. Introduction
Oat (Avena sativa L.) is a species of cereal grain primarily cultivated for animal feed (Cuddeford, 1995). In recent years, oats have gained popularity in the human diet due to their health-promoting properties, including a high content of nutrients such as β-glucan (Wang & Ellis, 2014). This compound is found in products made from whole grains and has beneficial effects on human health, including its ability to regulate cholesterol levels (Luithui et al., 2019; Stewart & McDougall, 2014).
Despite their advantages, oats are sensitive to many biotic and abiotic factors (Pandey et al., 2017; Ramegowda & Senthil-Kumar, 2015; Suzuki et al., 2014), especially fungal diseases. One of the most serious fungal pathogens is Blumeria graminis (powdery mildew), an obligate biotrophic fungus that attacks oat leaves (Dean et al., 2012; Troch et al., 2013). Fungal infections cause crop losses, which can reach up to 40% but usually remain around 10% (Aung et al., 1977; Jones et al., 1987). These losses reduce both the quantity and quality of the yield. Therefore, effective protection of oats against powdery mildew is one of the most crucial challenges in crop cultivation. There are currently two ways to combat powdery mildew: using chemical fungicides and through genetic methods. Unfortunately, there is insufficient knowledge about how resistance gene mechanisms work, the limitations of using R genes, or their possible combinations in breeding programs (Montilla-Bascón et al., 2015; Zuccaro & Langen, 2020). To develop effective protection strategies against powdery mildew, it is essential to first understand its biology and infection mechanisms.
. Biology and epidemiology of powdery mildew
B. graminis is a fungus that depends on living plant cells for survival. These cells are necessary for its growth and reproduction. As a result of evolution, B. graminis hasdeveloped eight different specialized forms (Oku et al., 1985; Schulze-Lefert & Panstruga, 2011; Wyand & Brown, 2003), each adapted to a specific host (Anon, 1912; Liu et al., 2021). One of them is B. graminis f. sp. avenae, which attacks oat leaves and causes a disease called powdery mildew.
The infection cycle of B. graminis (Figure 1) begins with the development of an asexual spore (conidium). This spore germinates and develops in a strictly regulatedway (Green et al., 2002). The germination process initiates the formation of a primary germ tube. The next step is the formation of a secondary germ tube – an appressorial tube. This structure differentiates and elongates, forming a hooked appressorium at its tip within approximately 12 hours.
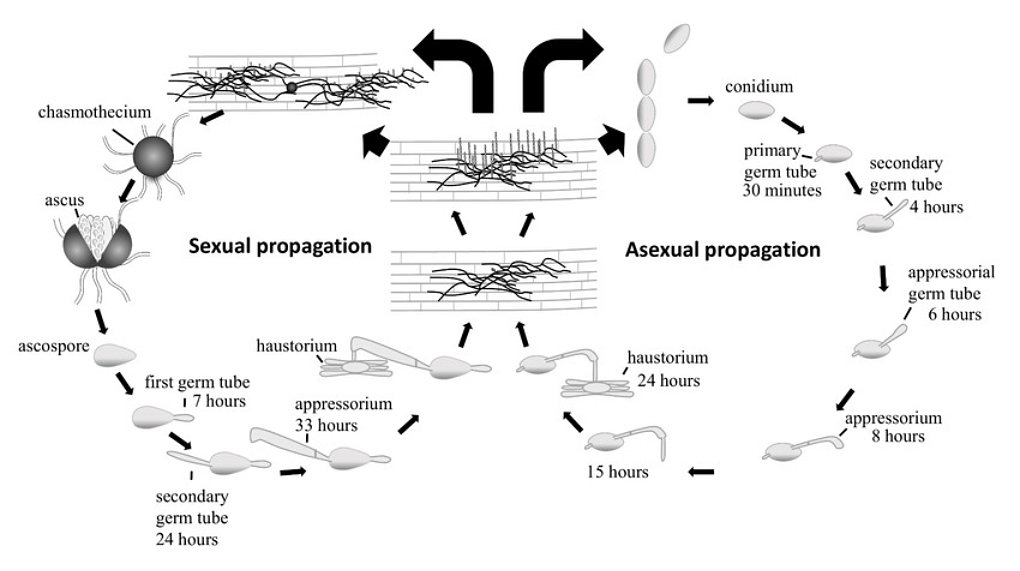
From the appressorium, a penetration peg grows, piercing the cell wall of the plant’s epidermis. This occurs due to a combination of mechanical pressure and enzymatic degradation. Once penetration is complete, the penetration peg enters the cell. At this stage, the tip of the peg swells, forming a growing haustorium.
Over the next 4–5 days, the haustorium develops multiple finger-like structures, which are essential for nutrient uptake from the host’s epidermal cells.
At the same time, outside the cell, secondary hyphae develop, which are responsible for forming the next generation of haustoria (by the third day of infection) and conidiophores, which produce new spores starting from the fourth day (Jennings et al., 2024; Sánchez-Martín et al., 2011).
The short life cycle of B. graminis allows it to produce new spores within 4–5 days after infection, enabling rapid spread of the pathogen. In temperate climate regions, B. graminis poses a significant problem for crops. Its occurrence is most prominent in Europe and North America (Dean et al., 2012). Understanding the pathogen’s biology reveals why it spreads rapidly, but equally important is understanding how plants defend themselves against attacks. Plant resistance operates at multiple levels, from basic recognition systems to complicated mechanisms of genetic responses.
. Mechanisms of plant resistance
. Innate immunity
During plant evolution, plants have developed two strategies for pathogen detection (Chisholm et al., 2006; Jones & Dangl, 2006). On plant cell surfaces, there are special receptors called Pattern Recognition Receptors (PRRs) (Akira et al., 2006). These receptors detect unique molecules from pathogens, known as Pathogen-Associated Molecular Patterns (PAMPs). PAMPs are essential compounds found in entire groups of pathogens, such as bacterial flagellin or fungal chitin. Plants also respond to pieces of their own cell walls or cuticles released during infection, known as Damage-Associated Molecular Patterns (DAMPs) (Tanaka & Heil, 2021). The activation of PRRs induces immunity triggered by PAMPs (PTI), which helps the plant fight pathogens (Boller & Felix, 2009; Pastor et al., 2022).
The second mechanism of plant perception involves the recognition of specific pathogen virulence molecules, called effectors, by receptors inside plant cells. This mechanism initiates Effector-Triggered Immunity (ETI). This type of response creates a unique, coevolutionary dynamic between the plant and the pathogen. The difference between PTI and ETI is that the effectors in ETI responses are more diversified than those in PTI and are not essential for the pathogen’s survival. ETI receptors and pathogen effectors exhibit intense diversity within and between species, whereas some PRR receptor functions are conserved across plant families (Dodds & Rathjen, 2010; Yuan et al., 2021). Both PTI and ETI responses trigger similar reactions. However, ETI is usually stronger, faster, and is associated with the Hypersensitive Response (HR). HR is responsible for local cell death, which limits the spread of infection. PTI is effective against pathogens that have not evolved to the plant-host relationship, resulting in immunity known as non-host immunity. On the other hand, ETI is activated in response to pathogens adapted to the plant species. This correlation is flexible and depends on the specific molecules that signal the presence of infection (Yu et al., 2024). While innate immunity provides the first line of defense at the site of infection, plants have also evolved mechanisms to extend the protection throughout the entire organisms.
. Acquired immunity
Local Acquired Immunity (LAR) in plants is a mechanism activated near the site of pathogen infection (Jacob et al., 2023). LAR provides defense around damaged cells and offers long-term protection against subsequent pathogen attacks. LAR includes the induction of proteins associated with pathogenesis, as well as the production of secondary metabolites and salicylic acid (SA). This situation creates a favorable environment for Systemic Acquired Resistance (SAR). An important role in this process is played by SA, which is essential for both local and systemic immunity (Gao et al., 2015; Jacob et al., 2023).
When plants are attacked by pathogens, they can initiate LAR at the site of infection. Over time, this reaction can spread throughout the entire plant, leading to SAR. The key role in this process is played by SA, which drives signals from the infected area to other regions of the plant via conductive tissues. SA activates changes in plant genes responsible for defense mechanisms, providing long-term protection against pathogens and enhancing immunity against future infections (Gao et al., 2015; Wenig et al., 2019; Zhou & Zhang, 2020).
Another form of systemic immunity is Induced Systemic Resistance (ISR). ISR is stimulated by plant-growth-promoting bacteria (such as Pseudomonas and Bacillus), arbuscular mycorrhizal fungi, and synthetic compounds. ISR works through a process called “priming”, which prepares the plant for a faster and stronger response to future pathogen attacks. This mechanism operates via signal pathways dependent on jasmonic acid (JA) and ethylene. These compounds make the defense more effective and help the plant prepare for subsequent biotic stress (Costet et al., 1999; Pieterse et al., 2014; Yu et al., 2022). To better understand the immune mechanisms occurring in the plant, a schematic diagram (Figure 2) was prepared and is presented below.
Figure 2
Schematic summary of the plant immune system (Plant Defense Signaling Network: From Recognition to Whole-Plant Immunity)
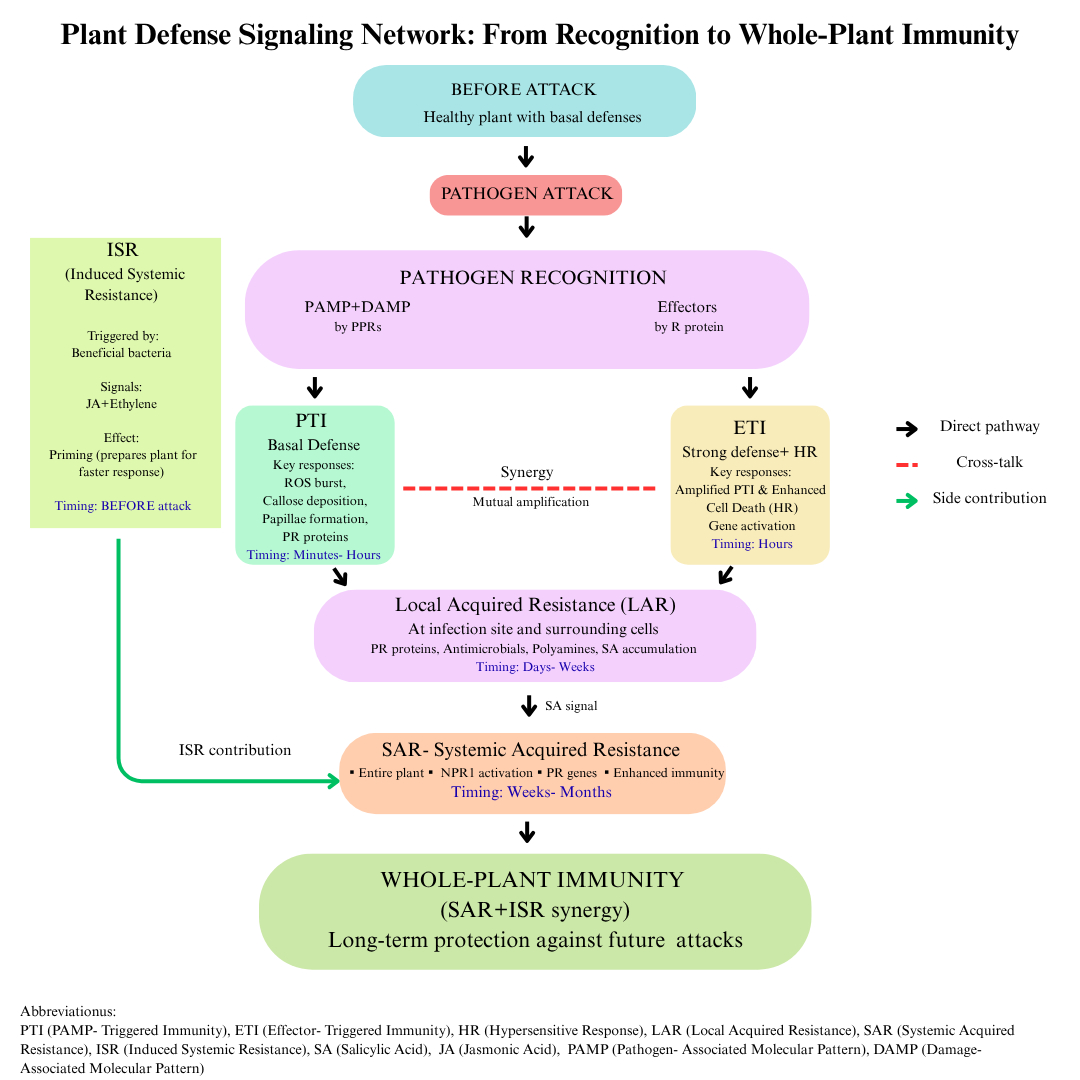
The diagram summarizes the main stages of the plant immune response. Recognition of PAMPs and DAMPs by pattern recognition receptors (PRRs) initiates PAMP-triggered immunity (PTI), characterized by early defense responses such as ROS burst, callose deposition, and PR protein synthesis. Pathogen effectors are detected by resistance (R) proteins, leading to effector-triggered immunity (ETI), a stronger response often associated with the hypersensitive reaction (HR). Mutual reinforcement of PTI and ETI results in local acquired resistance (LAR) at the infection site. Systemic responses include systemic acquired resistance (SAR), mediated by salicylic acid (SA), and induced systemic resistance (ISR), triggered by beneficial microbes via jasmonic acid (JA) and ethylene (ET). The cooperation of SAR and ISR leads to whole-plant immunity, providing long-term protection against subsequent infections. Understanding these immune layers provides the foundation for identifying genetic determinants of resistance, which are critical for breeding programs.
. Genetic response
In accordance with the gene-for-gene interaction model (Figure 3), a defensive reaction in plants occurs when a resistance gene (R gene) in the plant is compatible with an avirulence gene (Avr gene) in the pathogen (Dodds, 2023; Nürnberger et al., 2004). The dominant resistance gene (R) in the plant confers immunity by recognizing the matching Avr gene product and activating defense mechanisms. Similarly, the dominant avirulence gene (A) in the pathogen determines its recognition by the plant’s R gene, thereby triggering host defenses, whereas the recessive allele allows the pathogen to escape detection and establish infection. If either the Avr or R gene is absent or mutated, disease development may occur (Garcia-Brugger et al., 2006; Valent, 2025).
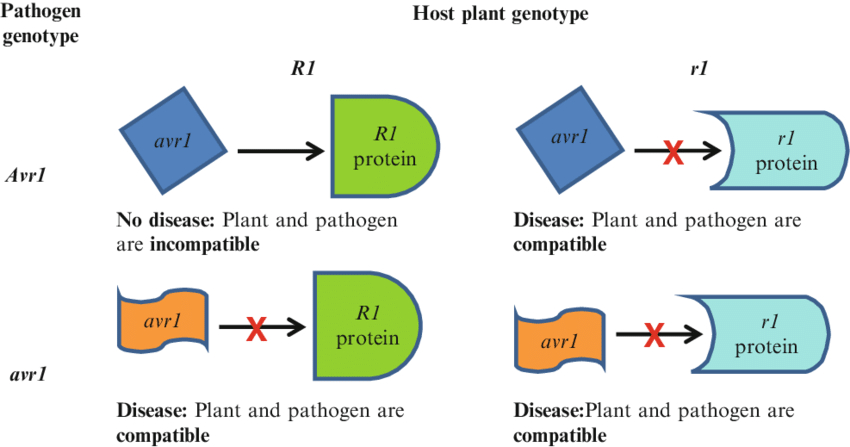
The Avr gene encodes virulence proteins, while the R gene is responsible for producing intracellular immune receptors, specifically NB-LRR proteins. NB-LRR proteins can recognize Avr proteins in infected plant cells. The R protein interacts with the pathogen either directly or indirectly through target molecules. Structural changes in these target molecules influence the function of the R gene and the activation of immune responses (Sharma & Gupta, 2020).
Despite the identification of many R genes in plants, pathogens have the ability to evolve rapidly, allowing them to evade recognition by plant R genes. Consequently, this evolution increases the risk of pathogens overcoming plant immunity (Campos & Félix, 2023; Friedman & Baker, 2007).
The gene-for-gene model illustrates plant defense responses depending on the compatibility of plant and pathogen genotypes. When the plant resistance gene R1 matches the pathogen avirulence gene Avr1, defense mechanisms are activated, resulting in no disease (plant and pathogen are incompatible). In other combinations (e.g., r1 and Avr1 or R1 and avr1), recognition fails, leading to disease and compatibility between the plant and pathogen (compatible). The red “X” indicates the lack of recognition between the resistance protein and the avirulence protein. In oats specifically, researchers have identified and catalogued multiple resistance genes that follow this gene-for-gene model.
Thirteen genes resistant to powdery mildew have been identified and cataloged in oats, designated as Pm genes (Ociepa et al., 2020; Ociepa & Okoń, 2022; Schurack et al., 2024). These genes originate from several wild oat species: Pm1, Pm3, Pm11, and Pm12 come from Avena sterilis (Hsam et al., 1998; Hsam et al., 2014; Ociepa et al., 2020; Ociepa & Okoń, 2022); Pm2 is derived from A. hirtula (Herrmann & Mohler, 2018); Pm4 from A. barbata (Hsam et al., 2014); Pm5 from A. macrostachya (Yu & Herrmann, 2006); Pm7 from A. eriantha (also known as A. pilosa); and Pm8 was identified in the Rollo variety (Hsam et al., 2014). Meanwhile, Pm6, Pm9, and Pm10 originate from A. byzantine (Herrmann & Mohler, 2018; Hsam et al., 2014), and Pm13 was found in the ‘Husky’ variety (Schurack et al., 2024).
Quantitative resistance, which is temporary and incomplete, is based on multiple minor-effect genes. Each of these genes enhances plant resistance and makes it more difficult for new pathogen strains to overcome immunity (Sánchez-Martín et al., 2011). These genes may contribute to PTI (Pattern-Triggered Immunity) responses. ETI (Effector-Triggered Immunity) responses can also play a role in quantitative resistance, especially when some R genes weaken due to pathogen evolution (Cowger & Brown, 2019). Unfortunately, at this time, there is no information about the mechanisms by which these genes function.
The R genes identified in oats activate defense mechanisms through signal transduction cascades. These cascades ultimately result in physical barriers at the cellular level that prevent pathogen entry from the very beginning of the infection process.
. Penetration resistance
Resistance to penetration in the cell area is a key defense mechanism that prevents fungal pathogen invasion. This resistance is known as Cell Wall Adhesion (CWA), which involves the formation of papillae. Papillae act as a barrier that blocks pathogenpenetration (Qin et al., 2021; Schmelzer, 2002; Zeyen et al., 2002). Local CWA occurs through the targeted accumulation of callose and the release of antimicrobial substances. Research has shown that the accumulation of subcellular components at the site of infection depends on the functioning of the plant cytoskeleton. This has been confirmed by studies on plant-fungus interactions (Hardham et al., 2007).
Research on oats (Montilla-Bascón et al., 2014) has shown that the level of putrescine increases within 24 hours after inoculation with B. graminis f. sp. avenae in both resistant and susceptible varieties. Despite being classified as a susceptible variety, Selma has exhibited moderate resistance, suggesting an important role of putrescine in the defense mechanism. Moreover, the application of putrescine enhances Selma’s resistance to a level observed in the resistant variety Charming. In Charming, an increase in spermidine levels has also been observed during papilla formation, which positively influences penetration resistance.
The results have shown that polyamines, such as putrescine, spermidine, and 1,3-diaminopropane (DAP), play an important role in the oat resistance response to B. graminis f. sp. avenae, especially in the early stages (Cowley & Walters, 2002; Hückelhoven, 2007; Prats et al., 2005). A rapid increase in putrescine levels after inoculation could be key to an effective defense, as it leads to the fast activation of defensive mechanisms. The speed of defensive mechanism activation is crucial in the oat–powdery mildew interaction (Hückelhoven, 2007; Majumdar et al., 2019). These penetration resistance mechanisms are part of broader defense strategies that cereals have developed specifically against powdery mildew infections.
. Defensive mechanisms in cereals
Two main defensive mechanisms observed in cereals against powdery mildew are the development of papillae and cell death in pathogen-attacked cells. Papillae form due to cell wall modifications, which aggregate inside the epidermal cell wall area near the pathogen’s appressoria. They act as a physical and/or chemical barrier, making pathogen penetration more difficult (Zeyen et al., 2002). Cells that successfully form effective papillae will survive.
The structure of papillae contains a callose matrix, which is built from various compounds, especially proteins and autofluorescent phenolic compounds (Bhuiyan et al., 2009). The effectiveness of these mechanisms depends on the speed of papilla growth, as well as the quality and quantity of their components (Zeyen et al., 2002).
The second defense mechanism is the death of epidermal cells, which occurs after pathogen penetration or as a result of papilla activity. This reaction stops haustoria development and prevents the formation of pathogen colonies. In barley, the interaction between an avirulent powdery mildew isolate and the host plant leads to a hypersensitive response (HR). The speed of cell death depends on specific loci or alleles responsible for immunity (Boyd et al., 1995; Kruger et al., 2002). The efficiency and timing of this reaction vary depending on gene alleles and macroscopic response phenotypes (Kruger et al., 2002). A delay in haustoria formation can contribute to post-haustorial immunity, which further inhibits the development of secondary hyphae and the sporulation of pathogen colonies (Jennings et al., 2024; Niks & Rubiales, 2002). While plants have evolved defense mechanisms, pathogens have simultaneously developed counter strategies to overcome these barriers and successfully infect their hosts.
. Pathogen attack
A pathogen is capable of recognizing its host, and thanks to chemical interactions with the plant system, the plant can also detect the pathogen. However, the mere presence of a susceptible host and a virulent pathogen is insufficient to cause disease. Favorable environmental conditions and a sufficient quantity of inoculum are necessary to infect the plant (Surico, 2013). The inoculum plays a key role because it enables an infection to develop.
During the host-pathogen interaction, the pathogen may be repelled by the plant’s defense mechanisms or may manipulate the plant’s biological processes (Goyal & Manoharachary, 2014). When the pathogen manipulates the plant, it creates a favorable environment for its growth and reproduction. Usually, the pathogen associates with the plant by occupying external cellular niches. However, it still extracts nutrients from the living host cells. Since the host’s cytoplasm and organelles play a key role in molecular interactions, the pathogen strives to penetrate the entire plant (Dodds & Rathjen, 2010).
Fungi, such as B. graminis, frequently transition between different environments, from moist conditions to air, which enables them to adhere to the plant cell surface. The pH of the environment has a significant influence on the growth and development of pathogenic fungi (Alkan et al., 2013). Additionally, pH changes play a key role in the infection process in plants. During an infection, the fungus causes alkalization of plant tissues, increasing the pH and enhancing the pathogen’s ability to infect (Fernandes et al., 2017). A fungal factor called RALF can further elevate the pH around the plant, supporting pathogen invasion by activating important protein kinases (Penalva et al., 2014).
Depending on their mode of nutrition, plant pathogens employ different invasion strategies (Lo Presti et al., 2015). Biotrophic pathogens, such as B. graminis (Meadows, 2011), require living host cells to complete their life cycle. This fungus disrupts the plant’s physiological processes to extract as many nutrients as possible. It secretes specialized factors, known as effectors, which suppress plant immunity while minimizing damage to the host cells. This process enables the colonization of living plant tissues (Lo Presti et al., 2015; Lowe & Howlett, 2015).
Some pathogens are capable of producing effectors that interfere with the host’s physiology and metabolism by disrupting cytoskeleton formation. For example, B. graminis f. sp. hordei can manipulate the spatial structure of the plant cytoskeleton. Its primary target is microtubules, which facilitate the pathogen’s entry into the host cell (Nottensteiner et al., 2018). Given the complex nature of the oat powdery mildew interaction and the continuous evolutionary arms race between the host and the pathogen, ongoing research is essential to develop effective and sustainable disease management strategies.
. Further research and perspective
There are many methods to prevent fungal diseases; however, none of them can provide 100% protection for the plant. Additionally, there is no guaranteed way to completely eliminate the pathogen. The primary methods of disease prevention include appropriate agrotechnical treatments (such as crop rotation, optimal sowing dates, and suitable pre-crops), chemical methods (fungicides), and the use of genetic resistance (Sánchez-Martín et al., 2011).
The application of genetic control is more environmentally friendly because it reduces the need for fungicides and lowers agricultural production costs (Aktar-Uz-Zaman et al., 2017; Goutam et al., 2015).
Breeding techniques currently focus on increasing innate immunity in plants and preventing the loss of efficiency in the R-Avr system. These efforts include the introgression of new resistance genes from wild plants and R-gene pyramiding, which involves introducing several resistance genes into a plant simultaneously (Pietrusińska & Czembor, 2015; Sharma & Gupta, 2020).
The presence of resistance genes in a plant enables the effective identification of avirulence factors. A better understanding of the mechanisms by which these genes function will contribute to a more effective use of genetic resistance in plant breeding. The urgency of this research becomes even more apparent when we consider the broader context of climate change and evolving agricultural practices. Understanding these molecular mechanisms provides the foundation for developing practical breeding strategies. Modern oat improvement programs must integrate this theoretical knowledge with field-based approaches.
. A practical approach to oat breeding
. Monitoring pathogen populations and resistance management
Effective deployment of resistance genes in oat (Avena sativa L.) cultivation requires an integrated approach combining resistant genotype identification with systematic pathogen population monitoring and strategic resistance management. As demonstrated by Grzelak, Nucia and Okoń (2025) in their study of B. graminis f. sp. avenae populations in Poland, regular surveillance enables early detection of shifts in pathogen virulence patterns toward Pm resistance genes and facilitates assessment of population dynamics.
Population monitoring focused on both plant resistance genes and pathogen virulence factors allows breeders to detect resistance breakdown caused by increasingly virulent strains that emerge through evolutionary processes (Babayants et al., 2015; Traskovetskaya et al., 2019). Given the high variability and adaptive potential of pathogens, continuous surveillance remains essential for successful breeding program implementation.
Eco-evolutionary models and empirical studies indicate that crop structure, host genetic diversity, and the presence of non-host or cover plants can decelerate pathogen evolution and reduce epidemic intensity, thereby enhancing resistance durability in field conditions (Bargués-Ribera & Gokhale, 2020). The reciprocal interactions between host-pathogen co-evolutionary processes and agricultural practices such as crop rotation exert selective pressures that impede aggressive strain fixation and maintain epidemiological equilibrium.
Monitoring data also enables optimization of resistance gene deployment through pyramiding, rotation, or genotypemixing.
Reilly et al. (2024) found that the most commonly cultivated oat variety in Ireland lacked effective resistance against powdery mildew. Currently, the cultivar ‘Husky’, which carries the Pm13 gene, is recommended for cultivation. Although this cultivar does not exhibit complete resistance to B. graminis f. sp. avenae, the presence of Pm13 significantly reduces disease incidence, decreasing reliance on chemical plant protection and contributing to improved yield stability.
Integrating pathogen monitoring into breeding and crop protection programs promotes durable resistance, enables dynamic responses to shifts in pathogen population structure, and represents a key component of sustainable resistance management under increasing environmental pressure and ongoing climate change.
While population monitoring and strategic gene deployment provide crucial foundations for resistance management, recent advances in molecular biology offer complementary tools for enhancing disease resistance at the genetic level. Among these innovations, genome-editing technologies have emerged as promising approaches for achieving precise modifications in resistance-related genes.
. Modern techniques used in breeding programs
Fungal pathogens pose serious threats to cereal crops, causing diseases such as powdery mildew, rust, and blight that result in substantial yield losses and grain quality deterioration. In response to these challenges, genome-editing technologies – particularly the CRISPR/Cas9 system – have attracted considerable attention in recent years, enabling precise modification of genes governing plant resistance to fungal infections (Zaynab et al., 2020).
The MLO gene represents one of the best-characterized targets for genome editing in cereals. This gene encodes a protein with seven transmembrane domains and is highly conserved across cereal species. Studies have demonstrated that CRISPR/Cas9-generated mutations in the TaMLO-A1 gene confer enhanced resistance to B. graminis f. sp. tritici, the causal agent of wheat powdery mildew (Wang et al., 2014). Similar outcomes have been reported in barley, Arabidopsis thaliana, and tomato, where MLO gene function loss resulted in durable powdery mildew resistance (Piffanelli et al., 2004).
In oats, however, research on CRISPR/Cas9 applications for enhancing resistance to B. graminis f. sp. avenae remains limited. Nevertheless, given the high degree of MLO gene conservation among cereals, comparable gene-editing strategies are anticipated to prove successful in oats.
Beyond direct gene editing, comprehensive genomic approaches provide essential tools for identifying resistance-associated loci and accelerating breeding progress.
Genome-wide association studies (GWAS) enable integration of high-density genotypic and phenotypic data to identify quantitative trait loci (QTL) regions and markers associated with desirable traits (Hamblin et al., 2011). When genomes are saturated with molecular markers, mapping can achieve single-gene resolution (Brachi et al., 2011). In oats, however, QTL research has predominantly focused on grain quality and yield traits rather than pathogen resistance (Ardayfio et al., 2025).
Despite significant advances in identifying markers and loci associated with agronomic traits, improving complex characteristics such as disease resistance remains challenging and time-consuming. Quantitative traits are typically controlled by numerous small-effect genes, limiting their practical breeding application (Merrick et al., 2021). Although marker-assisted selection (MAS) facilitates mapped gene utilization in breeding, its efficiency for polygenic traits is often limited (Hasan et al., 2021; Heffner et al., 2009). Conversely, genomic selection (GS), originally developed for animal breeding, enables breeding value prediction based on genome-wide marker information without requiring specific QTL identification (Su et al., 2010). GS implementation in oat breeding has demonstrated improved selection accuracy and accelerated genetic gain (Asoro et al., 2013; Dhakal et al., 2024)
Integrating genome-editing methods such as CRISPR/Cas9 with advanced genomic approaches (GWAS, GS) offers unique opportunities for crop resistance improvement. Although research on CRISPR/Cas9 applications for enhancing oat resistance to B. graminis f. sp. avenae remains at an early stage, results from other cereals indicate substantial potential. Continuous progress in bioinformatics, sequencing technologies, and post-editing plant regeneration methods enables increasingly precise and efficient modification of resistance-related genes. Consequently, this field is expected to expand to oats in the coming years, opening new perspectives for breeders seeking to enhance powdery mildew and fungal disease resistance in this species. The integration of monitoring strategies and molecular technologies discussed above must be considered within the broader context of oat biology and pathogen ecology to ensure effective resistance breeding programs.
. Summary
Plant diseases are highly dependent on environmental changes and agricultural practices (Chakraborty et al., 2000). The successful adaptation of crops, such as oats (Avena sativa), requires a deep understanding of the molecular mechanisms and pathways involved in pathogen resistance (Ziska et al., 2012).
Although genetic modeling has become easier thanks to modern techniques, a real challenge is the lack of comprehensive knowledge of the oat genome. Phylogenetic analysis shows that oats are positioned to highlight their genetic differences from other cereal pathogens. This suggests that the immunity mechanisms for biotic stress described in barley and wheat may not be entirely the same as those observed in oats (Inuma et al., 2007).
Additionally, the position of oats on the phylogenetic tree suggests a potential exchange of pathogens with other grasses, making it particularly susceptible to infections from various B. graminis strains (Inuma et al., 2007). These challenges, combined with the growing importance of oats in human nutrition, underscore the critical need for continued research in this field.
. Conclusion
The information regarding the specificity and evolution of B. graminis, as well as the limited knowledge of oat (Avena sativa) genetics and resistance, highlights the crucial importance of research on plant defense mechanisms, with a particular focus on oats. Due to its proven health benefits and increasing popularity as a highly nutritious cereal, agricultural producers are increasingly choosing to cultivate it (McCartney et al., 2011; Sanz et al., 2013).
Unfortunately, the profitability of oat cultivation is often undermined by significant yield losses caused by powdery mildew (B. graminis f. sp. avenae) infections. Therefore, it is essential to further expand the knowledge of oat defense mechanisms, build upon existing data, and develop new technologies to enhance its resistance.
Moreover, research on oat resistance could not only provide solutions for this species but also open opportunities for applying biotechnology to transfer oat defense mechanisms to other crops. Such a strategy could significantly improve global plant resistance to biotic stress while promoting sustainable agriculture and strengthening food security. Given these factors, research on oat resistance should be considered a high priority.
