. Introduction
The Śnieżnik Massif is one of the several regions in the Sudetes with the highest diversity of vascular flora. Therefore, it was the subject of intensive floristic research already at the end of the 19th and the beginning of the 20th century (Fiek, 1881; Hruby, 1915, 1923; Limpricht, 1942; Schube, 1903; Zeiske, 1902). Their results were then summarized by Limpricht in the late 1940s (Limpricht, 1949). Due to changes in national borders after the Second World War, this region was not explored as intensively as before. Therefore, a unique colony of dealpine plants, with the participation of species that had been known in Poland only from the Carpathians, was reported from here rather late, first by Jeník (1961) and then by Fabiszewski (1970). Both researchers observed a rocky grassland community with Sesleria tatrae, Scabiosa lucida, and Galium anisophyllon on an isolated outcrop located on the east-facing slopes above the Kleśnica Valley. These species had never before been recorded from the Śnieżnik Massif. Moreover, it is still the only location of these plants in the entire Polish Sudetes. This important discovery raised widespread interest among professionals and botany enthusiasts, and it continues to this day. However, it is mainly seen on social media. References in literature to this subject are rare. Szeląg (2000), in a comprehensive analysis of the flora of the Polish part of the Śnieżnik Massif, emphasized the unique character of this site and listed other species occurring only here in the Polish Eastern Sudetes (some of them also occur in the Czech part of this range), such as Allium lusitanicum, Cotoneaster integerrimus, Euphrasia micrantha, Hieracium bifidum, Festuca rupicola, and the extremely rare here, Epipactis atrorubens. He also confirmed the presence of Scabiosa lucida, whose extinction in the Polish part of the Sudetes had already been reported by Brej et al. (1997). Other literature data concern only floristic or karyological studies of selected species such as Epipactis atrorubens (Jakubska-Busse & Śliwiński, 2010), Cotoneaster integerrimus (Boratyński et al., 1999), Hieracium bifidum (Musiał et al., 2016) or Sesleria tatrae (Budzáková et al., 2014). Additionally, the rare basiphilous lichen Solorina saccata was also described from this site (Szczepańska & Kossowska, 2004).
Although more than fifty years have passed since this discovery, no one has attempted to describe the compositional and ecological changes that possibly may have occurred at this unique site and whether it still represents the phytocoenoses originally described. According to the recent studies, such changes in species composition and environmental conditions are expected due to both anthropogenic climate warming (Gottfried et al., 2012; Malhi et al., 2020; Rammig et al., 2010; Steinbauer et al., 2022; Zeidler et al., 2023), and intensive atmospheric nitrogen pollution, leading to an increase in the proportion of nitrophilous species within many of the plant communities studied (de Schrijver et al., 2011; Perring et al., 2018; Staude et al., 2020). The latter factor was already reported in the 1980s from the Śnieżnik Massif by Brej et al. (1997).
Many uncertainties surround the phytosociological identification of the described grassland community. Fabiszewski (1970) suggested that it could be an extremely impoverished form of the association of the alliance Seslerion tatrae Pawłowski 1935 corr. Klika 1955. His opinion was also confirmed by Budzáková et al. (2014), who conducted a phytosociological analysis of grasslands with Sesleria tatrae from the Carpathians, additionally including a relevè from the Pulinka. However, the Pulinka grasslands were not included in the Polish phytosociological literature (Matuszkiewicz, 2007) and in the manuals for the protection and monitoring of the Natura 2000 habitat “6170 Alpine and subalpine calcareous grasslands” in Poland (Kozak & Kozłowska, 2012; Perzanowska & Mróz, 2004). Therefore, the habitat 6170 was not included in the Standard Data Form for the Natura 2000 site PLH020016 “Góry Bialskie i Grupa Śnieżnika” (Świerkosz & Furmankiewicz, 2012), and was not protected in this area at the time of writing this article. Meanwhile, it may be the only site of habitat 6170 in the continental region of Poland with a unique contribution to the biodiversity of this part of the country. Consequently, in the pan-European synthesis of the distribution of syntaxonomic units at the alliance level (Preislerová et al., 2022), the Seslerion tatrae was not reported from the area of the Śnieżnik Massif.
The aims of the paper are, therefore: (i) to analyze the changes that took place in the community of the relic, basiphilous rocky grasslands with Sesleria tatrae in the years 1968–2022 at the site in the Kleśnica Valley in the Śnieżnik Massif, with particular emphasis on the species that, apart from the Pulinka, are only found in Poland, in the Carpathians; (ii) attempt to determine the syntaxonomic position of the studied rocky grassland community.
. Material and methods
. Study area
There are two mountain ranges in Poland that form the country’s southern border. The Carpathians, which are strongly diversified in terms of altitude and physiography, are part of a vast mountain range extending from Poland and Slovakia (and partly Czechia) through Ukraine to Romania. A long geological history of this mountain range, together with a weak influence of Pleistocene glaciations (Lindner & Grzybowski, 1982), have contributed to the development of a rich and diverse flora (Ronikier, 2011), with many endemic species and plant communities, especially in the alpine zone. The second mountain range, namely the Sudetes, forms the Polish-Czech border and belongs to the geomorphological province of the Bohemian Massif. In the Middle Pleistocene, the Sudetes were located closer to the front of the ice sheet, especially during the MIS-10 and MIS-6 (Marks et al., 2019). Therefore, their flora is marked by a higher proportion of post-glacial relics and a few non-endemic species (Kwiatkowski & Krahulec, 2016). On the other hand, limestone rocks are far less frequent here, hence the flora of the Sudetes is poorer in species as compared to the Carpathians. Therefore, in Poland, these two mountain ranges have the status of separate biogeographical regions, which differ considerably in terms of flora and plant communities (Matuszkiewicz, 2008).
The Śnieżnik Massif is located in the Eastern Sudetes Mts. in southwestern Poland. This region, like the whole of the Sudetes, is characterized by a complex geological structure, dominated by old metamorphic rocks: gneisses, with a smaller proportion of quartzites, amphibolites, schists, and – particularly important for the biodiversity of the area – limestones (crystalline marbles or dolomites) and serpentinites. The highest peak is Śnieżnik Kłodzki Mt. (1,425 m a.s.l.) from which six broad, flattened ridges extend, separated by deep valleys. One of these valleys is the valley of the Kleśnica River, a north-flowing watercourse that cuts through a large marble body at an altitude of ca. 890–820 m a.s.l. These marbles were mined in several quarries. The quarries are now closed, but on the steep slopes there are still some rocky walls or outcrops that serve as shelters for calcareous vegetation. One of them is a rock called Pulinka, whose presence, under the name of Haustein, was already marked on maps at the beginning of the 20th century. The Pulinka is a natural rock formation built of hardly weathered, coarse-crystalline metamorphosed Precambrian marble. It is located deep in the beech forest montane belt, at an altitude of 830–845 m a.s.l. The destruction of the rocks lying in the lower parts of the slopes, as a result of mining in the Kletno I and Kletno II quarries, does not allow us to determine whether other rocks with similar floristic characteristics had previously occurred below the Pulinka (Fabiszewski, 1970).
The climate of this region is of montane character, with high annual precipitation (800–1,000 mm) and low mean annual temperature (4–5 oC) (Głowicki et al., 2005). This combination of climatic factors has resulted in the formation of an upper timberline on the summit of Mt. Śnieżnik Kłodzki (ca. 1,250 m a.s.l.) and alpine communities (ass. Cetrario-Festucetum supinae Jeník (1961) and ass. Festuco supinae-Vaccinietum myrtilli Šmarda, 1950) above it (Krahulec, 1990; Szeląg, 2000). Well-preserved upper montane spruce forests (ass. Calamagrostio villosae-Piceetum abietis Schlüter 1966) cover almost 600 ha (Fabiszewski, 1968; Reczyńska & Świerkosz, 2013). At lower elevation, there are acidophilous and basiphilous beech forests (ass. Luzulo luzuloidis-Fagetum sylvaticae Meusel 1937 and ass. Hordelymo europaei-Fagetum sylvaticae Kuhn 1937), ravine forests (all. Tilio platyphylli-Acerion Klika 1955), tall-herb communities (cl. Mulgedio-Aconitetea Hadač et Klika in Klika et Hadač 1944), rocky communities (all. Ctenidio-Polypodion vulgaris S. Brullo et al. 2001), as well as many semi-natural plant communities (Świerkosz & Furmankiewicz, 2012). The Polish part of the Śnieżnik Massif is included in the Natura 2000 site PLH020016 “Góry Bialskie i Grupa Śnieżnika”.
. Field sampling and data analysis
The fieldwork was carried out in the summer of 2022. In order to document the current species composition of the rocky grassland community, five phytosociological relevès were collected in homogeneous vegetation plots according to the Braun-Blanquet method (Braun-Blanquet, 1964; Mueller-Dombois & Ellenberg, 2002). The area of the vegetation plots ranged from 4 to 16 m2. Additionally, two phytosociological relevès from 1968 (Fabiszewski, 1970) and one recorded in 2010 (Budzáková et al., 2014) were used in the analysis. The JUICE 7.01 software was used to organize the phytosociological table and to determine changes in species composition during different study periods (Tichý, 2002).
As only random data were available from 1968–2010, the only possible statistical analysis was a comparison of changes in community-weighted means (CWMs) calculated for each plot with reference to Ellenberg Indicator Values for Europe (EIVEs for temperature, light, nutrients, soil reaction and moisture) (Dengler et al., 2023) and selected functional traits: specific leaf area (SLA), seed mass (SM) and leaf dry matter content (LDMC). In the next step, the statistical significance of the differences between the medians of these values from each study period were tested using the Kruskal–Wallis test for equal medians, with full awareness of the small sample size.
Data on functional traits for individual species were mainly obtained from the LEDA database (Kleyer et al., 2008), and in the case of species not represented there, we used the E-Vojtkó et al. (2020) dataset. Of the 44 species reported in the phytosociological relevès, SM data were obtained for 43, SLA for 39, and LDMC for 37 species. CWMs for individual functional traits were calculated excluding species for which no data were available. Additionally, a functional diversity index (FD Rao) (Botta-Dukát, 2005) was calculated for each relevè. CWMs and FD Rao were calculated using Canoco 5.0 (ter Braak & Šmilauer, 2012), while significance tests were performed using PAST 4.03 software (Hammer et al., 2001).
As the response data were compositional and had a gradient 2.7 SD units long, a linear ordination method (Principal Component Analysis, PCA) was used in Canoco 5.0 software (ter Braak & Šmilauer, 2012) to identify the relationships between the species composition of vegetation plots documented in subsequent research periods and their relationship with the community-weighted means of selected EIVEs and functional traits.
The nomenclature of vascular plants and mosses was harmonized according to the World Flora Online Plant List (WFO. World Flora Online. Version 03.10.2023, 2023). The nomenclature of syntaxonomic units follows Mucina et al. (2016) and Preislerová et al. (2022) at the level of alliances, and with Chytrý (2007–2013) at the level of associations, except for Hordelymo europaei-Fagetum sylvatici (Świerkosz et al., 2018).
. Results
. The first locality of Calamagrostis varia in the Sudetes Mts.
During a field visit on 07.07.2022 Pavel Novák noticed unusual specimens of the genus Calamagrostis sp. resembling C. varia. He collected and identified these specimens, which confirmed the initial field diagnosis (rev. Milan Štech, University of South Bohemia). The herbarium specimen was deposited in the herbarium of the Department of Botany and Zoology of the Masaryk University, Brno (BRNU 685330). This species has not been reported from the Pulinka itself, nor has it been found so far in either the Polish (Zając & Zając, 2001) or Czech part of the Sudetes (Chytrý et al., 2021). C. varia was mainly found in basophilic rocky grasslands and calcicolous, nutrient-rich beech forest that developed on shallow, skeletal soils, just below the summit of the Pulinka. The population of C. varia was scattered over an area of about 500 m2 and co-occurred with C. arundinacea (Figure 1A–C).

. Changes in species composition of the rocky grassland community
The species composition of the studied grassland community (Figure 2A–B) is characterized by high variability over time, but this is related to individual taxa, which are usually found in low abundance. Among the more important changes is the absence of Asplenium viride and Thymus pulegioides recorded in 1968 (Fabiszewski, 1970) and nemoral species, some of which were still recorded in 2010, such as Daphne mezereum, Convallaria majalis, Epipactis helleborine and Lilium martagon (Table 1). Moreover, Budzáková et al. (2014) recorded in 2010 eight species: Crepis conyzifolia, Gentianopsis ciliata, Leontodon hispidus, Phleum pratense, Phyteuma spicatum, Pimpinella major, Veronica officinalis and Veratrum lobelianum, whose occurrence was not confirmed either in 1968 or 2022. Furthermore, species which were not recorded in 1968, became present in the relevès collected in 2010 and 2022. Some of these taxa such as Allium lusitanicum, Arabidopsis arenosa, Fagus sylvatica, Hieracium bifidum and Mercurialis perennis were reported from the Pulinka by Szeląg (2000). Moreover, Botrychium lunaria was observed in the previous two research – in 1968 within the relevè (Table 1) and in 2010 outside the surveyed vegetation plot. On the other hand, the previously unlisted Calamagrostis varia, Silene vulgaris, Linum catharticum and Campanula rapunculoides were recorded in 2022. As for the abundance of relic species, it has remained constant (Sesleria tatrae, Galium anisophyllon) or increased (Scabiosa lucida) over time. The species composition of the basiphilous rocky grassland is presented in Table 1, and divided into archival data (Budzáková et al., 2014; Fabiszewski, 1970) and current data from 2022.
Figure 2
(A–B) The basiphilous, rocky sward with Sesleria tatrae, Calamagrostis varia and Scabiosa lucida on the Pulinka (KŚ, 01.08.2022).

Table 1
Changes in the species composition of basiphilous rocky grasslands with Sesleria tatrae on the Pulinka rock in the Kleśnica Valley in the years 1968–2022.
. Changes in environmental conditions and functional structure of the rocky grassland community
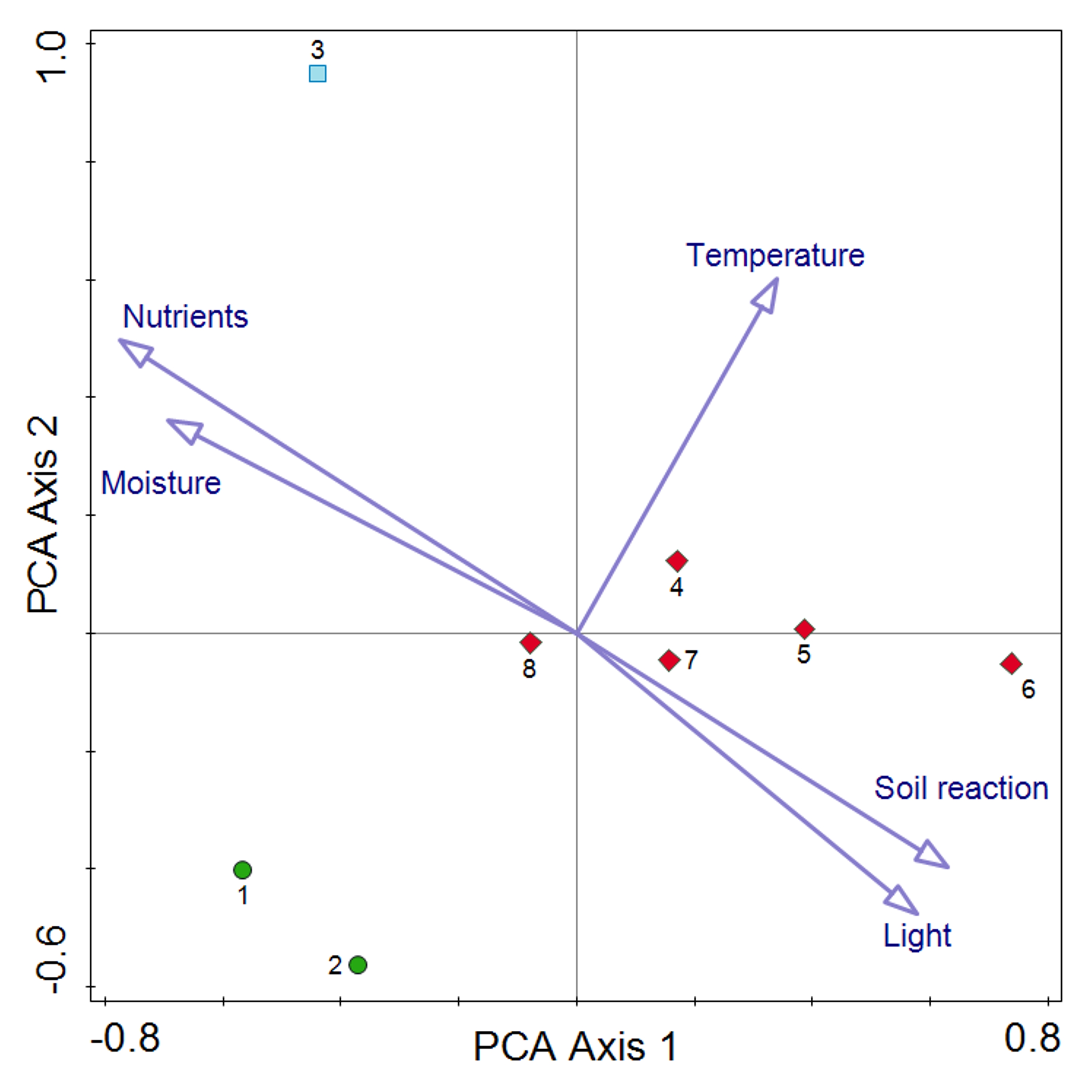
The comparison of the CWMs for temperature, light, nutrients, soil reaction and moisture did not reveal statistically significant differences between phytocoenoses documented in 1968, 2010 and 2022 (Kruskal–Wallis test Hc = 0.02, p = 0.99). In particular, highly comparable are data from 1968 and 2022 (Wilcoxon test W= 9, p = 0.81). The relevè collected in 2010 was distinguished from the others by the CWMs for moisture and nutrients (Figure 3).
Figure 3
PCA plot of distribution of the Sesleria tatrae grassland samples on the Pulinka rock from subsequent periods (1968–2022), with the passive projection of the community-weighted means of EIVEs. There are noticeable differences in the distribution of phytosociological relevés in accordance with the study period, but they are not statistically significant in relation to individual ecological indicators. Explanation: Vegetation plots documented in: 1968 - green circles, 2010 - blue square, 2022 - red diamonds.
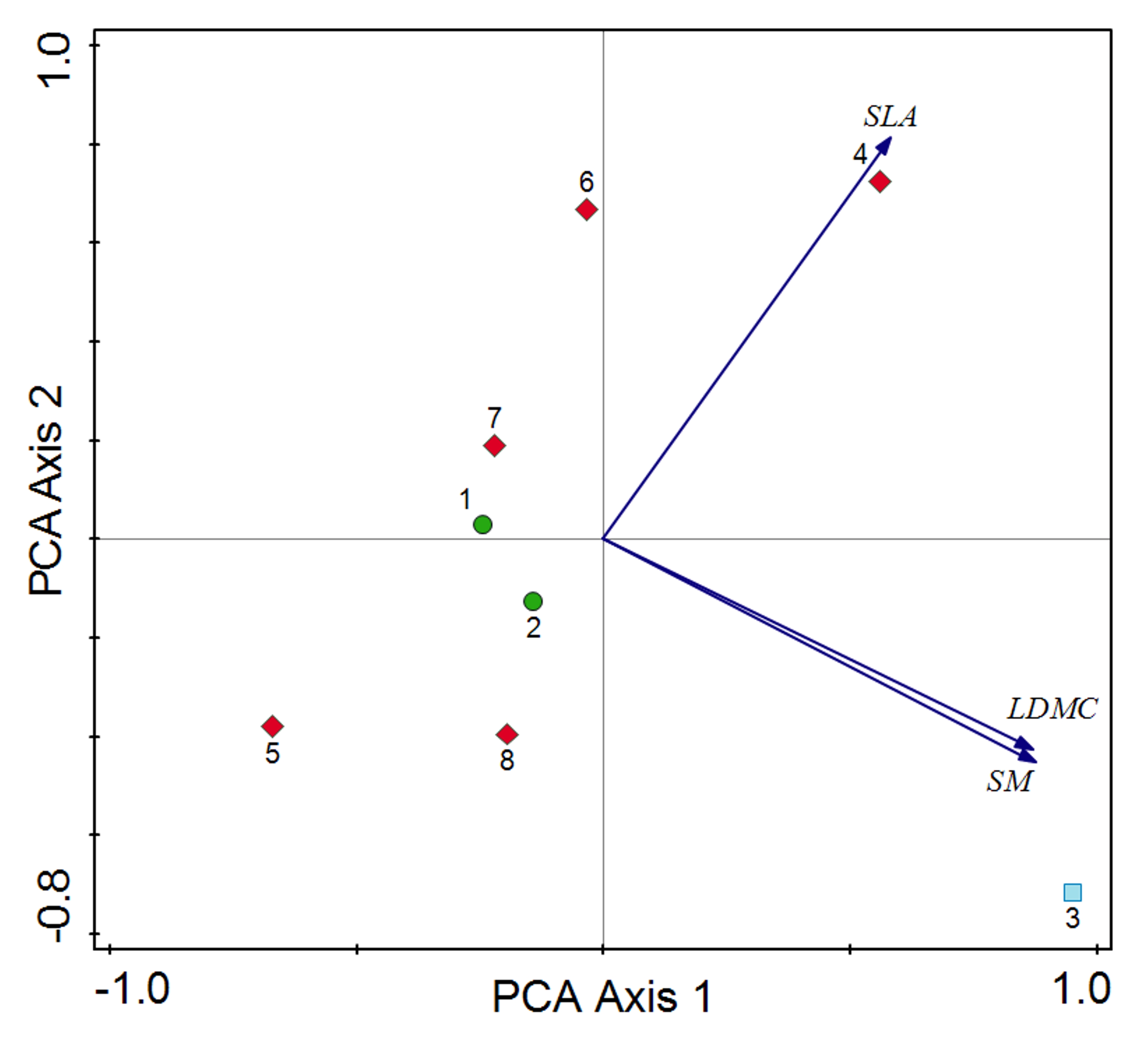
Similarly, the comparison of the CWMs for functional traits did not reveal statistically significant differences between the subsequent study periods (Kruskal–Wallis test results: SLA – Hc = 0.45, p = 0.79; SM – Hc = 3.38, p = 0.18; LDMC – Hc = 2.45, p = 0.29). The relevè collected in 2010 was distinguished from the others by higher values for SM and LDMC (Figure 4), which was associated with the higher abundance of species such as Phleum pratense, Fagus sylvatica, Cotoneaster integerrimus and Veratrum lobelianum, characterized by higher values of these particular traits.
Figure 4
PCA plot of distribution of the Sesleria tatrae grassland samples on the Pulinka rock from subsequent periods (1968–2022), with the passive projection of the community-weighted means for plant functional traits. The differences in the CWMs between 1968 and 2022 are statistically insignificant. Only the relevé from 2010 differs from the others, probably due to the location covering more shaded parts of the grassland with the participation of Veratrum lobelianum, Phleum pratense and high proportion of Fagus sylvatica and Cotoneaster integerrimus juveniles. Explanation: Vegetation plots documented in: 1968 - green circles, 2010 - blue square, 2022 - red diamonds.
The high proportion of nemoral species, which were not recorded in the other study periods, resulted in the highest functional diversity index in the plot collected in 2010 (FD Rao = 1.666). The 1968 and 2022 plots were very similar in this respect (median FD Rao 1968 = 1.162 and 2022 = 1.169), and the differences in functional diversity between them were not statistically significant (Kruskal–Wallis test: Hc = 2.333, p = 0.31).
. Discussion
. The origin of Calamagrostis varia on the Pulinka rock
In Czechia, Calamagrostis varia is known from 12 scattered sites on the lowlands and foothills (Chytrý et al., 2021). The closest one is located 50 km west of the Pulinka site, near České Meziříčí (Hradec Králové Region) at the 250 m a.s.l., where it survives in the vegetation of degraded calcareous fen, formerly harbouring other light-demanding and presumably relic basiphytes (e.g., Tofieldia calyculata, Schoenus nigricans). In Poland, the species has so far only been recorded in the Western Carpathian mountain ranges (Tatra Mts., Pieniny Mts.), and its nearest sites are almost 200 km southeast of the Kleśnica Valley (Zając & Zając, 2001). The genesis of the population of C. varia on the Pulinka could be explained by future genetic studies, but it is likely that the species has been overlooked in field surveys so far, because it co-occurs with the physiognomically similar C. arundinacea. It is therefore possible that C. varia, like other species associated with the Carpathian arc in Poland (Sesleria tatrae, Scabiosa lucida and Galium anisophyllon), is a permanent feature of the flora of the studied area. This could support the hypothesis of an autochthonous origin of the entire colony of dealpine plants, which has been isolated from the compact range that once existed. The co-occurrence of many light-demanding species in relic sites that are strongly isolated from the main range, is a regularly recorded phenomenon both in the Carpathians (Dítě et al., 2018) and in the Czech part of the Sudetes (Kwiatkowski & Krahulec, 2016; Šmarda, 1950). For example, an isolated erlan (metamorphic rock consisting mainly of calcium silicates) rock cliff in the mostly forested Šumárník Reserve (Hrubý Jeseník Mts., 1,070 m a.s.l.) harbours isolated stands of Bupleurum longifolium subsp. vapincense, Conioselinum tataricum, Saxifraga paniculata and Valeriana tripteris. In the Hrubý Jeseník Mts., there are also other local refugia in calcium-rich habitats in Velká Kotlina, Petrovy Kameny and Tabulové Skály, with many relic and disjunctive species including light-demanding basiphytes (Kwiatkowski & Krahulec, 2016). In this respect, Pulinka fits well the concept of old relic, calciphilous, light islands surrounded by generally acidic landscapes of the Sudetes.
. Changes in the size of populations of Scabiosa lucida and Sesleria tatrae over the 50 years
Scabiosa lucida was recorded in 1968 only in one of the two phytosociological relevès (Fabiszewski, 1970). Soon after, it ceased to be observed. As early as 1989, it was reported to occur in the Śnieżnik Massif only in Czechia (Fabiszewski, 1968), and was later declared extinct in the Kleśnica Valley (Brej et al., 1997). In 2000, the species was again reported as rare (Szeląg, 2000), and in the relevè from 2010 (Budzáková et al., 2014) it was not represented at all. Today however, it is one of the dominant species at the site, and its population can be estimated at several hundred tufts. The reason for such significant temporal fluctuations in the abundance of the species is difficult to explain. At the same time, the population size of Sesleria tatrae has remained stable over the study period.
. Changes in species composition of the rocky grassland community
The differences observed in the species composition of the studied plant community are not directional, i.e., they do not involve the withdrawal of a particular group of plants (e.g., nemoral or chasmophytic) and their replacement by another group with specific habitat characteristics. The vascular plant species recorded here, seem to disappear and reappear in a stochastic manner. This may be related to random fluctuations in species composition, but may also be related to the locations of the documented vegetation plots themselves. Admittedly, the site is small, but the selected vegetation plots do not cover the full area of the site, and individual species may have been outside the boundaries of the relevès when they were collected.
All the species that were recorded in one of the subsequent study periods still occur in the Kleśnica Valley, so their appearance and disappearance may be a natural phenomenon, related to the dynamics of individual populations. On the other hand, the stability of the species composition of the rocky grassland community throughout the study period is evidenced by the absence of statistically significant changes observed in the CWMs for ecological vegetation indices and functional traits analyzed, especially between 1968 and 2022. This ecological stability of the Pulinka grasslands contrasts with other studies of high mountain grasslands in the temperate zone (Chen et al., 2014; Dibari et al., 2021; Möhl et al., 2022; Rammig et al., 2010; Staude et al., 2022; Steinbauer et al., 2022), including the Czech part of the Śnieżnik summit (Zeidler et al., 2023), which confirm their exposure to rapid changes in terms of homogenization and poorer species composition, as well as the distribution of plant functional groups due to climate change and eutrophication.
On the other hand, preliminary studies on the natural rocky communities of the lower mountain altitudes indicate that they are still resistant to the global changes taking place, and that any fluctuations in species composition are mainly dependent – as in the case of the Pulinka – on the local factors, and do not show a clearly directional character (Świerkosz & Reczyńska, 2023). It is also possible that the substrate, which is rich in calcium carbonate, has a buffering effect on climate change and increasing atmospheric nitrogen deposition (Nicklas et al., 2021). Noteworthy is the constant but sporadic occurrence of seedlings of some tree species (Picea abies, Salix caprea, Acer pseudoplatanus, Fagus sylvatica), which, however, do not form higher vegetation layers. This may indicate that the stable nature of the studied phytocoenoses may be the result of harsh habitat conditions and microclimate and/or herbivores pressure.
. Syntaxonomic position of the studied grasslands
To determine the syntaxonomic position of the calcareous grasslands from Pulinka, we analyzed their species composition in the context of current classifications in Poland (Matuszkiewicz, 2007), Slovakia (Jarolímek & Šibík, 2008; Kliment et al., 2005; Kliment & Valachovič, 2007), Czechia (Chytrý et al., 2021) and against the background of the latest classifications of plant communities in Europe (Mucina et al., 2016; Preislerová et al., 2022). It has been so far suggested, that the studied phytocoenoses belong to the Seslerion tatrae alliance (Budzáková et al., 2014; Fabiszewski, 1970) which is defined as “Alpine and subalpine chionophilous blue-grass swards on leeward slopes with deeper soils on calcareous bedrock" in Slovakia (Jarolímek & Šibík, 2008) and similarly in Europe (Mucina et al., 2016). In general, European classification is somewhat similar to the Slovak approach where the Western Carpathian alpine vegetation is best developed and represent most of the highest syntaxonomic units. According to European scope, three alliances of calciphilous grasslands: Seslerion tatrae, Astero alpini-Seslerion calcariae Hadač ex Hadač et al. 1969 nom. invers. propos., and Caricion firmae Gams 1936 are distinguished. In this respect, the Polish classification system is considerably simpler, since it recognizes only one alliance within the Seslerietea variae Br.-Bl. 1948 em. Oberdorfer 1978 class (separated from Elyno-Seslerietea Br.-Bl. 1948), namely Seslerion tatrae (Matuszkiewicz, 2007). However, if we intend to classify Pulinka grasslands, we should follow the classifications based on a larger part of the Western Carpathian or European system; they appear to be much more complex thus better recognize various types of vegetation.
Geographically, in the Sudetes there is only one base-rich type of grassland included in the Carici rupestris-Kobresietea bellardii Ohba 1974 class (Mucina et al., 2016): Agrostion alpinae alliance, represented by two associations of very limited range. The first one is Saxifrago oppositifoliae-Festucetum versicoloris Wagnerová et Šírová 1971 known from the Karkonosze Mts., from two glacial cirques on the Czech side (Velká Kotelní jáma, Obří důl) and one cirque (Mały Śnieżny Kocioł) on the Polish side of the mountain range (Kočí, 2007a). The second one, Saxifrago paniculatae-Agrostietum alpinae Jeník et al. 1980, is endemic to the steep slopes of the Velká Kotlina cirque in the Hrubý Jeseník Mts. (Kočí, 2007b). This alliance represents mainly impoverished types of the subalpine grasslands and it has no really good diagnostic species. It should also be noted that the discussed phytocoenoses from the Pulinka share only a single species with this alliance which is Scabiosa lucida – a species diagnostic to the whole class, what strongly decreases its distinctive value.
There is also a possibility to classify Pulinka grasslands as dealpine communities outside the Elyno-Seslerietea or Carici-Kobresietea classes. Pulinka is located deep in the beech forest zone and Śnieżnik Mt. is devoid of basiphilous alpine or subalpine vegetation, typical of this class. Analogous communities in Slovakia and Europe are assigned to the alliance Diantho lumnitzeri-Seslerion (Soó 1971) Chytrý et Mucina in Mucina et Kolbek 1993 of the Festuco-Brometea Br.-Bl. et Tx. ex Soó 1947 class (Mucina et al., 2016), but this alliance combines thermophilous and dealpine elements. In Pulinka, thermophilous species are mostly missing (which is typical of the Sudetes in general compared to the Carpathians). Therefore, it seems that Pulinka grasslands represent a very special and unique combination of biogeographic and historical circumstances. For the following reasons it is very difficult to classify this relic grassland community:
Compared to the Carpathians and Alps, the Sudetes are almost lack of basic substrates and basiphilous sub/alpine vegetation, except for the alliance Agrostion alpinae which is significantly species-impoverished unlike stands in the high mountain ranges of Central Europe.
The Sudetes were much more influenced by glaciers (compared to the Alps/Carpathians). Therefore, glacial refugia were rarer here due to less diverse landscape, relatively uniform geomorphology and prevailing acidic substrates. Such specific conditions also decreased the number of species typical of the basiphilous, relic rock vegetation.
For this reason, we propose to classify Pulinka grasslands as a community representing the class Elyno-Seslerietea and the order Seslerietalia caeruleae, but without assigning it into any so far described alliances. This approach is supported by a constant and high proportion of Sesleria tatrae and Scabiosa lucida, and the lack of other diagnostic species of subalpine grasslands. However, it will be the task for future research at larger spatial extents to evaluate more precisely the syntaxonomic position of the studied grasslands.
. Conclusions
The rocky grassland community developing on the top of the Pulinka rock in the Kleśnica Valley (Śnieżnik Massif) represents a unique, local community, of the Elyno-Seslerietea class, and the order Seslerietalia caeruleae. The latter is the phytosociological identifier of the habitat “6170 Alpine and subalpine calcareous grasslands” in Poland. As such it should be included in national, scientific studies on the diversity of plant communities, and protected within the Natura 2000 network.
Between 1968 and 2022, fluctuations in species composition occurred within the studied community, but these fluctuations did not significantly affect its functional structure and ecological indicators. The observed fluctuations do not apply to the geographically important species, which here have the only sites in the Polish Sudetes (Sesleria tatrae, Scabiosa lucida, Euphrasia micrantha, Galium anisophyllon). Moreover, in 2022 Calamagrostis varia, which is known in Poland only from the Carpathians, was found in the studied community.
Our results are in contrast to the research conducted in other types of plant communities which revealed significant changes in species composition such as disappearance of rare species of vascular plants with narrow ranges due to thermophilization and eutrophication phenomena (Staude et al., 2020, 2022).
