. Introduction
Worldwide, more than 1500 species belonging to the genus Entoloma (Fr.) P. Kumm. are known (Co-David et al., 2009; Kirk et al., 2008). However, it is presumed that the global diversity of these fungi is significantly greater, exceeding 2500 species (Noordeloos et al., 2022). In Europe, this genus is represented by more than 350 species (e.g., Noordeloos, 1992, 2004; Noordeloos et al., 2018; Noordeloos et al., 2022), while in Poland it includes at least 160 species (Kujawa, 2024; unpublished materials; Wojewoda, 2003).
Entoloma fungi produce cap-shaped, gymnocarpous fruiting bodies, equipped with a lamellar type of hymenophore. One of the basic distinguishing features of these fungi is the presence of pink or pinkish brown (in print) basidiospores with an angular or angularly tuberculate outline (Singer, 1986). Most Entoloma species are saprotrophic, but some form biotrophic relationships with Sphagnum mosses or engage in mycorrhizal associations with higher plants (e.g., members of the families Betulaceae, Pinaceae, Salicaceae, and Rosaceae) or parasitize the fruiting bodies of other fungi (Largent, 1994; Noordeloos, 1992).
Entoloma conferendum (Britzelm.) Noordel. (syn. Entoloma staurosporum (Bres.) E. Horak) is one of the easiest to identify and the most frequently recorded Entoloma species in Poland. This fungus is documented in the literature from at least 132 sites located in the northern, southern, eastern, western, and central parts of the country (Adamczyk, 2011; Adamczyk & Kucharski, 2005; Bujakiewicz, 1979, 2004, 2018; Bujakiewicz & Kujawa, 2010; Bujakiewicz & Lisiewska, 1983; Domański et al., 1960, 1963, 1967; Domański, 1965, 1999; Flisińska, 2004; Frejlak, 1973; Friedrich & Orzechowska, 2002; Gierczyk et al., 2009; Gierczyk et al., 2018; Grzesiak et al., 2017; Gumińska, 1972; Kałucka, 2009; Karasiński et al., 2015; Kryza & Puciata, 2009; Kujawa & Gierczyk, 2016; Kujawa et al., 2023; refer to Figure 1; Kujawa et al., 2019; Lisiewska, 1978, 1979, 2004; Lisiewska & Nowicka, 1979; Ławrynowicz, 1973; Łuszczyński, 2008; Łuszczyński et al., 2022; Nespiak, 1960, 1962; Pietras et al., 2016; Ronikier, 2012; Rudnicka-Jezierska, 1969; Stasińska, 2011; Stasińska & Sotek, 2017; Szkodzik, 2005; Ślusarczyk, 2007, 2019; Wojewoda, 1974, 2003; Wojewoda et al., 2004; Wojewoda et al., 2016). In the Sudetes, E. conferendum has so far been recorded from the western part of this mountain range, i.e. the Karkonosze Mts, Izera Mts, Kaczawskie Mts, Bardzkie Mts, the Upper Nysa Depression, and the Śnieżnik Massif (Mt. Śnieżnik) (Gierczyk et al., 2018; Kujawa et al., 2023; Schröter, 1885–1889). The main goal of this study is to present new records of this species found by the author in 2011 in the Śnieżnik Massif (near Biała Woda), provide a morphological characterisation, and summarise the knowledge on the ecology and distribution of E. conferendum in Poland. Achieving the main objective of the paper also required targeted studies on the contemporary interpretation of taxonomic nomenclature used in older mycological literature, particularly from the 19th and 20th centuries, with special attention to selected data presented in Schröter’s monumental work “Die Pilze Schlesiens”.

. Material and methods
Fieldwork was conducted in September 2011. Mycological data was collected using a transect method (cf. Jongman et al., 1995), with the aid of a GPS device and dedicated database software. Field photographs of fresh basidiomata were taken with the aid of a Pentax K10D camera equipped with a F2.8 Tamron SP 90mm macro lens. The basidiomata were collected, transported to the laboratory, and dried for further study. Characteristics that change over time (colour of basidiomata, smell, texture) were noted in the field.
Taxonomic analysis of the collected material was carried out using classical methods of mycological taxonomy, utilising a light microscope and specific chemical reagents and stains (Clémençon, 2009; Noordeloos, 1992). The macroscopic description is based both on the study of fresh material and on the analysis of photos. All microscopic structures were observed from dried material with a Nikon Eclipse E–400 light microscope equipped with a Nikon digital camera (DS-Fi1). Freehand sections of rehydrated basidiomata pieces were examined in squash preparations in 5% NH₃•H₂O and 1% aqueous solution of Fuchsin B in 5% NH₃•H₂O. Image capture and biometric analyses were performed with NIS-Elements D 3.1 imaging software. The dimensions of the microcharacters are given as follows: (minimum) 10th–90th percentile values (maximum), mean ± standard deviation. For basidiospores, randomly selected mature spores were measured without the hilar appendix. The length of the basidia was measured excluding sterigmata. Statistical computations were carried out using Statistica software (StatSoft). The following abbreviations are used: L = number of lamellae reaching the stipe, l = number of lamellulae between each pair of lamellae, Q = the length-width ratio of basidiospores (mean value). The number of analyzed elements is indicated in parentheses (n =), sequentially providing the number of measurements taken, the number of specimens analyzed, and the number of collections examined. The fungal nomenclature is united according to the Index Fungorum database (www.indexfungorum.org), while the plant nomenclature is based on Mirek et al. (2002) and Ochyra et al. (2003). Syntaxon names of plant communities are given according to Matuszkiewicz (2001, 2007). The collections studied are deposited at the Museum of Natural History of Wrocław University, Poland (WRSL).
In compiling data on the ecology, habitat requirements, and national distribution patterns of the studied species, a variety of information sources were utilized. This included data from available literature (cf. Kujawa, 2024; Wojewoda, 2003), a revision of materials deposited in the WRSL-F herbarium, as well as additional data and spatial information layers of a natural-geographical nature (e.g., resources of the Forest Data Bank and the geoportal website www.geoportal.gov.pl). Spatial data management and analysis were performed using the QGIS geoinformation software. A list of national localities of E. conferendum included in the study is provided in Appendix A.
. Results and discussion
Distinguishing features and morphological characteristics
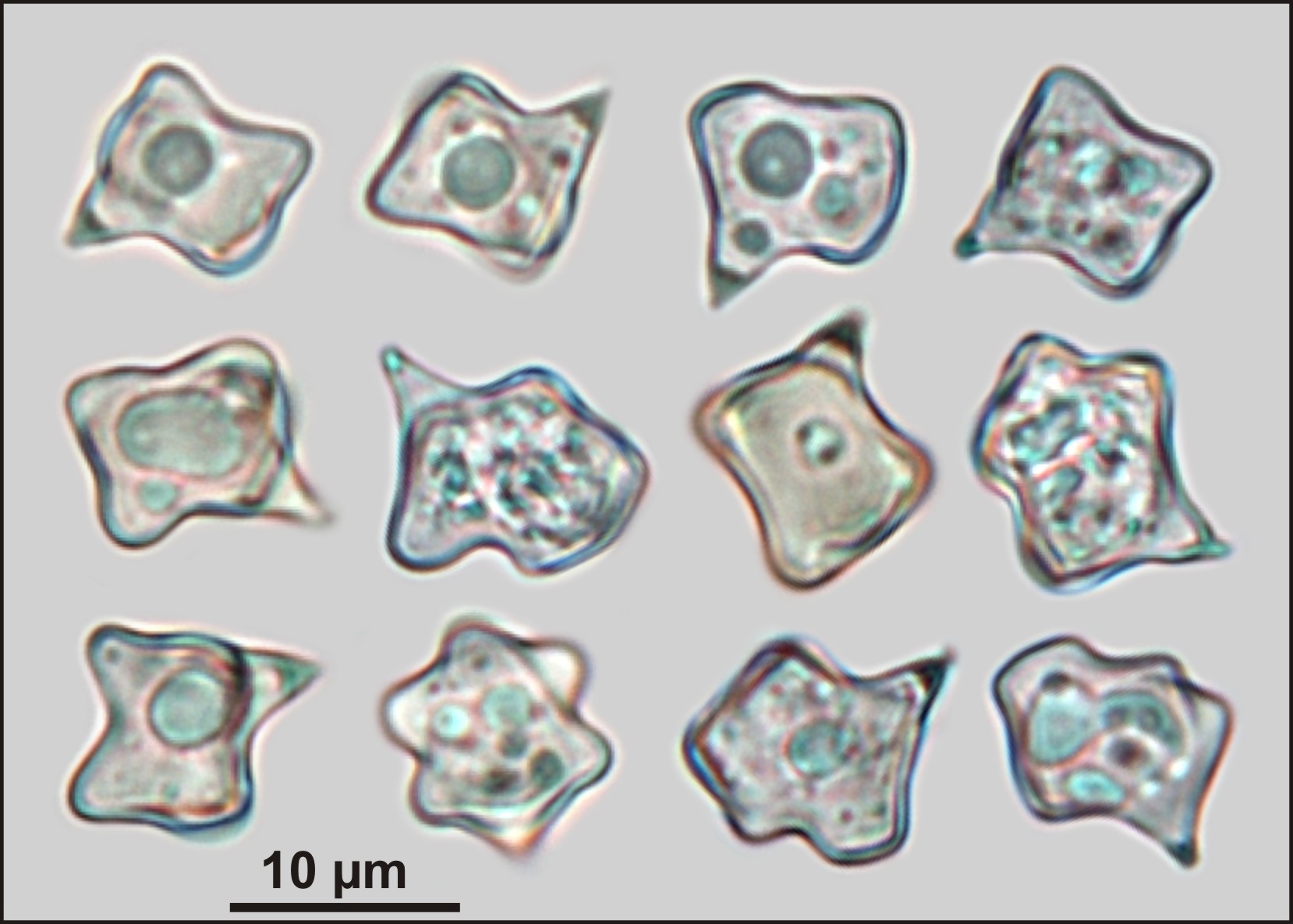
Although E. conferendum exhibits considerable morphological variability, it is distinguished by its clearly hygrophanous, shiny, grey brown cap, which, upon drying, often appears wrinkled in the central part and is covered with short fibrils. Furthermore, it is characterised by its silvery, fibrously striated, typically twisted stipe, and, most notably, its characteristic basidiospores. These spores, though diverse in shape, are predominantly cross-star-shaped in outline and feature 4 (rarely 5 or 6) corners when viewed in profile. Additionally, the species lacks clamp connections (Noordeloos et al., 2022). The morphological characteristics of the basidiocarps of E. conferendum found in new localities within the Śnieżnik Massif align with the contemporary concept of this species and descriptions available in the literature (e.g., Ćetković et al., 2020; Kaygusuz & Battistin, 2020; Noordeloos, 1980, 1992, 2004).
The colouration and general habit of the fruiting bodies of E. conferendum are illustrated in the accompanying photographs (Figures 1-3). Context pale brown grey or yellow grey in cortex of pileus and stipe, whitish or greyish in inner parts. Smell weak farinaceous. Taste not verified. The most important features of the micromorphological structure are as follows:
Basidiospores: (8.4) 9–11.9 (13), 10.4±1.1 × (7) 7.9–10.6 (11.6), 9.2±1.1 µm, Qw = (1) 1–1.3 (1.4), 1.1±0.1 (n=60/5/3), varied in outline and complex in shape, predominantly cross-star-shaped with 4 corners (less frequently 5 or 6); thin-walled and colourless in ammonia water (Figure 2). Basidia: (24.7) 25.7–31.5 (32.1), 28.8±2.4 × (10) 10.3–12 (13.2), 11.1±0.9 µm (n=13/5/3), 4-sterigmate, from ventricose-cylindrical to clavate. Cystidia: hymenial cystidia were not observed. Pileipellis: of the “cutis” type, transitioning to a “trichoderm” structure in the central region of the cap; it consists of an upper layer of cylindrical hyphae, 6.2–14 µm wide, with an intracellular beige-brown pigment (rarely also incrustatory-parietal) and a subpellis layer composed of inflated hyphae measuring 40–100 × 20–35 µm. Clamps not observed.
Examined collections: Poland, Śnieżnik Massif: 1. extensively managed mountain meadow, in humus composed of herbaceous and moss remains, near Achillea millefolium, Deschampsia caespitosa, Festuca sp., and Rhytidiadelphus squarrosus, 855 m a.s.l., 50°15'60"N, 16°47'53"E, 16.09.2011, leg. M. Halama (HM-2011-1034); 2. Ibidem, habitat as described above, 856 m a.s.l., 50°15'59"N, 16°47'51"E, 16.09.2011, leg. M. Halama (HM-2011-1034); 3. Poland, Ślęża Mountain (Zobtenberg), 10.1872, leg. L. Becker, det. J. Schröter as Nolanea pascua (WRSL-F 002073).
Taxonomic diversity and similar species
Noordeloos et al. (2022) assigned E. conferendum to the subgenus Nolanea (Fr.) Noordel. and the section Staurospora (Largent & Thiers) Noordel. Among the European representatives of Staurospora, only E. conferendum and Entoloma milthalerae M. Kamke & Lüderitz are known to exhibit cross-star-shaped spores. The latter species was recently described from oligotrophic, moist crowberry (Empetrum) heaths in Schleswig-Holstein, Germany (Lüderitz et al., 2016) and has also been recorded in Austria (Reschke et al., 2022). Entoloma milthalerae has not yet been confirmed in Poland, although some earlier records of E. conferendum in the country may in fact pertain to this species (cf. Noordeloos et al., 2022). Unfortunately, E. milthalerae remains a poorly known species. Its current interpretation (Noordeloos et al., 2022; Reschke et al., 2022) differs significantly from the original description. Initially, the species was reported as having a non-striate, non-hygrophanous pileus with a tomentose surface, the presence of cheilocystidia, significantly larger spores, and occasional clamp connections on the hyphae of the stipe (Lüderitz et al., 2016). According to Noordeloos et al. (2022), most of the holotype of E. milthalerae has been lost; however, a study of the remaining fragment revealed smaller spores and aberrant basidia, which correspond well with the previously reported so-called cheilocystidia. Despite its similarity to E. conferendum, E. milthalerae seems to be not its sister species according to phylogenetic analyses (Reschke et al., 2022). It should be noted, however, that the minor genetic differences have been observed between these taxa (with a p-distance not exceeding 2.0% based on ITS sequences data), which until recently, has not been considered sufficient grounds for distinguishing separate species within the genus Entoloma (e.g. Morozova et al., 2014). The smaller size of the fruit bodies (pileus ≤ 20 mm in diameter vs. 20–60 mm; stipe 30–50 × 1–1.5 mm vs. 25–80 × 2–6.5 mm), together with differences in spore size and shape (9–12 × 7.5–9 µm, Q = 1.1–1.5 – often irregularly cruciform-stellate with sharp angles, vs. 7.5–9.5 × 6–8 µm, Q = 1.1–1.3 – relatively regular and cruciform), seem to have key diagnostic value, allowing for the differentiation of E. milthalerae from E. conferendum (Noordeloos et al., 2022). However, the current study suggests that such morphological differentiation may not be entirely reliable. Although the analyzed specimens more closely match E. milthalerae in terms of spore size, the spore shape and the relatively large dimensions of the fruit bodies support their identification as E. conferendum. These findings, in agreement with those of other authors (e.g. Ćetković et al., 2020), indicate that the morphological delimitation of E. conferendum proposed in the most recent monograph of the genus Entoloma (Noordeloos et al., 2022) may be overly simplistic.
Entoloma conferendum exhibits a broad distribution and ecological amplitude (see below), as well as considerable morphological variability. This is reflected in the numerous synonyms of the species (Ludwig, 2007; Noordeloos, 1980, 1992). Among the several varieties and forms of E. conferendum (e.g., Bon & Courtecuisse, 1987; Courtecuisse, 2008; Jamoni & Bon, 1991; Largent, 1994; Noordeloos, 1980; Noordeloos & Hausknecht, 1998), two taxa deserve a brief discussion here, as their presence has been previously reported from Poland.
Entoloma conferendum var. pusillum (Velen.) Noordel. (syn. Rhodophyllus xylophilus J.E. Lange), is characterised by smaller and slenderer basidiomata (cap up to 15 mm in diameter, stipe 1–1.5 mm wide) (Noordeloos, 1980; Noordeloos et al., 2022). This taxon is now regarded as an occasional, aberrant form of E. conferendum fruiting on (very rotten) woody substrates (Noordeloos et al., 2022). In Poland, E. conferendum var. pusillum has been recorded several times to date (Domański et al., 1967; Gierczyk et al., 2018; Lisiewska, 1978, 1979; Łuszczyński, 2008). Noordeloos et al. (2022) point out that some reports of the presence of the wood-inhabiting variety on other substrates, particularly in swampy and Sphagnum dominated environments, may in fact represent E. milthalerae.
Entoloma conferendum var. incrustatum (Largent & Thiers) Noordel. & Hauskn. (syn. Nolanea staurospora var. incrustata Largent & Thiers) – taxon described from western USA (Largent & Thiers, 1972), was recently reported in Poland by Gierczyk et al. (2018). It differs morphologically from the typical variety in having a minutely incrusting pigment present on some of the slender hyphae in the pileal trama (Largent, 1994). However, this incrustation may occasionally be present in both E. conferendum and E. milthalerae, especially in narrow hyphae beneath the subpellis layer (Noordeloos et al., 2022). In the case of European collections, the aforementioned taxa lack incrusting pigment organised in ring-like patterns, as observed and described by Largent & Thiers (1972). Therefore, for European collections, the taxonomic status of E. conferendum var. incrustatum remains unclear, and it is currently not recommended to apply this name to European materials (Noordeloos et al., 2022).
A strong morphological resemblance to E. conferendum is also exhibited by Entoloma nothofagi G. Stev. and Entoloma botanicum by G. Stev., which occur in forest communities dominated by Nothofagus, Weinmannia, Metrosideros, and Dacrydium in New Zealand (Horak, 2008; Stevenson, 1962). Similarly, though possessing noticeably smaller spores, Entoloma brevispermum G.M. Gates & Noordel. is recognized as a species widespread in the moist sclerophyll forests of Australia and Tasmania (Gates & Noordeloos, 2007; Noordeloos & Gates, 2012). Subsequent morphological analyses of the holotypes of E. nothofagi and E. botanicum revealed no significant distinguishing features, and the differences observed in comparison to E. conferendum were deemed negligible (Noordeloos & Gates, 2012). Both taxa are now considered conspecific (Horak, 2008) and likely synonymous with E. conferendum (Horak, 1980; Noordeloos, 1980; Noordeloos & Gates, 2012). Molecular phylogenetic studies are needed to conclusively resolve the taxonomic status of E. nothofagi and E. botanicum within the Staurospora section and to better define the global distribution of E. conferendum.
At this point, it is also necessary to briefly address the name Agaricus pascuus Pers.: Fr., which frequently appears in older mycological literature (e.g., Schröter, 1885–1889). Noordeloos (1980) concluded that this name, along with its later homotypic synonyms (e.g., Nolanea pascua (Pers.: Fr.) P. Kumm., Rhodophyllus pascuus (Pers.: Fr.) Quél., Hyporrhodius pascuus (Pers.: Fr.) J. Schröt., Entoloma pascuum (Pers.: Fr.) Donk), represents a case of doubtful application. In the literature, it has been used to refer to a taxonomically complex group comprising at least three grassland species: E. conferendum, Entoloma sericeum (Bull. ex) Quél., and Entoloma vernum Lundell. Recently, however, the name Agaricus pascuus Pers.: Fr. has been recognised as an earlier homotypic synonym of Entoloma sericeum Quél. (Reschke & Noordeloos, 2022). In the “Checklist of Polish larger Basidiomycetes” (Wojewoda, 2003), E. pascuum is listed among the representatives of the genus Entoloma, categorized as a “doubtful taxon”. This publication does not provide a detailed interpretation of A. pascuus sensu J. Schröt., and only two (out of a total of 26) Polish localities of E. pascuum, originally reported in “Die Pilze Schlesiens” by Schröter (1885–1889) under the name Hyporhodius pascuus, are mentioned in a simplified manner. Additionally, data from two other authors, Chełchowski (1898) and Eichler (1900), are also included. Schröter’s (1885–1889: page 614) description of the taxon – particularly the spore dimensions and shape, clearly points to E. conferendum as the modern equivalent of A. pascuus ss. J. Schröt. This identification is further supported by micromorphological analysis of selected collections housed in the WRSL fungarium (e.g., WRSL-F 002073). Unfortunately, due to the overly general nature of the available information, an unambiguous taxonomic interpretation of the data published by other 19th-century authors regarding A. pascuus remains unattainable (cf. Chełchowski, 1888: page 81, 1898: page 155; Eichler, 1900: page 191; Zawadzki, 1835: page 168).
Distribution, phenology, and ecology
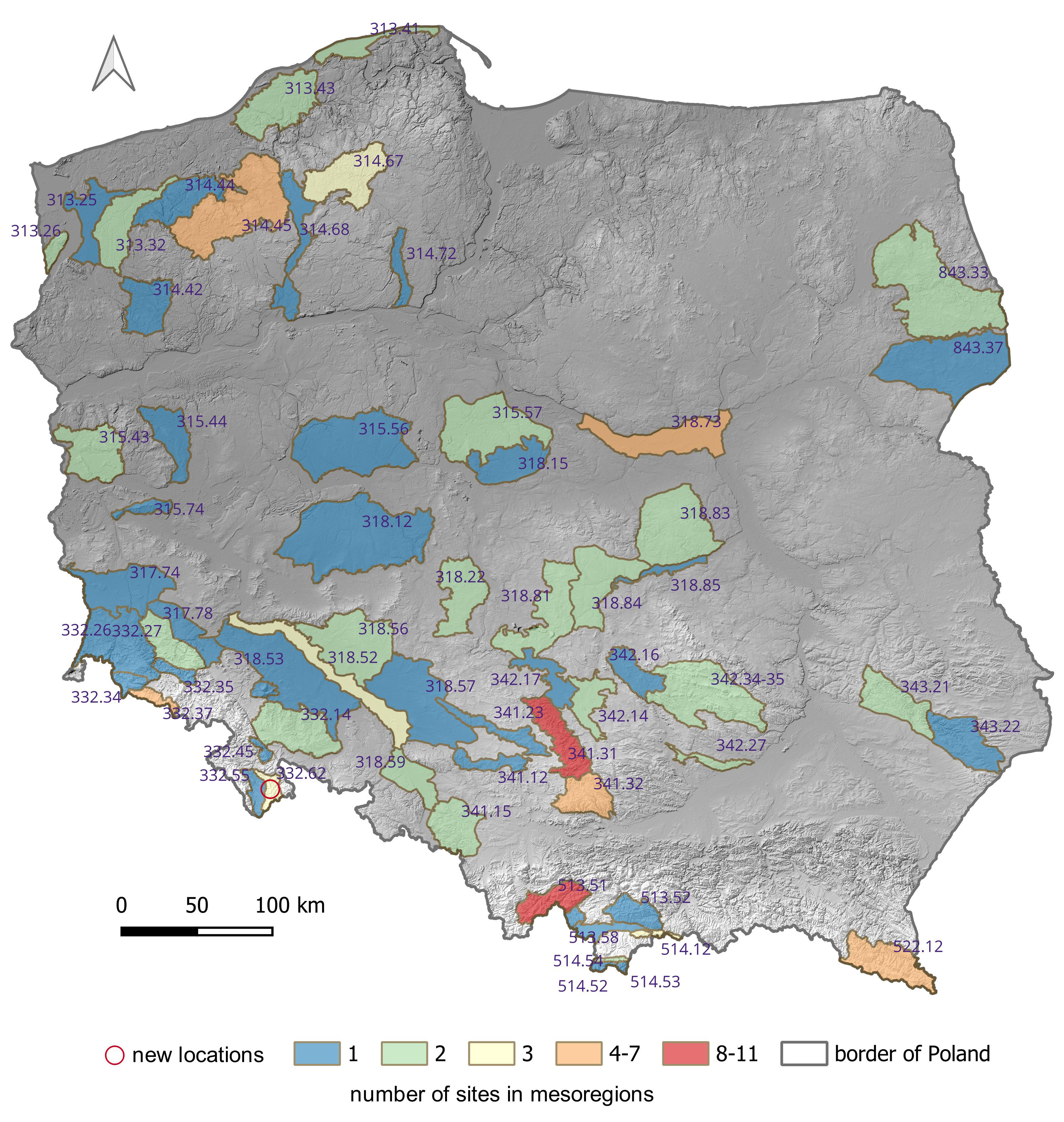
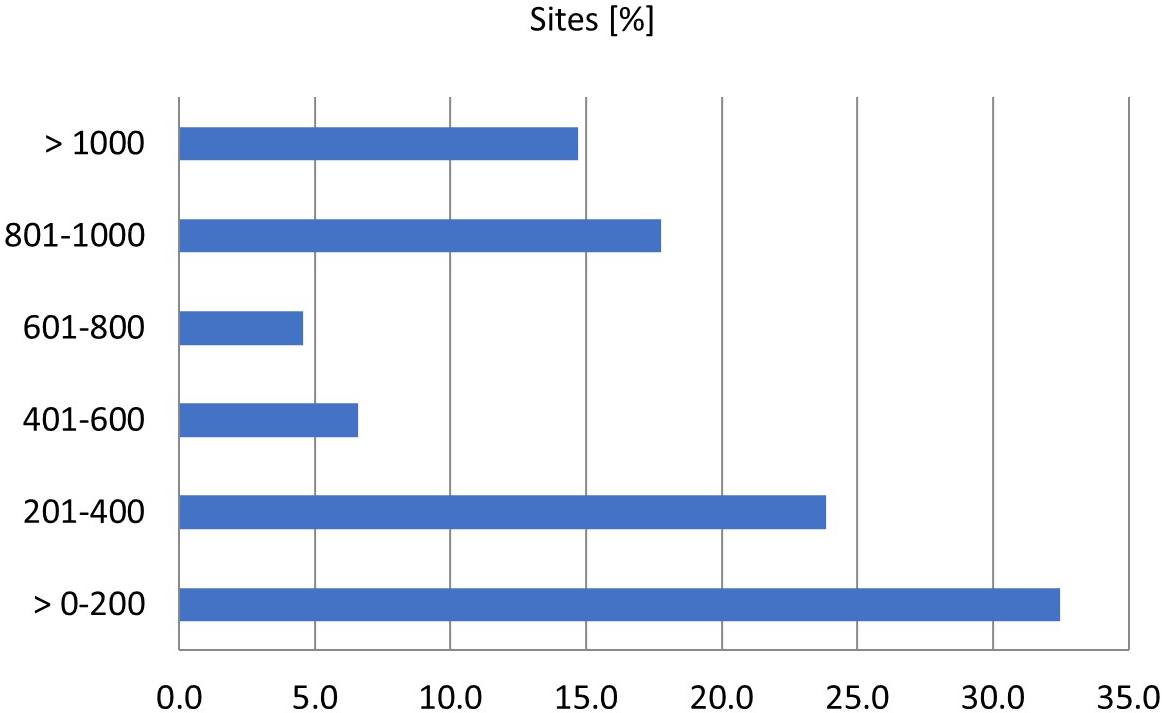
The natural range of E. conferendum is likely very broad, encompassing areas in both the Northern and Southern Hemispheres (Krieglsteiner, 2003; Noordeloos & Gates, 2012). This fungus is a widely distributed taxon in Poland. Its most numerous sites (4–11) are in the following mesoregions: Drawsko Lakeland, Warsaw Basin, Karkonosze Mts, Bieszczady Mts, Olkusz Upland, Częstochowa Upland, and Żywiec Beskid Mts (Figure 3). Entoloma conferendum seems to demonstrate a very wide ecological amplitude, both in terms of the habitats it occupies and other environmental parameters, including climatic and geological-soil conditions (see below). The vertical distribution of the Polish sites of the species varies and includes both lowland and mountainous areas (Figure 4). The highest occurrences of this species in Poland are found in the Tatra Mountains and the Babia Góra Massif, occupying habitats at elevations between 1100 and 1900 meters above sea level, including alpine environments (Bujakiewicz, 1979; Frejlak, 1973; Nespiak, 1960, 1962; Ronikier, 2012). Although a significant proportion (over 23%) of E. conferendum sites are located at elevations between 751 and 1 000 meters above sea level – including, among others, records from the Śnieżnik Massif – the majority of the species' recorded locations in Poland lie below 500 meters above sea level (approximately 56%).
Figure 3
Distribution of sites of Entoloma conferendum in Poland against the background of mesoregions (codes of mesoregions after Solon et al., 2018; see Appendix A for additional information).
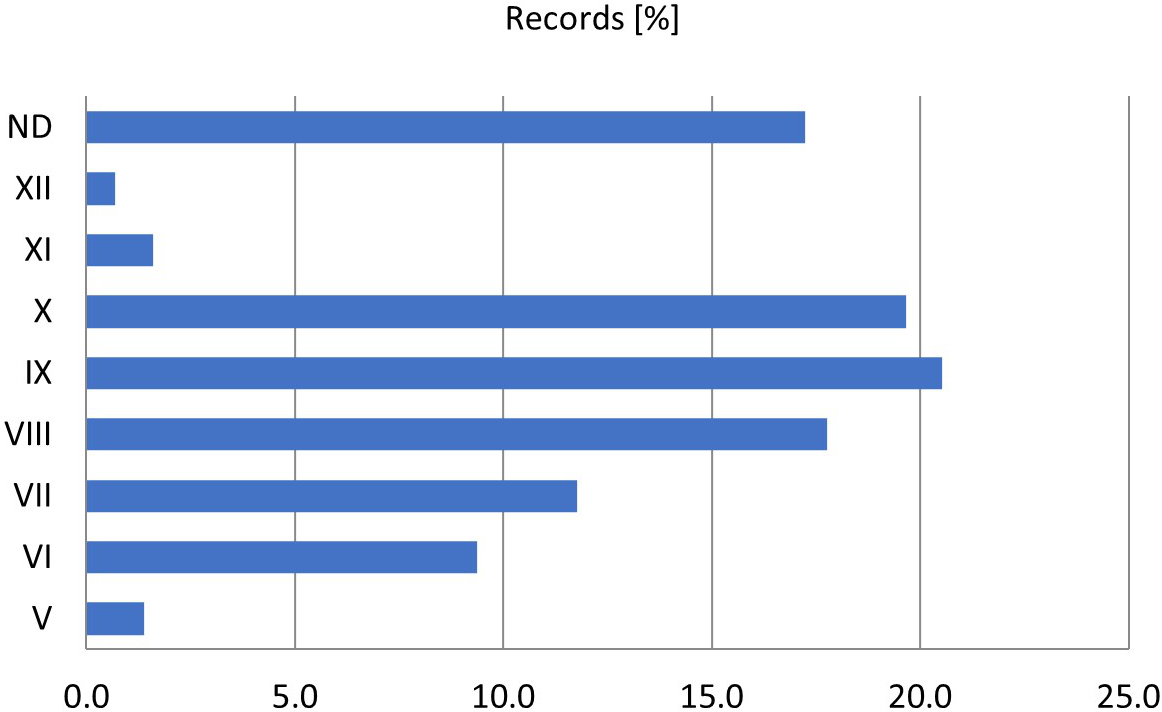
In general, basidiomata of E. conferendum appear singly or in small groups from April to November, although there are also some records of this fungus in January and December (Krieglsteiner, 2003; Ludwig, 2007). A similar phenology is observed in Poland (Figure 5), where E. conferendum fruits from spring to late autumn, with the peak period being from August to October (more than 58% of classified records).
In Europe, E. conferendum is considered a species of fresh and moist habitats, developing among mosses, grasses, and shrubs, and is found almost exclusively on acidic or at least surface-acidified soils (brown, podzol, peat), characterised by low calcium carbonate and nutrient content. The habitats of the species on the European continent include the interiors and edges of various forest types (coniferous, mixed, and deciduous), parks, clearings, poor meadows and pastures, and raised bogs (Krieglsteiner, 2003; Ludwig, 2007; Noordeloos, 1992; Noordeloos et al., 2022). The sites presented in this study from the Śnieżnik Massif, which include extensively used and unfertilized meadows, confirm the preference of the species for habitats characterised by low nutrient content in the soil. Available data indicate that E. conferendum in Poland is most often recorded in forested and shrub habitats. These environments constitute approximately 74% of classified sites for this species (Adamczyk, 2011; Bujakiewicz, 1979; Bujakiewicz & Kujawa, 2010; Bujakiewicz & Lisiewska, 1983; Domański et al., 1960, 19630, 1967; Domański, 1965, 1999; Friedrich & Orzechowska, 2002; Gierczyk et al., 2009; Gierczyk et al., 2018; Grzesiak et al., 2017; Karasiński et al., 2015; Kujawa & Gierczyk, 2016; Lisiewska, 1979; Ławrynowicz, 1973; Nespiak, 1960; Ronikier, 2012; Rudnicka-Jezierska, 1969; Schröter, 1885–1889; Stasińska, 2011; Stasińska & Sotek, 2017; Szkodzik, 2005; Ślusarczyk, 2007, 2019; Wojewoda, 1974; Wojewoda et al., 2004). Non-forest communities represent 21% of the sites of E. conferendum in Poland (Adamczyk & Kucharski, 2005; Frejlak, 1973; Gierczyk et al., 2009; Grzesiak et al., 2017; Gumińska, 1972; Karasiński et al., 2015; Kujawa et al., 2019; Łuszczyński et al., 2022, presented data; Schröter, 1885–1889; Stasińska, 2011). The remaining portion (a total of 5%) consists of unclassified habitats and park-garden type habitats (Flisińska, 2004; Gierczyk et al., 2018; Karasiński et al., 2015; Kryza & Puciata, 2009; Lisiewska & Nowicka, 1979; Pietras et al., 2016). Among the forest-shrub habitat categories of E. conferendum in Poland, 17 types of phytosociological associations have been distinguished (Abieti-Piceetum, Calamagrostio villosae-Pinetum, Dentario glandulosae-Fagetum, Fraxino-Alnetum, Galio-Abietetum, Leucobryo-Pinetum, Luzulo luzuloidis-Quercetum, Pinetum mughi carpaticum, Plagiothecio-Piceetum, Potentillo albae-Quercetum, Querco roboris-Pinetum, Ribeso nigri-Alnetum, Serratulo-Pinetum, Sorbo-Aceretum, Tilio-Carpinetum, Vaccinio uliginosi-Betuletum pubescentis, Vaccinio uliginosi-Pinetum) as well as 11 types of more general or simplified defined vegetation communities (e.g., unspecified forest, swamp forest, deciduous forest, mixed forest, pine forest, etc.). The highest proportion of E. conferendum sites are found in “unspecified” forest phytocoenoses (15%) and a group of 11 associations and communities, including: upper montane Carpathian spruce forest, mesotrophic fir forest, lower montane spruce-fir forest, dwarf pine thickets, subcontinental oak-hornbeam forest, swamp birch forest, and submontane acidophilous oak forest (35%). The proportion of reported E. conferendum sites in Poland representing non-forest habitats is 21%. These include meadow and grassland communities (Adonido-Brachypodietum, Thalictro-Salvietum), pastures, roadsides, clearings, fallow lands, and ombrotrophic moss communities (Sphagnetum magellanici). The last category, which accounts for the smallest share of E. conferendum sites (>5%), consists of records lacking habitat information and habitats reported as parks and garden environments.
. Conclusions
Entoloma conferendum has a wide global distribution, including most temperate zones of both hemispheres, and is widespread from the lowlands to the alpine zones. The species seems to be a very common fungus in Poland. Although the taxon exhibits significant morphological variability, it also possesses a distinctive set of characteristics that allow for its unequivocal identification, among which the most important role is played by its unique – regular cruciform-stellate spore form. The known varieties of the species reported from Poland, such as var. pusillum and var. incrustatum exhibit the following main differences: the first is characterised by woody substrate affinity, while the second is distinguished by the presence of a minutely incrusting pigmentation on some of the slender hyphae of the pileal trama. However, the taxonomic status of the latter taxon in Europe (and in Poland) remains uncertain, and this name is currently advised against using it for European materials. Nomenclaturally, H. pascuus sensu J. Schröt., must be considered as a misapplied name of A. conferendus Britz., which is consistent with data from the literature and results from morphological characteristics of the collections studied. Within the subgenus Nolanea and the Staurospora section, E. conferendum stands out alongside E. milthalerae due to its unique spore morphology, however, the latter differs from related E. conferendum, apart from other habitat preferences, in its larger and more irregular spores.
Entoloma conferendum is very common fungus, which demonstrates an amazing wide ecological amplitude, thriving in diverse habitats across Poland. In this country, the species is predominantly found in forested and shrub habitats, which constitute approximately 74% of its recorded sites. Non-forest habitats account for 21% of the records, while the remaining 5% represent unclassified or park-garden environments. The fungus appears to prefer acid soils and habitats with a low soil nutrient content. It is found across various elevations, from lowlands to mountainous regions (up to 1900 m above sea level). Its fruiting season spans from spring through late autumn, with peak occurrences observed from August to October.
Further research should focus on molecular analyses to clarify the taxonomic relationships within the Staurospora section and validate the distinctiveness of the E. conferendum variants on a global scale. Additionally, expanding ecological studies can improve understanding of the habitat preferences of the species and its ecological roles within fungal communities.
. Supplementary material
The following supplementary material is available for this article:
Appendix A. Polish localities of Entoloma conferendum documented in the study.
Acknowledgments
The author gratefully acknowledges Mr. Czesław Narkiewicz for the opportunity to discuss the distribution of the studied species in Lower Silesia. This research was funded through private means and co-financed by statutory funds from the Museum of Natural History, University of Wrocław. I also extend sincere thanks to the anonymous reviewers for their constructive comments.
