. Introduction
The invasion of alien species has seriously threatened the global biodiversity, and brought tremendous ecological and economic losses to the invaded regions (Ehrenfeld, 2010). The cost of eradication of invasive plants rises with the increase of invasion time, especially for those species that form persistent soil seed banks (Gioria & Pyšek, 2016). In the prevention and control plan of invasive plants, understanding the characteristics of the soil seed bank is crucial for the successful implementation of control measures (Cacho et al., 2006). The development of some seed persistence prediction technologies has provided some references for decision makers to make faster and wiser choices in weed management plans (Panetta et al., 2011). It is particularly important to evaluate the seed longevity of invasive plants when planning the orders of eradication or making budget allocation for eradication in some areas (Fenollosa et al., 2020). There are a variety of methods to evaluate seed longevity. Among them, the accelerated aging (AA) test is most commonly applied. In this test, seeds are exposed to elevated temperature (40–45 °C) and high relative air humidity (above water or salts solutions), leading to increase of water content in seeds and their rapid deterioration. This environment produces high relative humidity. The method was first proposed by Delouche and Baskin (1973). Currently, artificial aging methods to test the germination of seeds by exposure to controlled high temperature and humidity have been widely carried out in plant seeds such as food crops (Liu et al., 2008; Modarresi et al., 2002; Qin et al., 2010) and forage crops (Cookson et al., 2001; Mao et al., 2008; Y. H. Wang, Wang, et al., 2008). But the method has rarely been applied targeting invasive plants.
Aegilops tauschii, an annual grass species, is commonly known as “rough-spike hard grass.” The species originated in Eastern Europe and West Asia and is considered as one of the top 10 malignant weed species (X. Y. Wang, 2017). With a wide range of adaptation, the species is invasive in wheat fields, and is distributed from the arid Artemisia desert to the humid temperate forests (Fang, 2012; Miranda et al., 2007). Presently, this weed has invaded more than 10 major wheat producing provinces in China, including Hebei and Shandong. The damage caused by the species to wheat was surveyed by the Institute of Plant Protection of the Chinese Academy of Agricultural Sciences, in Hebei and Shanxi provinces. Results showed that a large-scale damage by A. tauschii to wheat had occurred, and the yield of wheat was reduced by more than 50% (Y. J. Wang et al., 2018). According to Fang (2012), A. tauschii seeds could survive for 3–5 years in the soil because of the protection by their tough glumes. Following the classification method of soil seed banks by Thompson and Grime (1979), A. tauschii should belong to the persistent soil seed bank group. However, so far, there has been no report on the seed longevity of A. tauschii. Therefore, considering the wide distribution, difficulty in eradication, and ecological impacts, the present study aims to evaluate the effects of various parameters related to seed germination. In this study, estimation of seed vigor on the seeds of A. tauschii in natural populations by rapid artificial aging tests was carried out. The results will serve as a reference for evaluating the invasion potential of A. tauschii and choosing the appropriate eradication strategies.
. Material and Methods
. Material
Seeds of A. tauschii were collected in May 2019 from the experimental field of Xinxiang Academy of Agricultural Sciences (35°18′ N, 113°52′ E), He’nan Province (Figure 1). After collection, the seeds were sun-dried and stored in a container in the laboratory. Seeds of Chinese cabbage (Brassica rapa L. var. glabra Regel), which were used in the determination of germination inhibitors, were purchased from Luoyang Seed Company. The experiment was carried out in the Garden Plant Laboratory of the College of Forestry, Henan University of Science and Technology, China, in September 2019. The 1,000-grain mass of A. tauschii seeds was 56.49 g and the initial relative water content (RWC) and germination rate (GR) were 7.54% and 75.33%, respectively.
. Methods
. Artificial Aging Test
The artificial aging treatment of A. tauschii seeds was conducted using the aging treatment temperature of 41–45 °C for a period of 48–144 hr, as suggested by the International Seed Testing Association (ISTA). A total of five groups of A. tauschii seeds that weighed about 35 g in each group were packed in separated gauze bags, labeled, and placed evenly in a sealed constant temperature and humidity chamber with preadjusted temperature of 40 °C and relative humidity of 95%. The aged seeds were sampled at Day 0 (control group; CK), 2, 4, 6, and 8. After indoor-drying, they were subjected to different tests to determine the related indicators.
. Seed Germinability and Vigor
Seed Vigor Index
The filter paper method was adopted, with 50 grains in each dish and three replications per group. The seeds were then cultured in an illuminated incubator with temperatures of 25 °C in the day and 15 °C at night, and 12 hr of daylight and 12 hr of darkness. The germination was recorded every day, considering the emerging radicle length greater than or equal to seed length as germination. After the experiment was completed, the seed GR, germination energy (GE), and germination index (GI) were calculated as follows:
where Dt is the corresponding number of days of germination, and Gt is the number of germinated seeds on the tth day after the start of germination. Higher values of GR indicate better seed quality, higher values of GE indicate higher germination speed and neatness of seeds, and higher values of GI indicate higher seed viability.
Seed Moisture Content
Appropriate quantities of seeds were taken on Day 0, 2, 4, 6, and 8 of aging. The high constant temperature oven method (Yao et al., 2015) was used to determine the water content of the seeds, using the following equation:
Higher values of RWC indicate higher seed metabolism.
. Biochemical Changes in Seeds
The glumes were removed, and approximately 2.50 g of embryo and endosperm of the seeds of each treatment group were extracted. To measure superoxide dismutase (SOD) activity, the nitro-blue tetrazolium photoreduction method was used according to H. S. Li (2000) and Zou (2003). The thiobarbituric acid method was used to measure the thiobarbituric acid (TBARS) content. The protocol followed for relative electric conductivity (REC) measurement was similar to the one described in Zou (2003) with slight improvement. The measurement of REC, SOD activity, and TBARS content were determined as described by N. Wang et al. (2021). Higher relative conductivity indicates greater cell membrane permeability, higher SOD activity indicates greater resistance to stress, and higher TBARS indicates higher cellular membrane lipid peroxidation.
In addition, appropriate quantities of seeds were taken on Day 0, 2, 4, 6, and 8 of aging, to obtain about 1.00 g of embryo and endosperm, which were then stored in an ultra-low temperature freezer at −70 °C. After all sampling was completed, the endogenous hormone levels were determined. The enzyme-linked immunosorbent assay (ELISA) (H. S. Li, 2000) was used to determine the levels of gibberellin (GA3) and abscisic acid (ABA), respectively, and then the GA3/ABA value was calculated. Higher gibberellin content indicates more favorability for seed germination, higher abscisic acid content indicates less favorability for seed germination and higher GA3/ABA ratio indicates more favorability for seed germination.
. The Inhibitory Effect of A. tauschii Extracts on Seed Germination of Chinese Cabbage
An amount of 2.50 g of the glumes, and 2.50 g of embryos and endosperm of the A. tauschii seeds of each treatment group were taken, ground by mortar, added with an appropriate amount of distilled water to transfer into Erlenmeyer flasks and extracted in a constant temperature incubator at 25 °C for 24 hr. The residue, left by filtering was re-extracted twice. Following this, the extract volume was adjusted to 25 mL and stored in a refrigerator at 4 °C for future use.
Seeds of Chinese cabbage are used as a test material for germination inhibitors because they have a high germination rate and are very easy to germinate. The Chinese cabbage seeds were soaked in 1% sodium hypochlorite solution for 10 min for disinfection, followed by rinsing with distilled water repeatedly, and then air-dried indoor. The seeds were placed in a Petri dish with a diameter of 9 cm covered with double-layer filter paper. Each Petri dish contained 100 grains and was added with an appropriate amount of glume extract, or embryo and endosperm extract, or the same amount of distilled water (CK). Each treatment was repeated three times. After incubating in a constant temperature incubator at 25 °C for 48 hr, the germination rate of each treatment was calculated, considering the radicle length as being half of the seed length as germination. In addition, 15 seedlings from five plants per dish and three repetitions per treatment were randomly selected to measure the shoot length (mm) and root length (mm).
. Data Analysis
The SPSS 18.0 software was used to carry out the statistical analysis. One-way analysis of variance (ANOVA) was used to compare the differences in the same index after different mesosulfuron-methyl treatments. The significance was tested using the least significant differences (LSD) at p = 0.05. Statistical values are expressed as mean (±SE). Microsoft Excel was used to summarize the data and plot figures.
. Results
As the aging time extended, the RWC of A. tauschii seeds kept increasing, while the seed GR, GE, and GI kept decreasing (Table 1). Starting from the second day, the RWC was significantly different from that in CK (p < 0.05). In addition, the GR, GE, and GI decreased significantly (p < 0.05) by 73.45%, 71.42%, and 67.61%, respectively, compared with that in CK on the second day of aging. The above results demonstrated that the increasing water content in the seeds during aging may accelerate their internal metabolism and cause the decline of the A. tauschii seed vigor.
Table 1
Effects of artificial aging on seed germination and relative water content of Aegilops tauschii seeds.
Figure 2
Effects of artificial aging on relative electric conductivity, thiobarbituric acid (TBARS) content, and superoxide dismutase (SOD) activity in Aegilops tauschii seeds. The values are shown as mean ±SE. Different letters above the bars denote significant differences (p ≤ 0.05) with Duncan’s test within the same planting conditions.
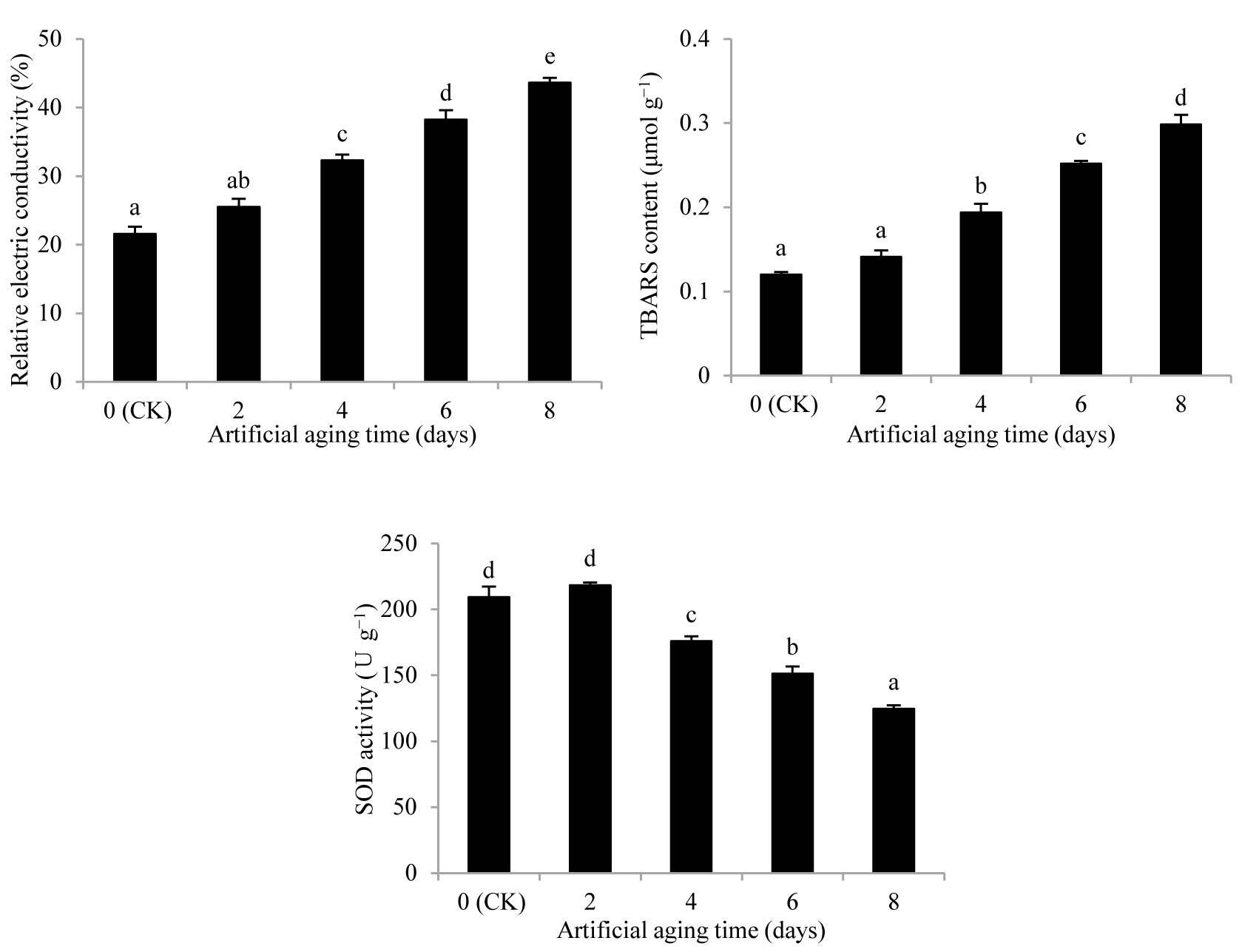
During the aging process, the REC and TBARS contents were both increasing (Figure 2). Further, the contents of REC and TBARS were significantly different from the values in CK on the fourth day of aging (p < 0.05), and significantly increased (p < 0.05) by 102.13% and 148.62%, respectively, compared with those in CK on the eighth day.
With the extension of the aging time, the SOD activity in the A. tauschii seeds first increased and then decreased (Figure 2). The SOD activity peaked on the second day, though not significantly differently from that in CK. It then declined by 43.22% compared with the value in CK on the eighth day and the value was significantly different (p < 0.05).
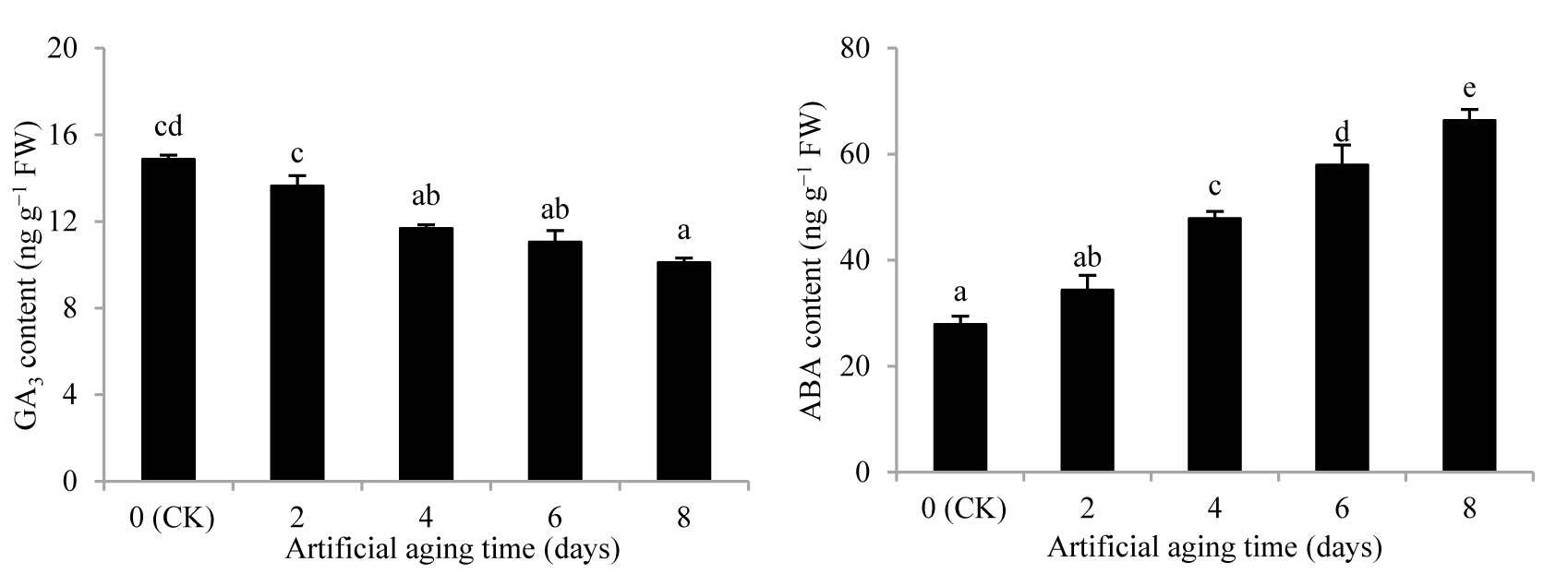
With the extension of the aging time, the GA3 content of A. tauschii seeds kept decreasing, whereas the ABA content was increasing (Figure 3). In addition, during the aging process, the GA3 and ABA contents were significantly different from those in CK on the fourth day (p < 0.05). By the eighth day, compared with the values in CK, the GA3 content decreased by 32.07% and the ABA content increased by 138.02% and both the differences were significant (p < 0.05).
Figure 3
Effects of artificial aging on endogenous gibberellin (GA3) and abscisic acid (ABA) content of Aegilops tauschii seeds. The values are shown as mean ±SE. Different letters above the bars denote significant differences (p ≤ 0.05) with Duncan’s test within the same planting conditions.
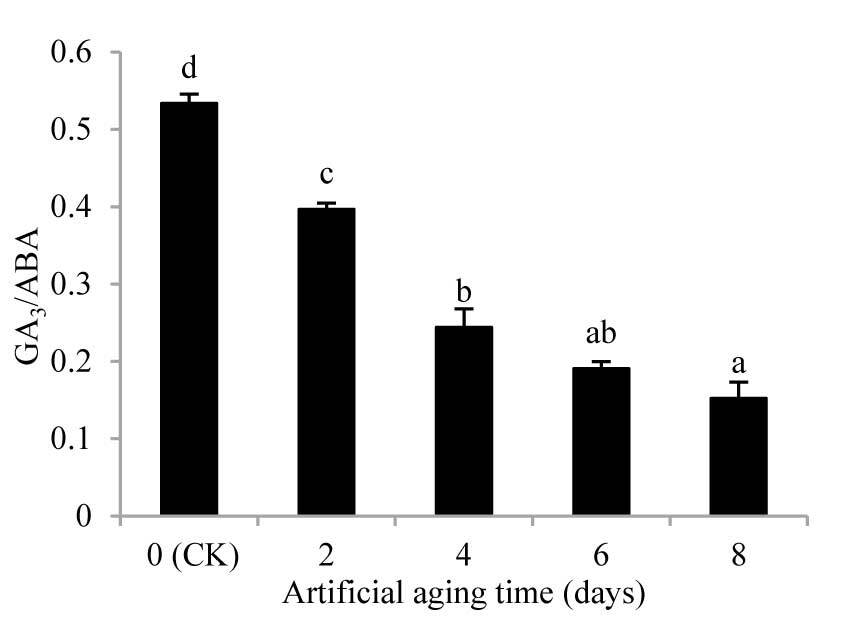
During the aging process, the GA3/ABA ratio exhibited a declining trend (Figure 4). From the second day onwards, the GA3/ABA ratio was significantly different from that in CK (p < 0.05). By the eighth day, the GA3/ABA ratio was significantly lower by 71.46% compared to that in CK.
Figure 4
The proportion of endogenous hormone contents of Aegilops tauschii seed in artificial aging. The values are shown as mean ±SE. Different letters above the bars denote significant differences (p ≤ 0.05) with Duncan’s test within the same planting conditions.
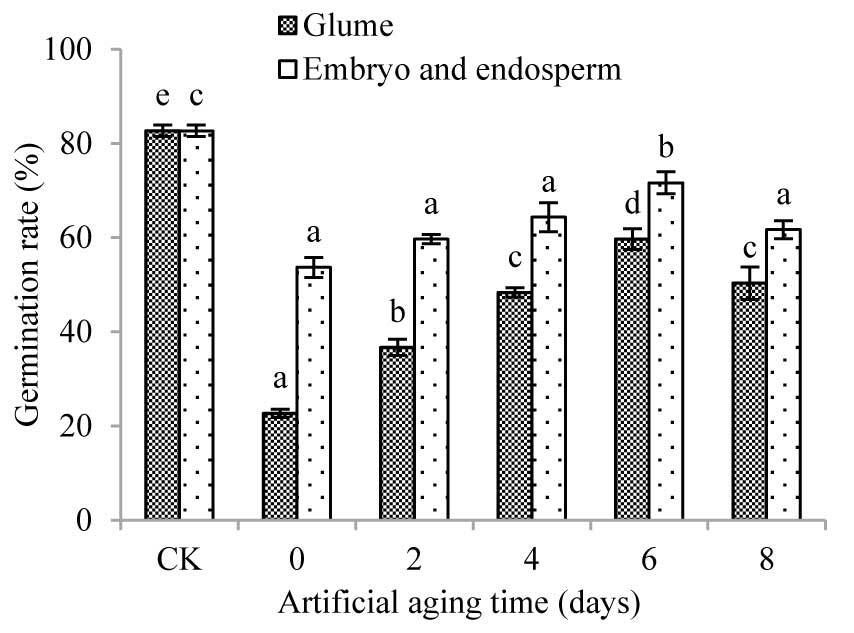
As the aging time increased, the germination rate of Chinese cabbage seeds treated with extracts from various parts of A. tauschii both increased initially and then decreased (Figure 5), indicating a concave shaped trend of the germination inhibitors in the glumes, embryos, and endosperms during aging. Regarding the treatment of glume extract, the GR of cabbage seeds on the second day of aging was significantly different from that on Day 0 (p < 0.05). Further, the GR on the eighth day of aging had increased by 122.01% compared with that on Day 0, but was still significantly lower than that in CK (p < 0.05). Regarding the treatment of embryo and endosperm extract, only the GR of cabbage seeds on the sixth day of aging was significantly different from that on Day 0 (p < 0.05), and it was still significantly lower than that in CK (p < 0.05). Since Day 0 of aging, the GR of cabbage seeds treated with glume extract was significantly lower than that of embryo and endosperm extract, indicating higher levels of germination inhibitors in the glume than in the embryo and endosperm.
Figure 5
Effect of water extract from glume, embryo, and endosperm of Aegilops tauschii on the germination percentage of cabbage seeds. The values are shown as mean ±SE. Different letters above the bars denote significant differences (p ≤ 0.05) with Duncan’s test within the same planting conditions.
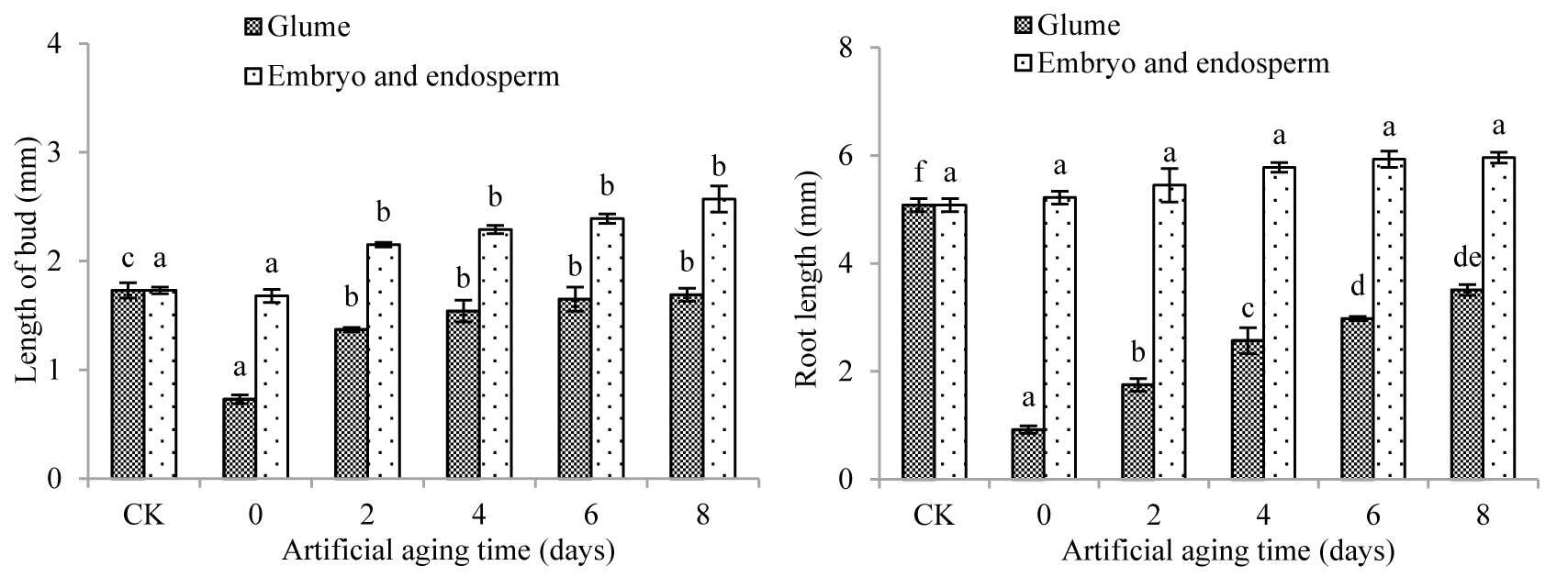
With the extension of aging time, the root length and shoot length of cabbage seedlings treated with extracts of various parts of A. tauschii seeds had both increased (Figure 6). For cabbage seedlings treated with the glume extract, the shoot length and root length on the second day of aging were significantly different from those on Day 0 (p < 0.05). By the eighth day, both the shoot length and the root length had significantly increased compared with Day 0 (p < 0.05), and both were significantly lower than that in CK (p < 0.05). For cabbage seedlings, treated with embryo and endosperm extract, the shoot length on the second day of aging was significantly longer than that on Day 0 (p < 0.05) and also compared to that in CK (p < 0.05). No significant differences were seen between the root length processed with various aging time, or from CK. In addition, from Day 0 of aging, the root and shoot lengths of cabbage seedlings treated with glume extract were significantly lower than those treated with embryo and endosperm extract. This further indicates that the germination inhibitors of A. tauschii seeds were more concentrated in glumes.
. Discussion
. Weed Control and Artificial Aging of Seeds
Company et al. (2019) conducted an aging study on the seeds of the invasive plant Cortaderia selloana and found that long-term monitoring was not needed following eradication because of its shorter seed longevity. Fenollosa et al. (2020) revealed that the two populations of the invasive plant edible dayflower Carpobrotus edulis (L.) L. Bolus showed different seed longevity after accelerated aging, which may increase the difficulty of eradication. In addition, artificial aging test can be used to determine the heat sensitivity of invasive plant seeds, providing ideas for effective eradication of soil seed banks. Soil solarization is a nonchemical and environmentally friendly agricultural method that is used to destroy weed seeds in the soil. Existing studies have shown that it was relatively easy to get a high temperature of 60 °C under polyethylene sheets with an air temperature of 30 °C from tropical forests to deserts (Cohen et al., 2019). Based on this concept, Fenollosa et al. (2020) combined the heat sensitivity of the seeds of C. edulis and the seed distribution characteristics in the soil seed banks and concluded that soil solarization was one of the effective methods to eradicate the seeds of this species.
. Effect of Artificial Aging on Seed Vigor
The relationship between seed moisture content and environmental temperature is the primary factor affecting seed life. Compared with Saccharum ravennae L., the invasive C. selloana showed a stronger water absorption capacity of the seeds in the aging process, leading to a significant decline in its seed vigor (Company et al., 2019; Mira et al., 2015). In this study, the RWC of A. tauschii seeds increased to a significant level compared with that in CK on the second day of aging process (p < 0.05) and it indicated that the continuous increase in water content promoted the metabolism of seeds in the aging, resulting in the decline of seed vigor.
Compared with natural aging, artificial aging greatly shortens the required time of aging on the A. tauschii seeds. On the second day of aging process, the GR, GE, and GI all decreased significantly, indicating a significant decrease in seed vigor due to the artificial aging treatment. The results were consistent with the findings of studies on the aging of seeds such as wheat Triticum aestivum L. (Qin et al., 2010; Y. J. Wang et al., 2018), maize Zea mays L. (Liu et al., 2008), and Psathyrostachys juncea Bozoisky (Y.-R. Li et al., 2005).
. Effect of Artificial Aging on the Physiological Characteristics of A. tauschii Seeds
Plasma membrane damage is one of the important physiological changes that occurs during seed aging, and the increased electrolyte and metabolite penetration are the main manifestations (Yao et al., 2015). The measurement of electrical conductivity provides an indirect approach to assess the loss of plasma membrane integrity (Delouche & Baskin, 1973); hence it is widely applied in artificial seed aging studies (Liu et al., 2008; Qin et al., 2010; Y. J. Wang et al., 2018). Furthermore, the decline of seed vigor in the aging process is also related to the loss of membrane integrity due to lipid peroxidation products. As the end product of lipid peroxidation, the content of TBARS becomes an important indicator of membrane damage (Niu et al., 2018). Some studies have revealed that the increase in electrical conductivity was negatively correlated to the seed vigor during aging (Mao et al., 2008; Perez & Arguello, 1995; Z. Wang, Na, et al., 2008), but other studies have showed no correlation between the two (Cookson et al., 2001; Liao et al., 2015; Y. R. Wang et al., 2002; Zhou & Wang, 2011). Our study demonstrated that both the relative conductivity and the TBARS content showed increasing trends during aging, but no significant differences from CK on the second day were observed. Presumably, it was related to the resolved damage to the membrane due to a short-term self-recovery of the A. tauschii seeds at the initial stage of aging. With the extension of aging, the intensification of lipid peroxidation further led to the increase of electrolyte extravasation.
As an important part of the enzymatic antioxidant system, SOD avoids damage to seeds by removing toxic substances produced by active oxygen (Ma et al., 2017). This result is consistent with the research conclusions on the species Elymus sibiricus L. (Fu et al., 2014) and Setaria sphacelata (Ma et al., 2017).
Seed germination is closely related to the levels of endogenous hormones. Various hormones can induce the decomposition and synthesis of internal substances and ultimately lead to the germination of seeds (Ogawa et al., 2003). Endogenous GA3 accelerates the decomposition and synthesis of internal substances in seeds by increasing the activity of various enzymes, thereby promoting seed germination (Brady & McCourt, 2003). ABA can antagonize the effect of GA3. It inhibits the normal metabolism of nucleic acid, interferes with the synthesis of ribonucleic acid, impedes the normal metabolic activities of seeds, and ultimately stops the normal gemination of seeds (Kucera et al., 2005; Rohde & Bhalerao, 2007). Furthermore, compared with a single endogenous hormone, the balance of hormones plays a more significant role in seed germination, especially the ratio of growth-promoting to growth-inhibiting hormones (Duan et al., 2011; Su et al., 2018). This study found that the artificial aging treatment resulted in a decrease in growth-promoting hormone GA3 in seeds, and at the same time resulted in an increase in growth-inhibiting endogenous hormone ABA. Together they contributed to a fast drop in the ratio of GA3/ABA, providing an unhealthy internal environment for seed germination.
. Effect of Artificial Aging on Seed Germination Inhibitors
Germination inhibitors exist in all parts of the seed (Hampton, 1995). From Day 0 of aging, the germination rate, shoot length, and root length of Chinese cabbage seeds treated with the glume extract were significantly lower than those treated with the embryo and endosperm extract. This result indicates that the germination inhibitors of A. tauschii seeds were more concentrated in the glumes. It is also consistent with our previous research on the causes of dormancy of A. tauschii seeds that the existence of germination inhibitors in the glume and its mechanical hindrance were important factors for dormancy (N. Wang et al., 2020). In addition, combining the observations that the germination rate of cabbage seeds treated with the glume extract showed a convex shaped alteration, and that the shoot length and root length increased fast first and then slowed down later, it could be seen that the variation of germination inhibitors exhibited a convex pattern. This indicated that an appropriate aging treatment could reduce the content of germination inhibitors in seeds and thus was conducive to seed germination, but a long-term aging treatment did not favor seed germination.
. Conclusion
During the aging process, the seed SOD activity increased slightly in a short period of time, and the germination inhibitors decreased marginally. Nevertheless, as aging went on, the aging treatment caused significant increases in seed RWC, REC, and TBARS, resulting in a decrease in GA3 and an increase in ABA, and thus a significant decrease in the GA3/ABA ratio. The changes in these internal factors eventually caused a rapid decline in the seed vigor of A. tauschii. Finally, according to the research results of artificial aging, considering the characteristics that the A. tauschii seeds fall off naturally after maturation and are mostly concentrated in the top soil, as well as the fact that the highest temperature in summer in Henan can reach 40 °C, soil solarization can be expected to be an effective method to eradicate the seed bank of this species.
