. Introduction
The family Amaryllidaceae s. l. is represented by highly ornamental plants that are commonly found on all continents, except Antarctica (Takhtajan, 2009). Many representatives are found in tropical and subtropical zones and less often in temperate climates. Most genera grow in Central and South America, Southern Africa, and the Mediterranean. Amaryllidaceae are perennial herbaceous plants that can grow up to 2 m in height. The main feature of this family is the presence of subterranean or aerial bulbs. The flowers are gathered in umbels and sometimes cymes. There can sometimes be a reduction in inflorescences and the formation of single flowers (Meerow & Snijman, 1998). The family was established in 1805 and now has approximately 1,600 species and approximately 75 genera, 17 tribes, and three subfamilies: Agapanthoideae Endlicher, Allioideae Herb., and Amaryllidoideae Burnett (The Angiosperm Phylogeny Group, 2016). The genera Clivia Lindl. (six species) and Scadoxus Raf. (9–12 species) belong to the tribe Haemantheae of the subfamily Amaryllidoideae, which has six genera (Stevens, 2001).
Representatives of the genus Clivia have a large number of varieties, the study of which is gaining popularity (Petravich et al., 2015). Clivia is also the study subject of frequent fungal diseases in tropical plants (Li et al., 2020; Liu et al., 2020). The toxicity of C. miniata fruits and seeds has been studied (Kiepiel & Johnson, 2019), as well as the genes involved in the formation of Clivia leaves (Qin-Mei et al., 2018; Wang et al., 2016). Changes in the pollinator community of Scadoxus cinnabarinus along its altitude belt on Mount Cameroon have been studied by a group of Czech and African scientists (Mertens et al., 2020).
Using sequences from the plastid region trnL-F and internal transcribed spacer (ITS) nrDNA, Meerow and Clayton (2004) investigated the phylogeny of the African tribe Haemantheae on 19 species of all genera of the tribe. ITS and combined matrix parsimony analysis generated a well-maintained tree. Two main clades were detected. One included the monophyletic rhizomatous genera Clivia and Cryptostephanus. A larger clade combines Haemanthus and Scadoxus as a sister group to the Apodolirion-Gethyllis subclade (Meerow & Clayton, 2004).
Flowering efficiency, increasing the growing degree days required for anthesis time, and the effect of cold storage (12 °C for 4 weeks) before forcing were evaluated in Scadoxus multiflorus subsp. katharinae (Baker) Friis & Nordal. (Funnell, 2008). The results of butterfly-wing pollination in two subspecies of S. multiflorus and a review of the prevalence of this pollination mechanism among other Amaryllidaceae in Southern Africa was recently published (Butler & Johnson, 2020). Karyotypic features include organizations of the 5S and 45S rDNA loci, telomeres of S. multiflorus (Monkheang et al., 2016), and tracking biological footprints of climate change using the flowering phenology of the geophytes in Pancratium tenuifolium and S. multiflorus (Kwembeya, 2021). The effects of light duration on flower development in S. multiflorus have been described and evaluated (Vendrame et al., 2004).
Haemantheae is distinct from the other Amaryllidaceae in its rhizomatous habit, baccate fruit, and phytomelan-free seeds (Meerow & Snijman, 1998). Precise examination of the gynoecium with the inferior ovary and septal nectaries in the members of the tribe has not been performed. However, the gynoecium structure in monocots has been actively assessed with regard to the evolution of epigyny, loss of septal nectaries, and pollination mechanisms (Remizova et al., 2010; Rudall, 2002). Amaryllidoideae is the only clade within the higher Asparagales in which the inferior ovary occurs (Rudall, 2002).
Species of Clivia are rhizomatous, evergreen plants that naturally grow in Southern Africa. Representatives of the genus Clivia have actinomorphic or weakly zygomorphic flowers, narrowly to widely funnelform, have a pendulous spread, and a short floral tube. The filaments are free, and the stigma is tricuspidate. There are four-to-five ovules per locule. The berry is subglobose and red. The seeds are turgid and ivory-colored (Meerow & Snijman, 1998). Clivia miniata has an erect, subterranean, or shortly aerial rhizome, with thick fleshy roots. The umbel has 16 to 30 flowers raised above the leaves. The stamens are weakly to strongly declinate. The fruit is a baccate capsule with few seeds growing from the inside of the capsule (Duncan, 2017). Clivia nobilis has a pendent umbel of 20 to 50 flowers raised below the leaf tips (Duncan, 2017).
The genus Scadoxus is characterized by actinomorphic flowers, funnelform to salverform, with perigonium segments equal to or longer than the tube. The stamens are free, and the stigma is undivided. There are just a few ovules per locule. The berry is ovoid to globose and red. The seeds are ovoid and ivory-colored (Meerow & Snijman, 1998). Scadoxus multiflorus is a deciduous, summer-growing plant with rhizomatous bulbs, which grows naturally in sub-Saharan Africa, Arabian Peninsula, and Seychelles (Duncan, 2017). Umbels are many-flowered and spherical, and leaf blades are ovate, erect, and shiny, with two or three reduced, sheathing basal leaves.
The aim of our study was to conduct a comparative morphological analysis using micromorphological and anatomical characteristics of the flower in representatives of the tribe Haemantheae with regard to gynoecium-specific features.
. Material and Methods
Three species were selected for this study: Clivia miniata (Lindl.) Bosse, Clivia nobilis Lindl., and Scadoxus multiflorus (Martyn) Raf. (Figure 1). Buds and flowers were sampled in the A. V. Fomin Botanical Garden of Taras Shevchenko National University of Kyiv. The samples were fixed in 70% alcohol in April 2020. Five flower buds of each species that were 1.0 to 1.7 cm in length were dehydrated in an increasing series of t-butanol (20%, 30%, 50%, 70%, 100%; 2 hr each for 20%–70%, 24 hr for 100%). Each sample was stored in a 1:1 preparation of 100% t-butanol and Paraplast. Infiltration with Paraplast (Merck; New York, NY, USA) was performed as previously described (Baryikina et al., 2004). Transverse sections 20 µm in thickness were obtained with an MPS-2 manual rotary microtome (Tochmedpribor; Kharkiv, USSR) using an existing staining protocol (Soukup & Tylová, 2019) with safranin (Sigma-Aldrich; St. Louis, MO, USA) and Astra Blue (Merck). Slides were mounted in Eukitt (Sigma-Aldrich). Light microscopy was used, and images with 3,584 × 2,748 resolution saved as .jpg files were obtained with a 10MP digital camera attached to a modelT490B-10M microscope (AmScope; https://amscope.com/). Photopea software (https://www.photopea.com/) was used for image processing.
Figure 1
Flowering scapes of Clivia miniata (Lindl.) Bosse. (A), Clivia nobilis Lindl. (B), and Scadoxus multiflorus (Martyn) Raf. (C).

For morphological analysis, measurements were performed on at least 15 fresh flowers. To analyze the internal structure of the gynoecium, we used Leinfellner’s (1950) concept of vertical zonation. According to this concept, the congenital multilocular synascidiate, unilocular symplicate, transitional hemisymplicate, and asymplicate (apocarpous) zones can be formed in the syncarpous gynoecium. In the conditions of incomplete fusion of carpels only in their outer part, a hemisyncarpous gynoecium with hemisynascidiate, hemisymplicate, and asymplicate zones arises. This concept was subsequently developed for monocots with epigynous and hypogynous flowers regarding the septal nectary structure (Novikoff & Odintsova, 2008; Odintsova, 2013). The height of the gynoecium zones was measured according to the number of cross-sections.
. Results
. Clivia miniata
The flowers of C. miniata were 8.3 cm long (Figure 2B). The scape was 35–40 cm in diameter, 1.5 cm in diameter at the base, and 1.3 cm above. There were 10–13 flowers in an inflorescence. Bracts were cone-shaped, 3.2–3.6 cm long, 1.7–2.0 cm wide, leathery, and light brown. The pedicel was 3.2 cm long and 0.4 cm in diameter. The perigonium was slightly zygomorphic and bright orange. The tepals were fused into a short floral tube, 0.7 cm long and 0.7 cm in diameter. The outer tepals were 6.0–6.2 cm long and 1.9–2.1 cm wide. The tepals of the inner whorl were 6.4–6.5 cm long and 2.7–2.9 cm wide (Figure 2A). The stamens were adnate to the floral tube at the base (Figure 2A). The length of the stamen filaments of the outer and inner whorl was 4.1–4.2 cm and 4.6 cm, respectively. The filaments became narrower at the top. Anthers were introse linear, 0.4 cm long and 0.1 cm wide, and were connected to filament below the middle (Figure 2D). The style was slightly zygomorphic. The inferior ovary was obovate, 1.4 cm high, and 0.7 cm in diameter (Figure 3A). The lower part of the ovary was green, and the upper part was yellow (Figure 2C). The style was located in the center of the ovary. The style was filamentous, curved, 5.8 cm long, 0.2 cm in diameter at the base, and yellow. The stigma was light yellow and three-lobed with short lobes.
Figure 2
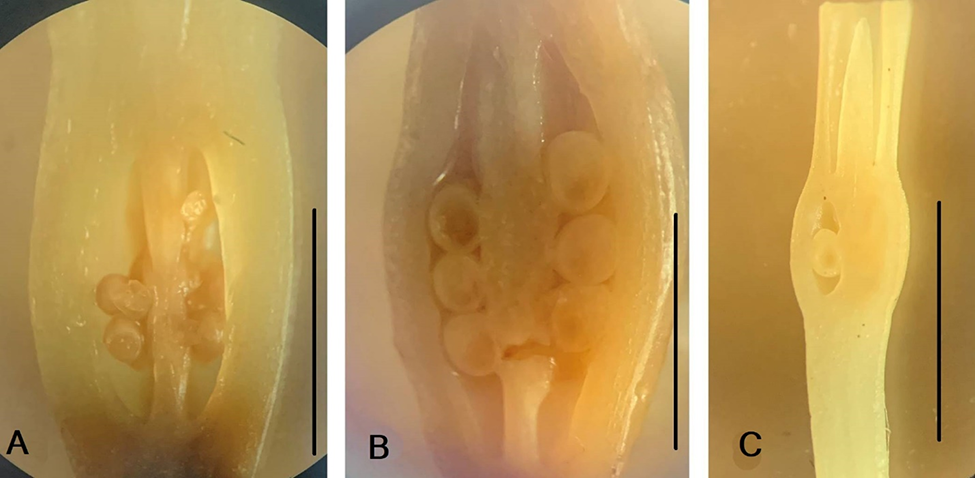
Anthetic flowers and flower details in members of the tribe Haemantheae. (A–D) Clivia miniata. (A) Flower, general view; (B) pistil; (C) series of tepals with connected stamens. (D) Preparated perigonium and androecium. (E,F) Clivia nobilis. (E) Flower; (F) pistil. (G–I) Scadoxus multiflorus. (G) Flower in the longitudinal section at late anthesis. (H) Fragment of the floral tube with stamens. (I) Pistil. Scale bars: 10 mm.

Figure 3
Longitudinal section of the inferior ovary in members of the tribe Haemantheae. (A) Clivia miniate; (B) Clivia nobilis; (C) Scadoxus multiflorus. Ovules are visible. Scale bars: (A) 10 mm; (B,C) 5 mm.
The ovary wall was thick (approximately 0.2 cm), as well as the ovary base and ovary roof, with five-to-six ovules in a locule (Figure 3A). In the C. miniata ovary, the following structural zones could be discerned (Table 1): a fertile synascidiate structural zone, fertile symplicate structural zone, and hemisymplicate zone (Figure 4). In the subapical portion, the locules became gradually rounded in cross section. They merged into a single channel within the ovary roof, which entered the center of the style as a style channel (Figure 5F). Septal nectaries appeared at the level of the hemisymplicate zone (Figure 5E, Figure 6B) and opened with slits that had no secretory tissue at the base of the style (Figure 6D). The cavities of the septal nectaries were oval in cross section and characterized by the absence of secretory tissue in the upper part. The asymplicate zone formed a style above the opening of the septal nectary (Figure 6D).
Table 1
Micromorphological and anatomical characters of the studied species of Haemantheae (30 individual plants).
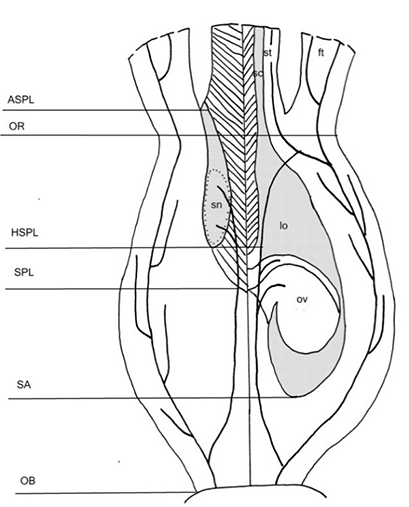
Figure 4
Gynoecium structure in Haemantheae (generalized view). The left side of the drawing represents the radial section in the septum plane, and the right one in the locule plane. Vascular bundles are shown. OB – ovary base; SA – synascidiate zone; SPL – symplicate zone; HSPL – hemisymplicate zone; OR – ovary roof; ASPL – asymplicate zone; lo – locule; ft – floral tube; ov – ovule; sc – style channel; sn – septal nectary; st – style. The postgenitally fused areas are shown in hatched lines.
The anatomical characteristics of the studied species are presented in Table 1. All floral parts displayed a parenchymatous, homogenous histological structure, with the presence of idioblasts with raphides in the peduncle and ovary wall (Table 1). The inner epidermis of the ovary comprised small cells (Figure 5D).
The peduncle of C. miniata contained six large vascular bundles and six small vascular bundles (Figure 5A), from which three septal veins and three dorsal veins formed at the ovary base (Figure 5B). The other vascular bundles were concentrated closer to the center of the ovary. Every dorsal (Figure 5D) and septal vein (Figure 6A) became three-bundled and produced small horizontal branches in the ovary wall. In the upper part of the ovary, the small bundles branched from the dorsal vascular bundles, which extended into the inner layer of the ovary wall (Figure 5E).
In the central column of the ovary, on the septum radius, a massive vascular bundle comprised the roots of the ventral complex (Figure 5C). Distally, each root of the ventral complex is divided into two bundles (ventral bundles of the carpel) that extended to the ovules. The small branches that supply the septal nectaries formed from the ventral bundles. Above the locules, the ventral bundles of the carpel, as well as the septal bundles and other small veins, fused with the dorsal bundles and entered the style. In the floral tube, the dorsal and septal veins were divided, and their branches entered the stamen and tepal traces (Figure 5F). Tepal traces branched higher in the floral tube, initially into three bundles and, higher, into a larger number of bundles (Table 1). The stamen traces were two-bundled.
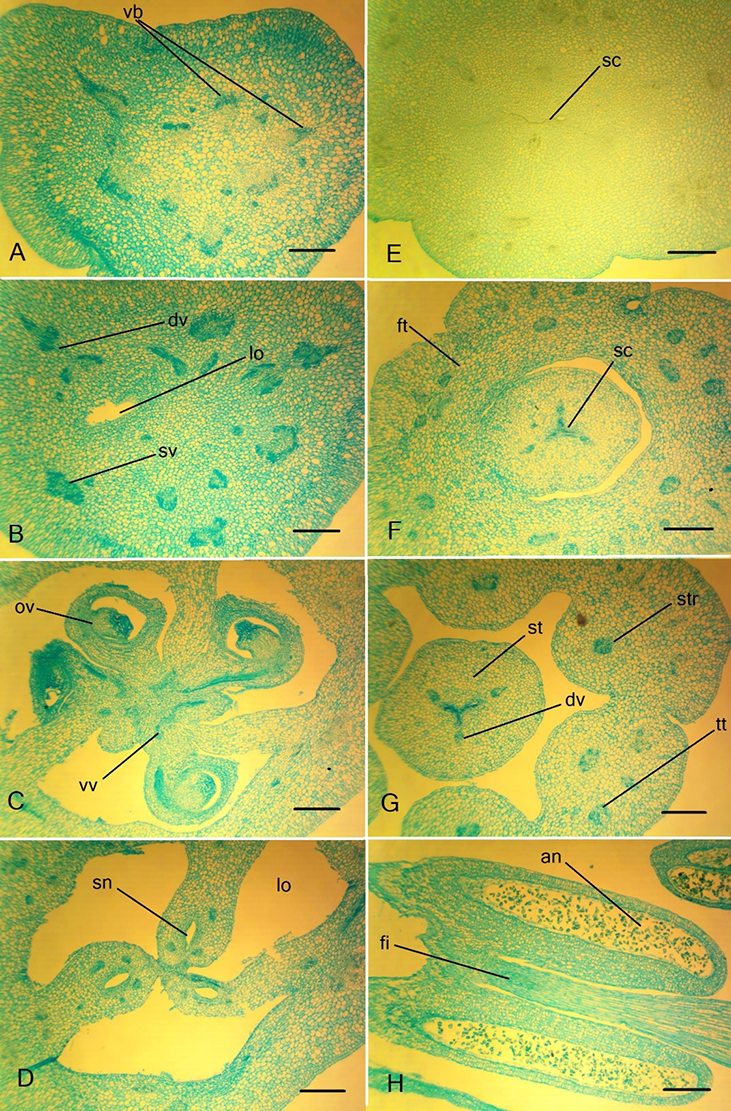
Figure 5
Ascending series of transversal sections of the Clivia miniata ovary. (A,B) Peduncle; (C–E) ovary; (F) ovary roof. dv – dorsal vein; lo – ovary locule; ov – ovule; rvc – roots of the ventral complex; sc – style channel; sn – septal nectary; str – stamen trace; sv – septal vein; vb – vascular bundle. (C) Synascidiate structural zone. (D) Symplicate structural zone. (E) Hemisymplicate structural zone. Scale bars: 500 µm.
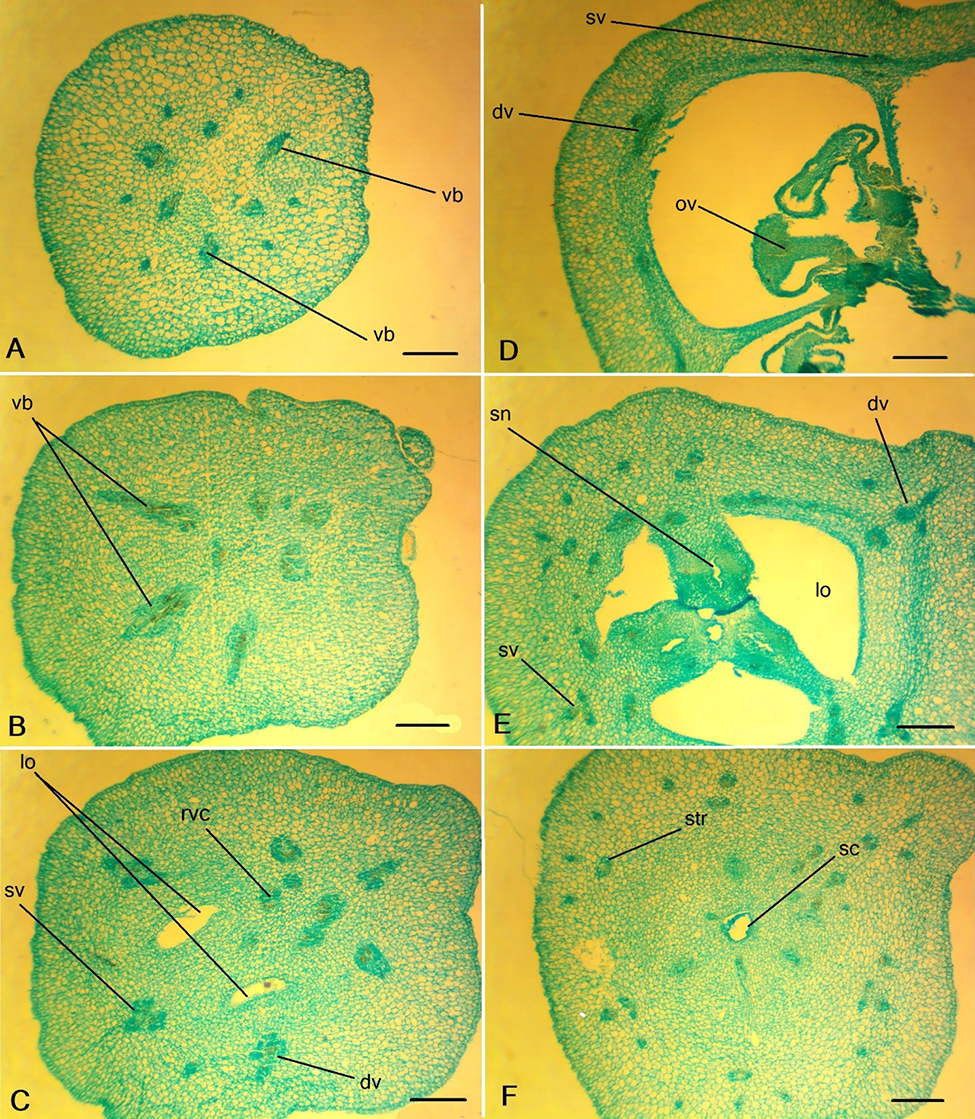
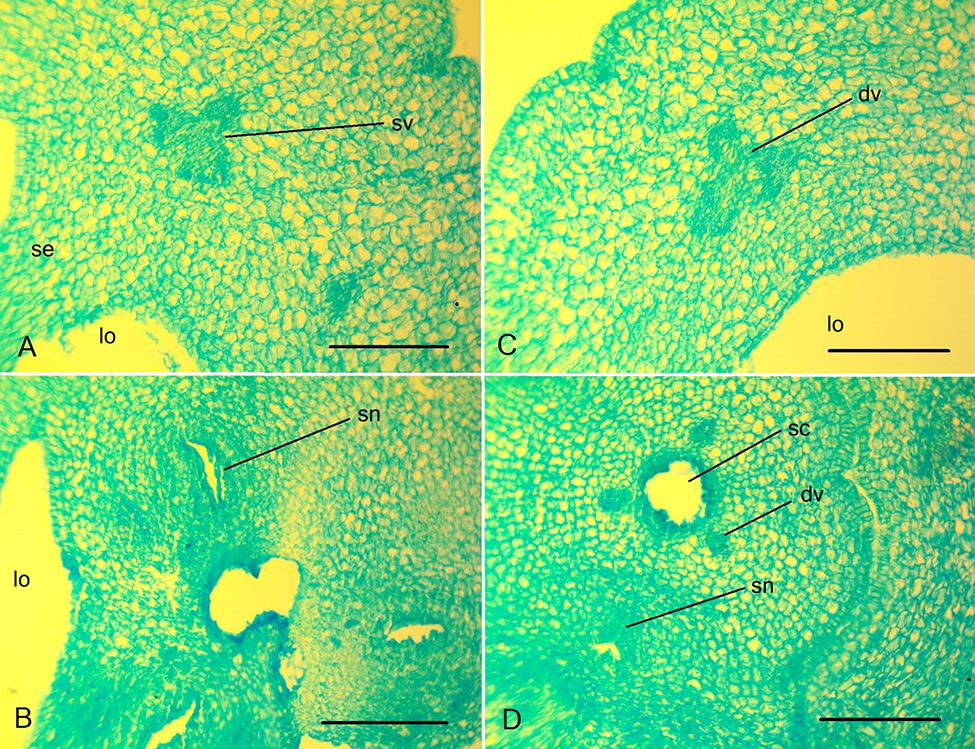
Figure 6
Details of the gynoecium structure of Clivia miniate. (A) Ovary wall with septa attached; (B) the central part of the ovary with three septal nectaries; (C) ovary wall in the median plane; (D) style base with triradial style channel. dv – dorsal vein; lo – ovary locule; sc – style channel; se – septa; sn – septal nectary; sv – septal vein. Scale bars: 250 µm.
. Clivia nobilis
The flowers of C. nobilis were 6.9 cm long and 1.0–1.1 cm in diameter (Figure 1B). The scape was 36–38 cm, 0.9 cm at the base, with 15–17 flowers in the inflorescence. The pedicel was 0.9–1.0 cm long and 0.2 cm in diameter. The perigonium was slightly zygomorphic and bright red-yellow with green tips of the tepals (Figure 2F). The tepals fused into a short floral tube (Figure 7F,G) that was 1.3 cm long and 0.8 cm in diameter. The outer tepals were 4.1–4.6 cm long and 0.9 cm wide. The inner tepals were 4.4 cm long and 1.4–1.5 cm wide. The stamens were adnate to the floral tube. The length of the stamen filaments of the outer whorl was 3.3–3.5 cm, and the stamen filaments of the inner whorl were 3.7–3.8 cm. The filaments were 0.1 cm in diameter in the outer stamens and 0.15 cm in the inner stamens and narrowed towards the top. The anthers were linear, 0.5 cm long, and 0.1 cm wide and connected to a filament below the middle (Figure 7H).
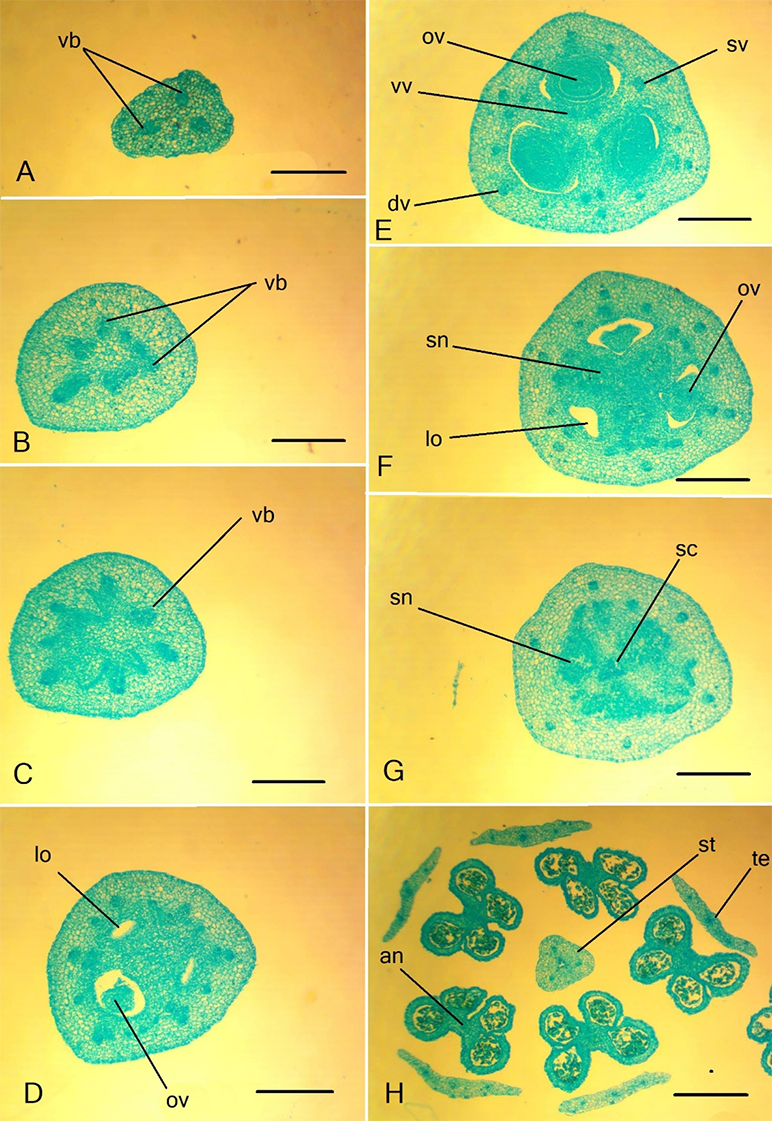
Figure 7
Ascending series of transversal sections of the flower details of Clivia nobilis. (A) Peduncle; (B–D) ovary; (E,F) ovary roof; (G) floral tube and style; (H) anther in the longitudinal section. an – anther; dv – dorsal vein; fi – filament; ft – floral tube; lo – ovary locule; ov – ovule; sc – style channel; sn – septal nectary; st – style; str – stamen traces; tt – tepal traces; sv – seplal vein; vb – vascular bundle; vv – ventral bundle. Scale bars: 500 µm.
The gynoecium was slightly zygomorphic (Figure 2F). The ovary was obovate, 0.6 cm in diameter, and 0.8–1.0 cm in height (Figure 3B). The lower part of the ovary was green, and the upper part was yellow. The style was located in the center of the ovary. The style was filamentous, yellow, curved in the middle, 5.6 cm long, 0.1 cm in diameter at the base. The stigma was light yellow and three-lobed. There were six-to-eight ovules per locule (Figure 3B, Figure 7C, Table 1). The C. nobilis gynoecium contained the same structural zones as in C. miniata. They were the same height, except for the symplicate zone (Figure 4), which is 3.5 times shorter (Table 1). The septal nectary (Figure 7D, Figure 8B) was similarly located in the hemisymplicate region of the ovary (Figure 4). The style channels formed apically (Figure 7E,F, Figure 8D). There were idioblasts with raphides in the peduncle and ovary wall in C. nobilis (Table 1).
The structure of the vascular system of the flower was similar to that of C. miniata, with some minor differences (Table 1). The C. nobilis peduncle contained 15 vascular bundles (Figure 7A). At the level of the ovary base, septal bundles (Figure 8C) and dorsal bundles departed (Figure 8A). Small vascular bundles were located proximally to them (Figure 7B–D). In the floral tube, the dorsal and septal veins were divided and their branches entered the stamen and tepal traces (Figure 7F). Small bundles located inside the ring of dorsal and septal bundles anastomosed with each other and formed roots of the ventral complex. The bundles of the ventral complex branched into paired ventral bundles of carpels, supplying ovules and septal nectaries.
Figure 8
Details of the gynoecium structure of Clivia nobilis. (A) Ovary wall in the median part of the carpel, dorsal vein. (B) The central part of the ovary with three septal nectaries. (C) Ovary wall with septa attached; septal vein composed of three vascular bundles is visible. (D) Style base. dv – dorsal vein; lo – ovary locule; ns – nectary slit; ov – ovule; sc – style channel; se – septa; sn – septal nectary; sv – septal vein; vv – ventral vascular bundles. Scale bars: 250 µm.

. Scadoxus multiflorus
The flower of S. multiflorus was up to 4.2–4.5 cm long (Figure 1C, Figure 2G,H). The scape was 35–41 cm long and 0.2 cm in diameter. The five bracts were 5.2–5.8 cm long, 1.0–1.6 cm wide, and narrowed at the base. The pedicel was 3.8–4.5 cm long and 0.2 cm in diameter. The perigonium formed a floral tube 0.9–1.1 cm long and 0.3 cm in diameter (Figure 2G). The outer tepals were 2.6 cm long and 0.2 cm wide. The inner tepals were 2.3 cm long and 0.1 cm wide. The floral tubes and tepals were bright red (Figure 2H). The stamens were adnate to the floral tube at the base (Figure 2H). The stamens of the outer whorl were slightly longer than those of the inner whorl. The filaments were acuminated and red. The anthers were linear. On the outer stamens, anthers were 0.2 cm long. On the inner stamens, anthers were 0.15 cm long and 0.05 cm in diameter. The anthers were attached to the filament below the middle (Figure 9H). The ovary was oval, light green, 0.3 cm in height, and 0.25 cm in diameter, with a single ovule in each locule (Figure 3C). The style was filamentous (Figure 2I), white at the base, red at the top, had a central location, and 3.6 cm high. The stigma was capitate.
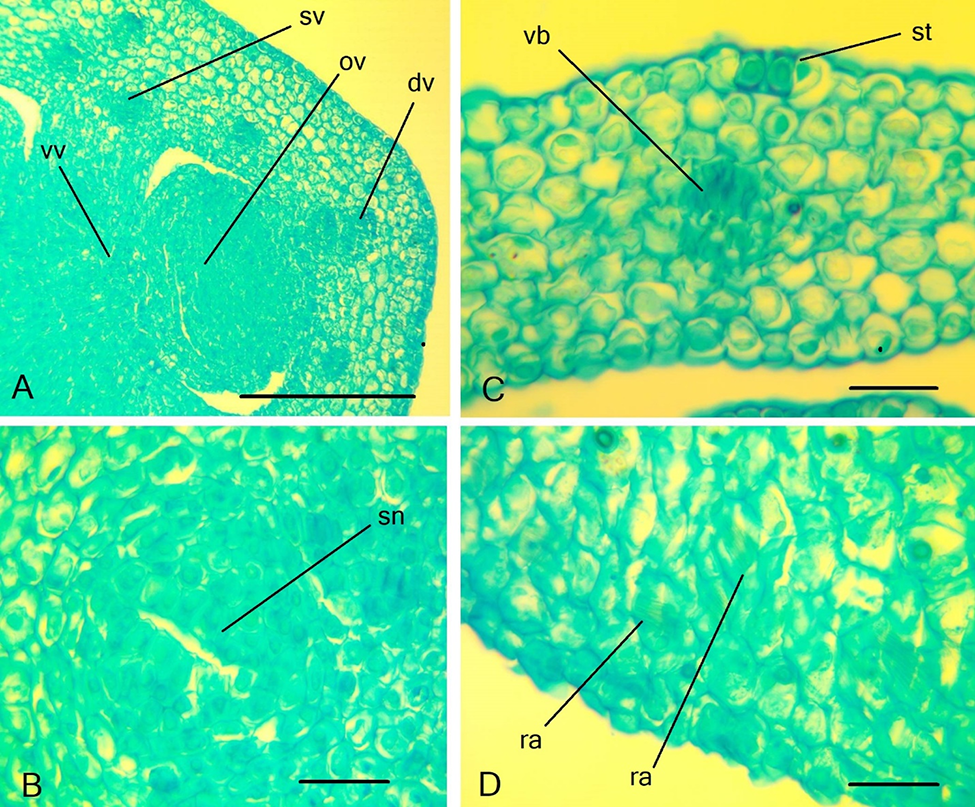
Figure 9
Ascending series of transversal sections of Scadoxus multiflorus flower. (A–C) Peduncle; (D–F) ovary; (G) ovary roof; (H) floral details at anther level. an – anther; dv – dorsal vein; lo – ovary locule; ov – ovule; sc – style channel; sn – septal nectary; st – style; te – tepal; sv – septal vein; vv – ventral vascular bundles; vb – vascular bundle. Scale bars: 500 µm.
The ovary had the same vertical zones, and the septal nectary was located in the hemisymplicate zone. The ovary base and roof were slightly thickened, but the ovary wall was thinner than that in Clivia species (Figure 3, Figure 9E,F, Figure 10A,C, Table 1). The septal nectaries were very narrow and barely visible in the cross section (Figure 9G, Figure 10B). The presence of a large number of idioblasts with raphides in all organs of the flower and stomata on the tepals was peculiar to the species (Figure 10C,D, Table 1).
In the peduncle of S. multiflorus, only three vascular bundles were available at the base (Figure 9A,B), which branched distally into three bundles each. At the level of the ovary base, nine vascular bundles were present (Figure 9C), from which septal bundles, dorsal bundles, and small vascular bundles were proximal to septal bundles and dorsal bundles departed (Figure 9D). Higher in the ovary in each septum, small vascular bundles were present as the roots of a ventral complex (Figure 9D). They formed paired ventral vascular bundles (Figure 9E,F). Each ovule was supplied by two bundles extending from the ventral bundles of the carpels. The dorsal veins of carpels were two-bundled (Figure 10A), and septal veins were single-bundled.
Figure 10
Details of the gynoecium structure of Scadoxus multiflorus. (A) Ovary wall at the level of ovule insertion; (B) septal nectaries; (C) stoma on the tepals; (D) raphides in the distal parenchyma of the ovary wall. dv – dorsal vein; ov – ovule; ra – raphides; sn – septal nectary; st –stoma; sv – septal vein; vb – vascular bundle; vv – ventral vein. Scale bars: (A) 250 µm; (B–D) 50 µm.
. Discussion
The flowers of the studied species have many common features, including a large size, slightly zygomorphic, short-tubular, and red-orange color, with the same set of organs. However, the functional type of flower in Clivia species is tubular, broadly tubular to disc-shaped; in S. multiflorus, the attractiveness is ensured by narrow, linear tepals, and colored elongated filaments (Leins & Erbar, 2010).
Understanding the evolution of flower morphology requires information about the identity of pollinators and knowledge about specific mechanisms of pollen transfer (e.g., Bożek, 2019). Based on preliminary field observations and flower structure, Butler and Johnson (2020) hypothesized that pollination mechanisms involving the transfer of pollen on butterfly wings occur in several genera of South African Amaryllidaceae, including S. multiflorus.
In the gynoecium of the studied species, we found the same vertical zonality, characteristic of eusyncarpous gynoecium, that is, the presence of synascidiate, symplicate, hemisymplicate, and asymplicate zones. The synascidiate zone was well developed in the studied species, with a central column and placenta. The symplicate zone was longer or shorter and contained the placenta in the Clivia species. The hemisymplicate zone in the studied species was sterile, occupying the upper half of the ovary. The main function of this zone was the formation of septal nectaries. In S. multiflorus, the size of all zones of the ovary was the smallest, as was the size of the whole ovary. However, in this species, the roof of the ovary was the thickest. Thus, in the studied species, the seed-bearing and nectar-bearing parts of the ovary were separated in space. In C. miniata and S. multiflorus, the style was slightly longer than that of the stamens, which provides approach herkogamy. In Clivia, the style also curved. Clivia flowers have nectary guides on the tepals, and the pollinator is rewarded with nectar from the septal nectaries. We have previously studied synascidiate and symplicate zones in Galanthus nivalis and Leucojum vernum gynoecium (Fishchuk & Odintsova, 2020). We found that in the Hippeastrum striatum gynoecium presented synascidiate, symplicate, hemisymplicate, and asymplicate vertical zones (Fishchuk, 2021).
As revealed in this study, the septal nectary opened at the base of the style so that the style formed by the postgenital fusion of the carpels were above the output of the nectary slits. The nectary is represented by three narrow cavities, which connect in the center of the ovary by a common epidermis, although they are postgenitally closed. The septal nectary in the studied species had different lengths. It was shortest in S. multiflorus and approximately the same in C. miniata and C. nobilis. The septal nectary portion in the ovary was the largest in S. multiflorus, where it was more than half the height of the ovary. This nectary structure corresponds to the “lilioid” distinct nonlabyrinthine type, according to Schmid (1985). This structure of the septal nectary is characteristic of other species of the order Asparagales, which have an inferior ovary and septal nectary, such as Gladiolus hybridus from Iridaceae (Skrypets′ & Odintsova, 2013). The nectaries are in the form of a nectary disk located between the stamens and the style of G. nivalis. The septal nectary sutures are absent, but in the center of the ovary in the simplicate zone, a triradial cavity forms between the ends of incomplete septa in Leucojum vernum. (Fishchuk & Odintsova, 2020). Hippeastrum striatum has internal septal nectaries that appear at the ovary base and are continued by nectar fissures (Daumann, 1970). In the genus Allium (Amaryllidaceae), nectar is secreted at the top of the ovary, where there is a greater risk of theft and less contact with the fertile part by pollinating visitors. Gynopleural nectaries are also absent from some genera of Amaryllidaceae. In Sternbergia (Amaryllidaceae), tepalar and staminal nectaries occur in addition to gynopleural nectaries (Smets et al., 2000).
According to Daumann (1970), the genera Clivia and Scadoxus have an internal septal nectary with slits in the ovary septa, characterized by the absence of vascular bundles that supply the nectary. Nectaries of the genus Clivia are composed of three septal nectary slits, which mostly extend above the locules, which appear only in the upper two thirds of the ovary height. The cells of the nectary epidermis are significantly elongated (in the form of a palisade). Nectary excretion occurs only in the lower two thirds of the septum, where the cells of the epidermis of the nectary are elongated. Towards the style, they gradually become shorter. At these points, nectary secretion becomes weaker and finally stops completely, so that (although without a sharp limit) approximately the upper one third of the nectary slit provides only the exit channel for nectary excretion, released by the lower parts of the nectary. Prominent groups of vascular bundles pass close to the nectary parenchyma but provide no branches into the nectary tissue (Daumann, 1970).
Examining other members of the order Asparagales, we found similar features of thickened ovary base and ovary roof, and the presence of all zones in the gynoecium in the family Asparagaceae s. l. (Fishchuk & Odintsova, 2013, 2014; Odintsova & Fishchuk, 2017). Most of the studied species also have a septal nectary, but they are usually prolonged from the ovary base to the ovary roof.
Despite the obvious adaptations to the attraction of pollinators, a low number of ovules are formed in the ovaries of the studied species, which is characteristic of indehiscent fruits. A solitary ovule occurs in each locule of S. multiflorus. In the H. striatum ovary, there are 16–20 ovules in each locule (Fishchuk, 2021), in G. nivalis there are 14 ovules, and in L. vernum there are 12–14 ovules in each locule (Fishchuk & Odintsova, 2020). The other traits, which are distinct in S. multiflorus compared with Clivia species, include abundant raphides in floral parts, three bundles in tepal traces and two bundles in ovule trace, and fewer cell layers in the ovary wall. Idioblasts with raphides are present in H. striatum in the upper part of the peduncle, receptacle, stamen filaments, ovary wall, and free tepals (Figure 4). They are absent in the connective style (Fishchuk, 2021). The upper part of the pedicel, base of the floral tube, stamen filaments, connectives, the wall of the ovary, and the style contain idioblasts with raphides in G. nivalis, and in the free apices of the tepals, they are absent. At the base of the floral tube, the stamen filaments, and the wall of the ovary, free apices of the tepals contain idioblasts with raphides in L. vernum. They are absent in the apical part of the pedicel and the style (Fishchuk & Odintsova, 2020). In plant cells, idioblasts with raphides can perform protective, supportive, and spare functions.
Comparing the structure of the flower with representatives of the tribe Galantheae (Amaryllidaceae) – G. nivalis and L. vernum (Fishchuk & Odintsova, 2020) – several differences were evident. In Haemantheae, anthers are attached to the floral tube and open along their entire length, a septal nectary is present, there is no aerenchyma in the ovary wall and other parts of the flower, and there are few ovules. In the tribe Galantheae, dehiscence of the anthers through the apical pores in Galanthus and Leucojum could be considered an extremely specialized variant of lengthwise splitting of short sutures in the apical part of the anthers, which is possibly attributed to the pendant position of flowers (Fishchuk & Odintsova, 2020). Common features are thick fleshy ovary walls and septa, multibundle traces of tepals, and the presence of synascidiate and symplicate zones.
The morphological and anatomical features of the flower in Asparagales are being studied to explain the appearance of some features of the flower and fruit (Odintsova & Fishchuk, 2017; Skrypec & Odintsova, 2020). One such feature is the organization of the flower vascular system. In the studied species, a large number of small vascular bundles were observed in the inferior ovary wall, which is an adaptation to the formation of fleshy fruit. The other trait characteristic of the fleshy fruit is a thickened ovary wall, base, and roof. The flower vascular system is characterized by three-bundled dorsal and septal veins and the presence of branches of the dorsal and septal veins in the ovary in C. miniata and S. multiflorus. In S. multiflorus, the dorsal bundles are doubled, and the septal bundles are single-bundled. In C. nobilis, the dorsal and septal bundles are three-bundled but have no branches. The present study of morphological and anatomical features of the flower will be useful in the taxonomical revision of the family Amaryllidaceae.
. Conclusions
We studied the micromorphology and anatomy of the flower C. miniata (Lindl.) Bosse, C. nobilis Lindl., and S. multiflorus (Martyn) Raf. The examination revealed common features in the gynoecium micromorphology and vascular anatomy of these species. Synascidiate, symplicate, and hemisymplicate zones were found in the inferior ovary, while the style was formed by an asymplicate zone. Ovules were located in the synascidiate and symplicate zones in C. miniata, C. nobilis, and in the synascidiate zone of S. multiflorus. The septal nectary is located in the hemisymplicate zone in the upper part of the ovary. The vascular system in the inferior ovary wall consisted of a circle of ascending bundles on the tepal radii, from which traces of stamens and tepals were formed. In the inner layer of the ovary wall, dorsal and septal veins were anastomosed with each other. Traces of ovules were formed from the ventral bundles of the carpel in the central column of the ovary. The studied species showed great similarity in their micromorphology and vascular anatomy.
