPK+Mg treatment were dark green.
Figure S2. The color status of cigar tobacco leaves during airing.
Figure S3. Changes of moisture content of cigar tobacco leaves during. The water loss rates of CK, NPK, and NPK+Mg treatments were 2.57, 2.2, and 2.50, respectively.
