. Introduction
Rehmia Kremp. is a recently resurrected genus that currently comprises 14 species that were formerly placed in the genus Rhizocarpon DC. (Möller et al., 2025). Along with Catolechia Flot., Haugania E. J. Möller & Timdal, Poeltinula Hafellner, and Rhizocarpon DC., it is placed in the family Rhizocarpaceae M. Choisy ex Hafellner (Fryday et al., 2024; Miadlikowska et al., 2006, 2014; Möller et al., 2025). Together with the family Sporastatiaceae Bendiksby & Timdal, it forms the order Rhizocarpales Miądl. & Lutzoni ex Miądl. & Lutzoni (Miadlikowska et al., 2006, 2014). Members of this genus are characterized by a crustose thallus, lecideine exciple, Rhizocarpon-type asci (without an ocular chamber) with a well-developed tholus, non-amyloid medulla, and halonate and hyaline, mostly muriform ascospores (Möller et al., 2025).
The most commonly used infrageneric classification of Rhizocarpon s.l., which encompasses a broad assemblage of species prior to the recent taxonomic revisions by Möller et al. (2025), was based on the presence of rhizocarpic acid. Thomson (1967) divided the genus into two subgenera: yellow Rhizocarpon (characterized by the presence of rhizocarpic acid) and brown Phaeothallus Thomson (lacking rhizocarpic acid). Previously, all species were divided into two sections based on the number of septa; species with one-septate ascospores were placed in section Catocarpon, while those with four-septate to muriform ascospores were placed in section Eurhizocarpon (Fries, 1874). However, Ihlen & Ekman (2002) noted that infrageneric classifications based solely on the presence or absence of rhizocarpic acid or the number of septa are unnatural. Furthermore, species characterized by the presence of stictic acid, amyloid medulla, or hyaline/pigmented ascospores were also not found to form monophyletic groups (Ihlen & Ekman 2002). These results were recently corroborated, and taxonomic changes were made by Möller et al. (2025), including the resurrection of the genus Rehmia.
Rehmia furfurosa (H. Magn. & Poelt) E. J. Möller, Timdal, Haugan, Bendiksby (= Rhizocarpon furfurosum H. Magn. & Poelt) is a non-yellow species that is easily distinguishable from other members of the genus and family, as it is the only species with blastidia (Schade, 1933; Poelt, 1955, 1969, 1980). It typically occurs in a sterile form (Smith et al., 2009). The protologue describes the asci as containing eight submuriform, ellipsoid to narrowly ellipsoid ascospores (Poelt, 1955). The thallus contains stictic acid, although some authors have suggested that norstictic acid may also be present (Wirth, 1995; Wirth et al., 2013). However, Ihlen (2004) indicated that specimens with the norstictic acid chemotype should be referred to as Rhizocarpon aff. furfurosum.
Rehmia furfurosa is a rare lichen found only in Europe (Smith et al., 2009), with records from Great Britain (Purvis & James, 1985), Germany (Schade, 1933, 1935), Norway (Timdal & Holtan-Hartwig, 1988), Czechia (Malìček, 2010; Steinová et al., 2013), Slovakia (Guttová et al., 2013), France (Roux et al., 2017), Russia (Urbanavichus & Urbanavichene, 2004), Svalbard (Konoreva & Chesnokov, 2021), Italy (Nimis & Stefano, 2003), and Austria (locus classicus) (Poelt, 1955). However, according to Purvis & James (1985), this species is overlooked. It is one of the species typically found in metal-rich areas and grows predominantly on siliceous rock and old walls rich in iron minerals, especially in old mines, where it can be abundant (Purvis & James, 1985; Schade, 1935; Smith et al., 2009). Schade (1935) emphasized that this taxon (as Rhizocarpon obscuratum f. granulosum) only occurs on sunny, old mining heaps in the Ore Mountains (in German: ‘Erzgebirge’) and the ‘Muldenland’, especially in Freiberg, Germany. Nevertheless, Purvis & James (1985) pointed out that the taxon was also found in sheltered situations, but then it differed in color of thallus which was cream-pale fawn rather than violaceous deep brown (when on exposed sites).
In 2022 and 2023, material morphologically similar to Rehmia furfurosa was found in the Western Sudetes, historically one of the most important centers of copper mining and smelting in Poland (Dziekoński, 1972). However, the collected specimens differed in ascospore size and number of cells per ascospore from those reported in the protologue. Therefore, it became necessary to generate nuITS rDNA sequences for this taxon, including specimens with submuriform ascospores (as described in the protologue) and those with muriform ascospores (as in the Polish specimens), to resolve whether the discrepancies are due to intraspecific variation or whether they represent two distinct species with similar thallus morphology and habitat preferences. Furthermore, given the rarity of this taxon in Europe and the challenges of its identification, particularly because of its frequent occurrence in the sterile form, the potential ecological niche was determined.
In parallel with the studies in Poland, two of us (EJM and ET) built a DNA barcode library for Rhizocarpon s.l. species, based mainly on Norwegian material. We merged our Polish and Norwegian sequence data to increase the robustness of the species hypothesis.
. Materials and methods
. Taxon sampling
Specimens of Rehmia furfurosa from Poland were collected in 2022 and 2023 in the Western Sudetes at the Colorful lakelets (in Polish: ‘Kolorowe Jeziorka’), a group of artificial ponds formed at the sites of former mines in the Landeshut Ridge (in Polish: ‘Rudawy Janowickie’) mountain range. All collected specimens were deposited in the herbarium of the University of Gdańsk (UGDA L) and one in the private collection of K. Szczepańska. Additional specimens were studied: twelve from Norway (herbarium O); five from the Czechia (herbarium Malíček); one from France (herbarium Poumarat); one from Austria (holotype, herbarium M) and three from Germany (including exsiccate published by Alvin Schade; herbaria M and GLM). Detailed data of specimens (including herbarium number, location, substrate, and date of collection) are presented in Chapter 6 – Taxonomy, at the end of the paper.
Morphological and anatomical identifications were performed using a Nikon SMZ-10 and a Nikon Eclipse E200 stereo and compound microscopes. The following features were measured (all in water): apothecial diameter (mm), hymenium height, length and breadth of ascospores, and the number of cells per ascospores. Images of the selected specimens were captured using a Zeiss SteREO Discovery V12 microscope and processed using ZEN 3.2 (blue edition) software, with additional image modifications performed in GIMP 2.10.38.
Secondary metabolites were analyzed by thin-layer chromatography (TLC) using solvent system C. Insoluble lichen pigments (if apothecia were present) were tested with 10% potassium hydroxide (KOH), 10% hydrochloric acid (HCl), and 50% nitric acid (HNO₃), and amyloidity was tested with Lugol’s solution without KOH pretreatment. All chemical analyses were performed as described by Orange et al. (2001).
. DNA extraction, PCR amplification, and DNA sequencing
Freshly collected specimens from the Colorful Lakelets were used for DNA extraction. The CTAB protocol was employed to isolate genomic DNA from the thallus (Guzow-Krzemińska & Węgrzyn, 2000), with minor modifications: the incubation at –20°C was extended to 24 hours instead of 30 minutes, centrifugation was carried out at 4°C, and Vivid Violet (EURX®) was used instead of glycogen.
The fungal nuclear internal transcribed spacer (ITS: ITS1 – 5.8S – ITS2) region of the rDNA was amplified using two primers: ITS1F (Gardes & Bruns, 1993) and ITS4 (White et al., 1990). The amplification process involved the following steps: an initial denaturation at 94°C for 3 minutes, followed by 33 cycles of denaturation at 94°C for 30 seconds, annealing at 52°C for 45 seconds, and extension at 72°C for 1 minute, with a final extension at 72°C for 10 minutes. The PCR products were purified using a Clean-Up Concentrator Kit (A&A Biotechnology, Poland) following the manufacturer’s instructions. Sequencing was performed by Macrogen® in the Netherlands using amplification primers. The newly obtained sequences were deposited in GenBank (numbers PX402211–PX402214 & PX363698–PX363703).
The Norwegian specimens were sequenced by the DNA barcoding project OLICH (Marthinsen et al., 2019) at the Norwegian Barcode of Life (https://www.norbol.org), that is, DNA extraction, PCR, and sequencing were performed at the Canadian Center for DNA Barcoding (https://ccdb.ca) using the primer pairs ITS1F/ITS4 or ITS5/ITS4 (Gardes & Bruns, 1993; White et al., 1990).
. Sequence alignment and phylogenetic analysis
The newly generated sequences were identified as belonging to the Rhizocarpaceae family based on BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) results. All available Rhizocarpon s.l. ITS sequences (excluding those labeled as Rhizocarpon geographicum (L.) DC.) were downloaded and automatically aligned using MAFFT (https://mafft.cbrc.jp/alignment/server/index.html). Sequences of R. geographicum were excluded because of the large number of available entries and uncertainty regarding their taxonomic accuracy (Jaskólska, unpublished). Sequences lacking either the ITS1 or ITS2 regions were removed from the alignment. To avoid bias, all terminal gaps were replaced with the letter “N” to represent any nucleotide. A phylogenetic tree was constructed using Bayesian Inference (BI) implemented in MrBayes v3.2.7 (Ronquist et al., 2012), run via the CIPRES Science Gateway (Miller et al., 2010). The best-fit evolutionary model, GTR+I+G, was selected based on the Akaike Information Criterion (AIC) using PAUP v4.0a169 (Nylander, 2004). Two parallel MCMCMC runs were conducted, each with four chains for 6 million generations, and sampling every 1,000 generations. The initial 25% (1500 trees) from each run were discarded as burn-in. The resulting tree, along with a summary of posterior parameter estimates, including convergence diagnostic (Estimated Sample Size – ESS; and Potential Scale Reduction Factor – PSRF) and Heidelberger diagnostic tests, is provided as supplementary material (S1). All diagnostic tests were performed in R using the ‘coda’ package (Plummer et al., 2006).
A second phylogenetic tree was constructed using all sequences that formed a clade near Rehmia furfurosa, along with two outgroup sequences of Haugania oederi (Ach.) E. J. Möller & Timdal (≡ Rhizocarpon oederi (Ach.) Körb.). H. oederi was chosen as the outgroup because it was formerly classified within Rhizocarpon, which is traditionally understood to include a broad range of species and represents a closely related but distinct genus (Möller et al., 2025). Sequences were automatically aligned using Multiple Sequence Alignment (MSA) via the MAFFT algorithm on the GUIDANCE web server. A total of 100 alternative guide trees were used. The GUIDANCE2 algorithm was applied to account for alignment uncertainties (Landan & Graur, 2008; Penn et al., 2010; Sela et al., 2015). The alignment score was approximately 0.96, and positions with reliability scores below 0.93 were removed. The GTR+I+G model was selected (AIC, PAUP 4.0a169), and Bayesian inference was run in MrBayes v3.2.7 via CIPRES, with two parallel MCMCMC runs, four chains, six million generations, sampling every 1,000 trees, and 25% burn-in (Miller et al., 2010; Nylander, 2004; Ronquist et al., 2012). All summaries of posterior parameter estimates, including convergence diagnostics (ESS, PSRF) and Heidelberger diagnostic tests, are provided as supplementary material (S1).
The consensus tree was visualized using R (v. 4.3.3) and RStudio (v. 2024.04.1 “Chocolate Cosmos”) using the ‘ape’ (v. 5.8) and ‘graphics’ (v. 4.3.3) libraries, and further edited in Inkscape (Inkscape Project, 2023; Paradis & Schliep, 2019; Posit Team, 2015; R Core Team, 2024). Support values for Bayesian Inference Posterior Probability (PP) are provided next to the branches of the phylogenetic tree.
. Species distribution analysis
. Dataset – occurrence and environmental data
To prepare a species distribution map for Rehmia furfurosa, occurrence data from the Global Biodiversity Information Facility (GBIF.org, 2024; accessed 26.04.2024) were used. The dataset consisted of GPS data from preserved specimens and human observations. Human observation records were primarily contributed by the British Lichen Society and the West Wales Biodiversity Information Centre, and almost all records originated from the United Kingdom of Great Britain and Northern Ireland. In addition, GPS data from Czech and Polish specimens were included. As R. furfurosa has only been reported in Europe, all explanatory variables were restricted to this region.
Data on the concentrations of ten metals in European topsoil were tested: iron (Fe), lead (Pb), cobalt (Co), chromium (Cr), cadmium (Cd), silicon (Si), zinc (Zn), copper (Cu), arsenic (As), and aluminum (Al) (Ballabio et al., 2018, 2024; Reimann et al., 2014; Tóth et al., 2016; Van Eynde et al., 2023). The predictors of species occurrence were based on GEMAS data for Fe, Al, and Si and LUCAS data for the remaining explanatory variables. For analysis purposes, raster data for iron, aluminum, and silicon were selected based on sampling from pastureland soils using X-ray fluorescence analysis. GEMAS concentration data are displayed at 72 levels on an interval scale, with Fe concentrations ranging from <2000 mg/kg to >80,000 mg/kg, Si from <50,000 mg/kg to >450,000 mg/kg, and Al from <10,000 mg/kg to >125,000 mg/kg.
As LUCAS data are only available for the European Union (excluding Croatia and including Great Britain) and GEMAS has some missing data, the final model does not introduce all potential arrangements.
. GIS and Maximal Entropy
Preparing data
All data transformations were conducted using R (v. 4.3.3) and RStudio (v. 2024.04.1 “Chocolate Cosmos”) using the following libraries: ‘terra’ (v. 1.8–10), ‘sf’ (v. 1.0.16), ‘raster’ (v. 3.6.26), and ‘spThin’ (v. 0.2.2) (Aiello‐Lammens et al., 2015; Hijmans, 2023, 2025; Pebesma, 2018; Pebesma & Bivand, 2023). The cell sizes of the predictor raster data were resampled to a uniform resolution of 1000 m, with consistent extents and a single coordinate reference system: the World Geodetic System 1984 (WGS 84; EPSG:4326). Resampling of Fe, Al, and Si used the nearest-neighbor method for estimating cell values, whereas bilinear interpolation was used for the other predictors.
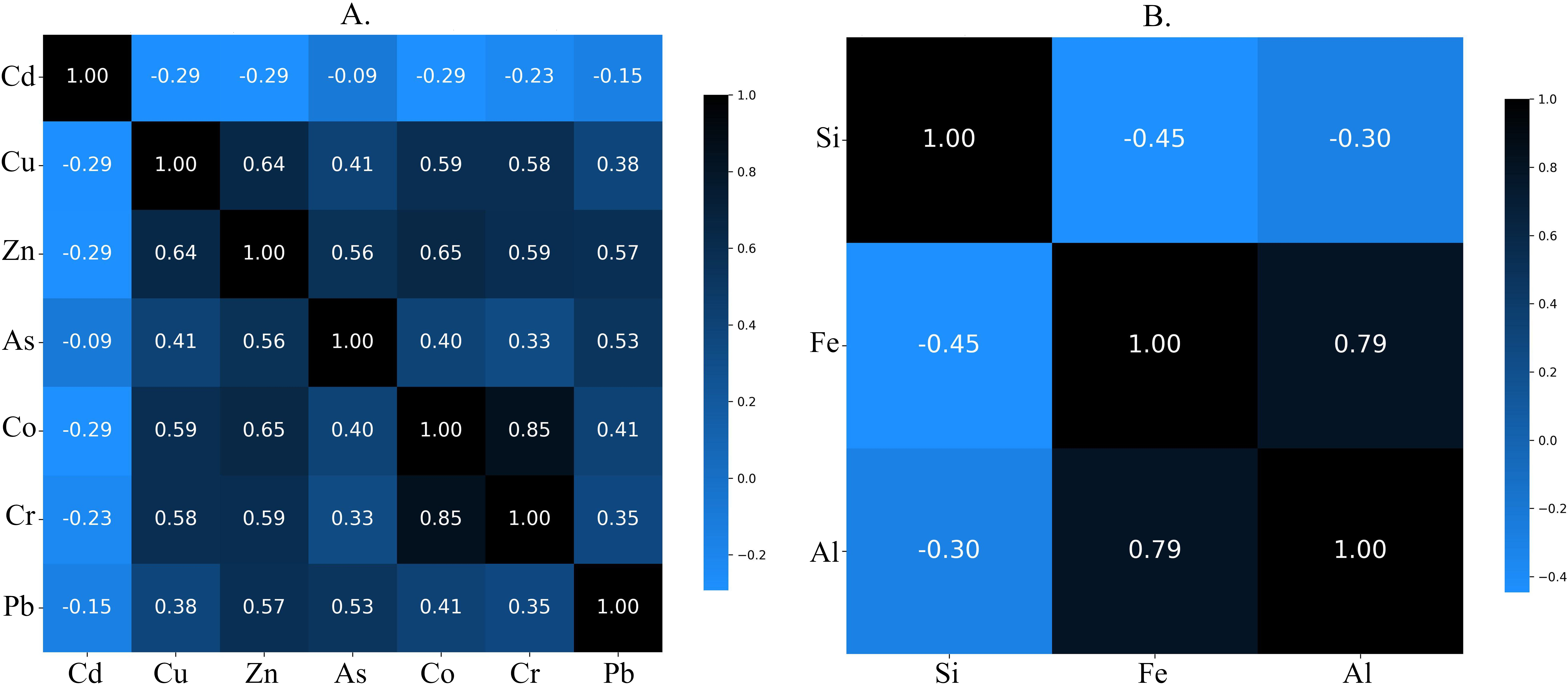
Because MaxEnt may be sensitive to correlated data, heavy metal concentrations were extracted for 10,000 random points in R using the ‘dismo’ (v. 1.3.14) and ‘raster’ (v. 3.6.26) libraries (Hijmans, 2023; Hijmans et al., 2023). A correlation matrix (Figure 1) was generated using Python 3.11.7 (Python Software Foundation, 2024) within the Anaconda environment 24.1.2 (Anaconda Software Distribution, 2024), with the ‘pandas’ (v. 2.1.4) and ‘matplotlib’ (v. 3.8.4) libraries (Hunter, 2007; McKinney, 2010). The Spearman method was applied for Si, Fe, and Al (Figure 1B), while the Pearson method was used for the remaining predictors (Figure 1A). Only Co and Cr showed a strong positive correlation (r above 0.8; r2 > 0.64), leading to the exclusion of Cr from further analysis.
Figure 1
Correlation matrix by A. Pearson and B. Spearman methods for extracted 10,000 random points.
All sites located outside the raster data range were removed. Additionally, because nearly half of the data points were located in Great Britain, spatial thinning with 100 iterations and a 5-kilometer threshold was applied to avoid sampling bias. Consequently, the dataset was reduced to 63 presence points for R. furfurosa from an initial 219. The thinned observations were then split into two groups: 47 for training and 16 for testing purposes.
Modelling
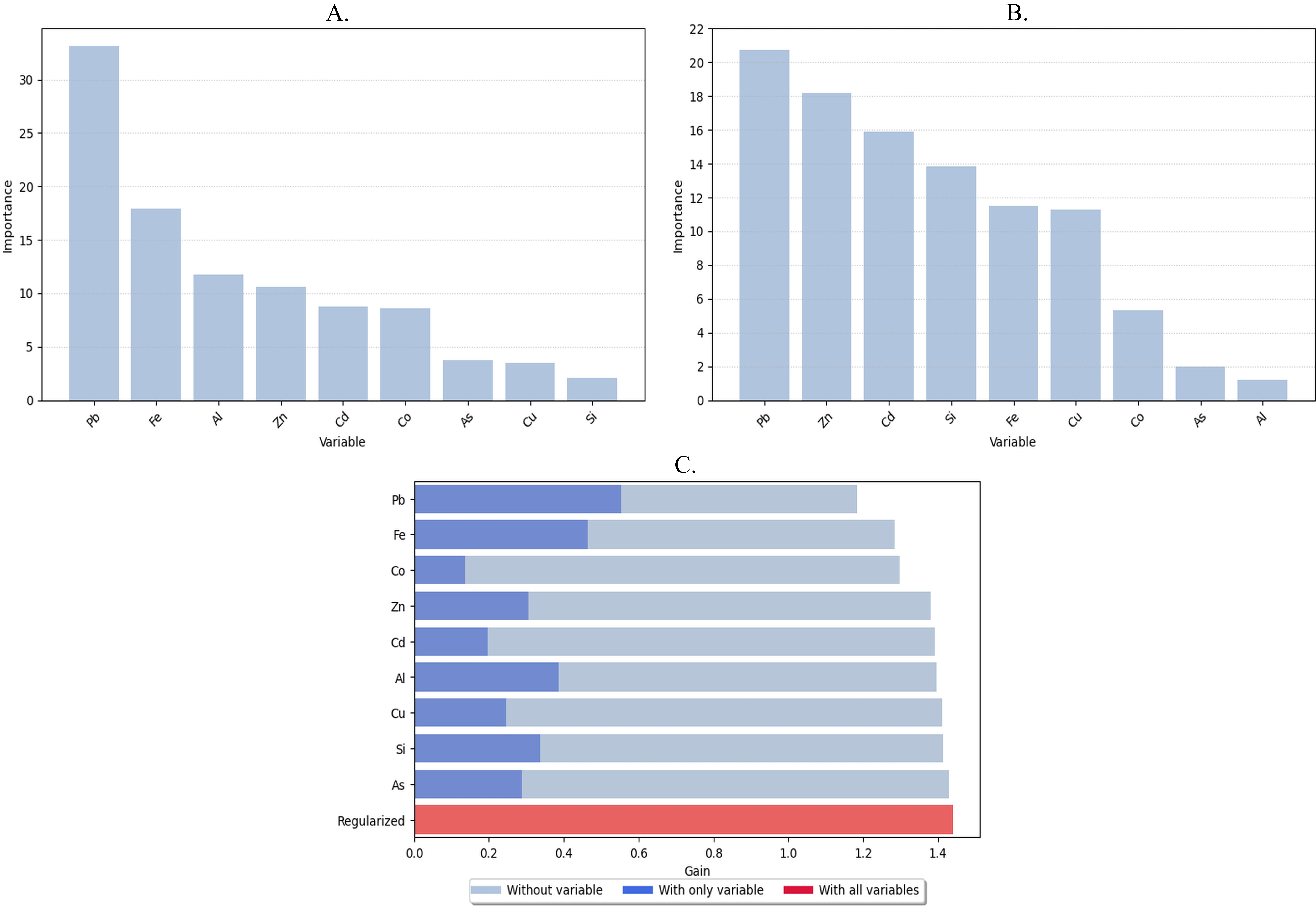
The species distribution map was modelled in R using the ‘dismo’ (v. 1.3.14) and ‘rJava’ (v. 1.0.11) libraries (Hijmans et al., 2023; Urbanek, 2024). To improve the model fitting, the explanatory variables were transformed using basis expansion functions (linear, quadratic, and hinge). The number of iterations was set to 500, with the complementary log-log (C-log-log) function selected as the link option. To select the most significant explanatory variables, a percent contribution analysis, permutation feature importance, and Jackknife test were performed. The visualizations for these plots were prepared in Python using the ‘matplotlib’ (v. 3.8.4) library (Hunter, 2007).
The top six metals with the highest importance (based on permutation importance, percent contribution, and Jackknife test results) were selected for the final model. Specifically, variables that had the highest gain when used alone and those with the lowest regularized gain when excluded were compared to each other. The final set of selected variables included Fe, Pb, Zn, Al, Cd, Si, and Co. Response curves for these metals were created, showing the relationship between each variable and species distribution when the other predictors were held at zero. The figure are available in the Supplementary Data (Supplementary S1).
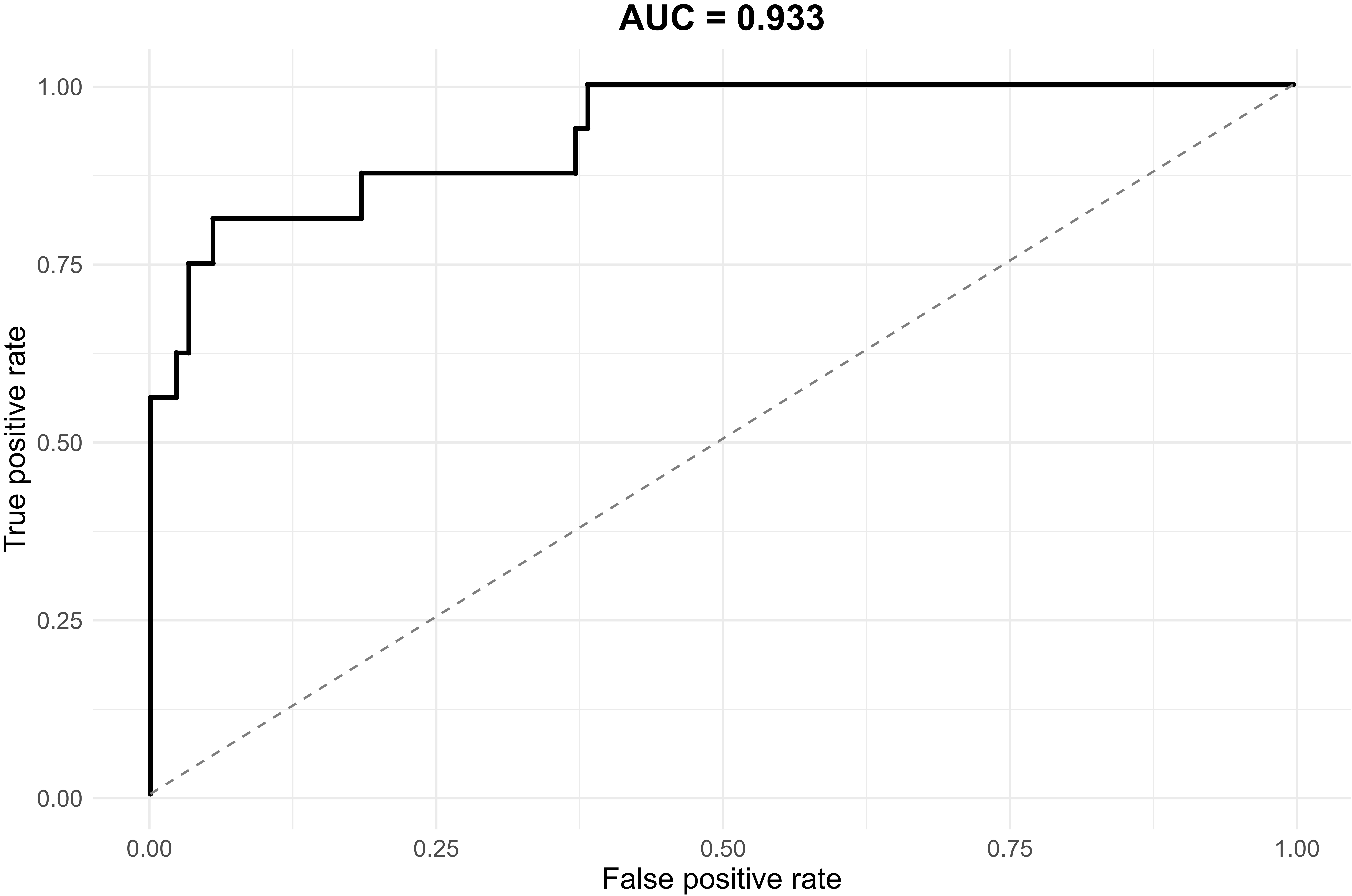
The final model was evaluated using 92 pseudo-absence data points and 16 presence data points (from the training set), with receiver operating characteristic (ROC) and area under the curve (AUC) methods employed for validation. Map visualizations were prepared using ArcGIS Pro 3.2.2 software (ESRI, Redlands, California, USA) with the Lambert Azimuthal Equal Area coordinate reference system for Europe (EPSG:3035).
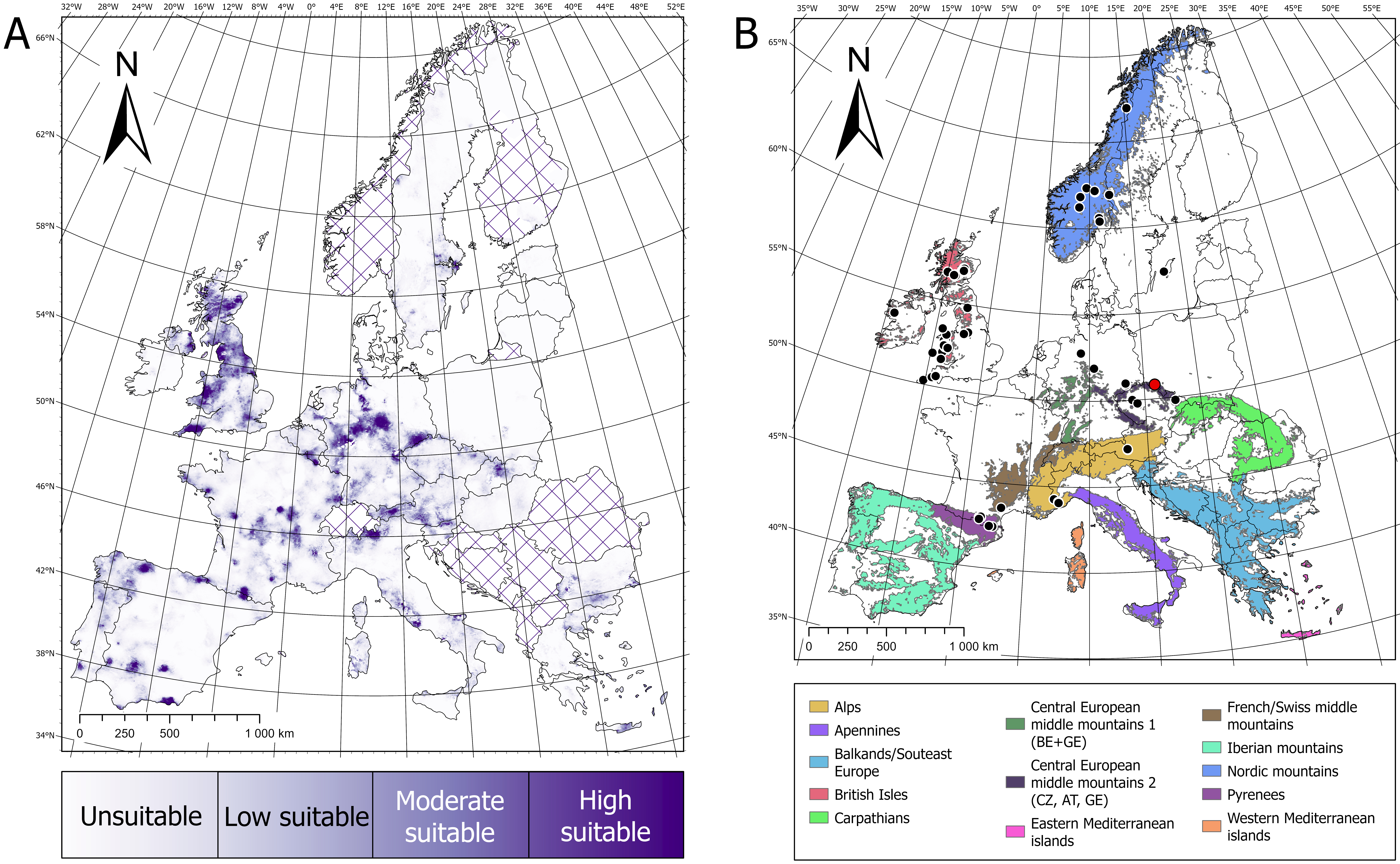
Additionally, a map of the known localities of R. furfurosa was created (based on GBIF data and newly obtained localities in this study). This map also includes a shapefile of mountain massifs or groups of massifs, as defined by the European Environment Agency (2010).
. Results
. Molecular data and morphological variation of Rehmia furfurosa
In this study, we generated 10 nuITS rDNA sequences from material referred to as Rehmia furfurosa from Polish and Norwegian specimens (Figure 2). The newly obtained sequences formed a strongly supported group (PP = 100) forming a clade together with R. reducta (Th. Fr.) E. J. Möller, Timdal, Haugan, Bendiksby, and R. umbilicata (Ramond) E. J. Möller, Timdal, Haugan, Bendiksby. Specimens referred to as R. furfurosa with a small number of cells per ascospore (as described in the protologue) were intermingled with specimens with a larger number of cells and muriform ascospores, thus confirming that they represent a single species.
Figure 2
Phylogenetic tree generated using Bayesian analysis based on the ITS dataset, with posterior probability values near the branches. Newly obtained sequences of Rehmia furfurosa from Poland and Norway are marked by blue rectangular polygons. Other sequences outside the polygon were retrieved from GenBank. The voucher contains the organism name and accession number. Specimens with muriform ascospores are represented by vouchers 1354 and UGDA L-64442 (from larger apothecia), while those with submuriform ascospores are represented by UGDA L-64442 (from smaller apothecia), O-L-163730, O-L-166436, O-L-169766, and O-L-179949. Whereas UGDA L-64421, UGDA L-64446, O-L-239088, O-L-243115 & O-L-239319 represent sterile specimens.

All studied specimens from Czechia were sterile, in contrast to those from Austria, France, Germany, Norway, and Poland. The apothecia measured 0.2–0.9 mm in diameter, with regular, flat to convex, and sometimes umbonate discs (Figure 3B, F–H). The disc was reddish-brown (especially when wet) or black. Ascospores from the holotype were not examined because of the small size of the specimen and the availability of data in the protologue (Figure 3A, B). Specimens from Germany had ascospores with up to 8 cells, and those from Norway had up to 9 cells and measured 10–22 × 10–14 µm. Ascospores in the French specimen (Figure 3H) were damaged, but exhibited a submuriform structure with up to seven cells. In Polish specimens, most had relatively large (22–29 × 7–14 µm), muriform ascospores with up to 18 cells, whereas one (Szymczyk s.n., UGDA L-65531) had only submuriform ascospores measuring 15–19 × 6–9 µm in size. In some specimens (for example, UGDA L-64442), larger and muriform ascospores were found in larger apothecia, which co-occurred with smaller apothecia on the same thallus and contained smaller and submuriform (sometimes only transversely septate) ascospores. Additionally, some mature apothecia contained both submuriform and muriform ascospores. In both types of apothecia, the ascospores appeared to be mature, despite the differences in size. The hymenium in all the studied specimens was up to 150 µm high.
Figure 3
Morphology of Rehmia furfurosa. A. Thallus of the holotype (M-0024899) (scale bar: 0.02 mm); B. Thallus and apothecium on a damaged part of the holotype (scale bar: 0.05 mm); C. & D. rarely encountered prothallus (O-L 163730) (scale bars: 0.1 & 0.02 mm); E. granular thallus (O-L 163730) (scale bar: 0.01 mm); F. – H. apothecia with umbonate discs; respectively GLM-L-0021351 (lectotype); O-L182027; Poumarat SP 2016-85 (scale bar: 0.02 mm).

In all tested specimens of R. furfurosa, we detected stictic acid (as the major compound) together with constictic acid (reported for the first time, but not found in all specimens) and cryptostictic acid, both in minor or trace concentrations. Norstictic acid was not detected in any of the analyzed specimens.
. GIS analysis
The presence of Fe, Pb, Zn, Al, Cd, Si, and Co was found to be a significant factor for species distribution, according to all permutated importance and jackknife tests (Figure 4). All results indicate that Pb and Fe are highly important predictors. Si and Al were less important; however, after comparing the AUC with and without these variables, these predictors were added to the model.
Figure 4
Results of the selection of significant explanatory variables. A. Percent contribution; B. Permutated importance; C. Jackknife test.
The model based on the proposed predictors was tested using 92 pseudo-absence points and 16 presence points with the receiver operating characteristic (ROC) area under the curve (AUC) (Figure 5). An AUC value of 0.933 indicates that the final model exhibited excellent discriminatory ability (Muller et al., 2005).
The species distribution model, based solely on the presence of heavy metals in the topsoil, highlighted the potential habitat suitability for Rehmia furfurosa across Europe (Figure 6A). Owing to the exclusion of certain countries from the metal sampling, which served as explanatory data, the final model may not capture the full potential range of the species. The most suitable habitats are mainly concentrated in mountainous regions within several major massifs, where R. furfurosa has been found according to literature data (Figure 6B):
The Western and Central Alps (Germany, Italy, France, and Austria), where the holotype was collected;
The Carpathian Mountains, particularly in Slovakia;
The Central European middle mountains 2 (CZ, AT, DE) include the Sudetes along the Czech-Polish border (notably near the Colourful Lakelets) and areas near Saxony, where Schade (1933, 1935) reported the abundant occurrence of R. furfurosa (as Rhizocarpon obscuratum f. granulosum);
The Central European middle mountains 1 (BE, DE) – within Central Germany;
The French-Swiss middle mountains, mainly in France;
The British Isles encompass Cornwall ceremonial county, the Cambrian Mountains, the Pennines in England, the Scottish Highlands, the Grampian Mountains, and the eastern parts of Northern Ireland.
Furthermore, the model identified several areas of high suitability where the species has not yet been documented:
The Balkans: western Bulgaria, the eastern regions bordering Turkey, and northern Greece;
The Eastern Mediterranean islands, including Crete and the surrounding Greek islands;
The Iberian Mountains – mainly in northern (Cantabrian Mountains) and southern Spain (Cordilleras Béticas, Sierra Nevada), as well as central Spain (Sierra Morena, Montes de Toledo), and in Portugal’s Serra da Estrela;
The Pyrenees, which stretch across northern Spain and southern France;
The Carpathian Mountains, mainly in northern Hungary and slightly in southern Poland (where R. furfurosa has been found, but only in the Sudetes);
The Apennines – central Italy (Appennino Centrale) and northern Sicily.
Figure 6
(A) Model of the distribution of Rehmia furfurosa in Europe. Below the map is a gradient of color describing the degree of suitability of the area. Areas with missing data are marked with a grid shape. (B) Map with confirmed distribution of R. furfurosa according to GBIF. Localities with new reports of R. furfurosa in Poland are marked by red circles. The names of the mountain massif with their color symbols are below the map.
. Discussion
Rehmia furfurosa most frequently occurs in a sterile form, making its identification more challenging, as determining the type of asci is crucial for distinguishing the family Rhizocarpaceae. This is particularly problematic, as such specimens may be mistaken for other morphologically similar lichen species, such as Lambiella furvella (Mudd) M. Westb. & Resl or Lecidella meiococca (Nyl.) Leuckert & Hertel (Fryday et al., 2024). However, these species differ in their chemical composition, as L. furvella contains gyrophoric and rarely norstictic acids, whereas L. meiococca produces atranorin and chlorinated xanthones (i.e., thiophanic, isoarthothelin, and other) (Smith et al., 2009). In this study, specimens of R. furfurosa containing norstictic acid rather than stictic acid were not tested, leaving the question open as to whether this is a result of chemical variability or indicative of a distinct species. Rehmia furfurosa is also one of the few species within the family that has evolved vegetative propagules (Fryday et al., 2024; Ihlen, 2004; Smith et al., 2009). Other species within the family that exhibit this character include Rhizocarpon ridescens (Nyl.) Zahlbr. and R. sorediosum Runemark, both of which produce soredia and contain rhizocarpic acid (Runemark, 1956). The former also contains psoromic acid (Timdal & Holtan-Hartwig, 1988), while the latter contains barbatic acid (Möller & Timdal, personal observation).
Rehmia furfurosa was first recognized in 1933, but as a form (f. granulosum) under Rhizocarpon obscuratum, a name sensu auct. is now considered a synonym of R. reducta (= Rhizocarpon reductum) (Ihlen, 2004; Schade, 1933). The placement of this form was based on the hypothesis that R. reducta is the most common species in the genus, displaying adaptability to different conditions and snail grazing. According to Schade (1933), this form was characterized by a dull brown (“sordide fuscis”) thallus, which could be thick or thin, areolate or verrucose, with the areoles or warts being completely granulose, and apothecia with gyrose discs. In a subsequent study, additional characteristics were added: apothecia up to 1 mm in diameter with grooved discs in later stages and ascospores typically featuring three transverse septa and one longitudinal septum, measuring 14–24 × 7–12 µm (Schade, 1935). Importantly, Schade (1935) emphasized that ascospores are often underdeveloped, and f. granulosum represents both a mature form and a stunted condition derived from f. reductum. However, Poelt (1955) noted that the smaller number of cells (4–6) is a diagnostic feature, and the presence of vegetative propagules, which are absent in other brown species of Rhizocarpon s.l., suggests that this taxon should have a species rank. However, Feuerer (1991), after examining the type specimen, could not determine whether Rehmia furfurosa was simply a damaged form, as Schade (1933, 1935) proposed, or a distinct species, as Poelt (1955) argued.
All specimens from Norway and Poland that were analyzed using molecular methods formed a strongly supported clade (PP = 100), including specimens with muriform or submuriform ascospores (Figure 2). The results indicate that, regardless of whether the ascospores are muriform or submuriform, they belong to the same taxon (R. furfurosa). This suggests that fully developed ascospores are muriform and larger. The Polish specimens also had a taller hymenium, up to 150 µm, whereas in the type specimen it was only 80–100 µm high, suggesting that the apothecia in the type material may not have been fully mature (Poelt, 1955). In addition, this lineage is genetically distinct from all R. reducta sequences deposited in GenBank, supporting Poelt’s (1955) conclusion that R. furfurosa is indeed a separate species.
Most lichen species can produce both vegetative propagules and sexual structures. One such example is Lobaria pulmonaria (L.) Hoffm., which can produce both vegetative propagules (soralia and isidia) and apothecia (Smith et al., 2009). In L. pulmonaria, the development of sexual structures depends on the life stage of the thallus: juvenile specimens produce vegetative propagules, whereas mature specimens develop apothecia. However, both micro- and macroclimatic factors influence the type of reproduction (Martínez et al., 2012). Macroclimatic conditions appear to be more important for sorediate morphs, whereas tree-level microclimatic factors appear to be more important for isidiate morphs. In the case of L. pulmonaria, apothecia are more abundant at the base of trees and on northern exposures, which are interpreted as the wettest surfaces (Martínez et al., 2012).
Rehmia furfurosa typically thrives in sunny environments but can persist in more sheltered sites (Purvis & James, 1985; Schade, 1935). In the Colorful Lakelets, specimens were found in partially shaded areas near the lakes, raising the question of whether microclimatic factors, such as light exposure or proximity to moisture, are critical for spore development. In Poland, R. furfurosa was collected in areas where the model predicted habitat suitability ranging from low to moderate. In contrast, sterile or submuriform-spored specimens from Czechia, Austria, France, and Germany were found in areas with moderate to high-suitability. As R. furfurosa was found in Poland under less favorable conditions, it would be valuable to investigate whether the concentration of different metals significantly affects the energy expenditure for apothecia production and, consequently, the full development of spores in the future. However, this hypothesis was not tested in this study, as precise measurements of metal concentrations in the substrata for each sample would be required.
In addition, the potential role of seasonality in ascospore development warrants further investigation. For example, another representative of Rhizocarpaceae, Rhizocarpon lecanorinum Anders, displays seasonal ascospore discharge, with a higher number of ascospores released in winter and early spring, and then a decrease in summer and autumn. In addition to spores collected from January to May, many ripening and mature asci were observed. Conversely, during the remaining months, the asci contained fewer ascospores, many of which showed aborted septal development (Clayden, 1997). Honegger (1978) also noted that Rhizocarpon ascospores develop well in winter or occasionally in early summer, whereas many are destroyed during the off-season. Moreover, optimal development of ascomata in Rhizocarpon occurs when the thalli remain moist for extended periods of time (Honegger, 1978). Specimens from the Colourful Lakelets were collected in late winter and spring, whereas Nordic specimens were collected from May to October. However, one specimen from the Szczepańska private collection was collected in September. Therefore, it is puzzling whether seasonality and wetter conditions in Poland influence the production of mature spores.
Rehmia furfurosa is typically found in areas rich in copper or iron (e.g., Poelt, 1955; Purvis & James, 1985; Schade, 1933, 1935). Despite the lack of studies on metal deposition in the substrates of R. furfurosa or the bioaccumulation of metals within the thalli or apothecia, the model presented in this study (Figure 6A) was based solely on predictors presumed to affect distribution. Interestingly, elevated concentrations of other metals have also been detected at known localities for this species. For example, in Erzgebirge (Saxony, Germany), where Schade (1933, 1935) collected specimens, there is a notable concentration of ores, including silver, lead, zinc, copper, tin, uranium, cobalt, and fluorite (Seifert, 2008 and references therein). Therefore, a model that incorporates a wider range of metals may better explain the distribution of R. furfurosa. However, because GBIF data also include records based on human observations, future studies incorporating a greater number of preserved specimens and employing standardized identification methods may yield more robust and reliable results. The final species distribution map was generated based on the concentrations of seven metals: Fe, Pb, Zn, Al, Cd, Si, and Co (Figure 6A).
According to Poelt (1955), R. furfurosa can be found in areas with higher copper concentrations, but all tests (Figure 4) showed that copper was less significant than other factors. Only in permutation importance did copper (≈11.3%) emerge as relatively significant, similar to iron (≈11.5%). All selected metals (except Si and Al) are known to accumulate in the thalli of other Rhizocarpaceae species (Lange & Ziegler, 1963; Upreti & Pandey, 2000). Moreover, higher iron concentrations were detected in areas where R. furfurosa was found (Purvis & Halls, 1996) in Great Britain. In addition to iron, lead also had a strong influence on occurrence rates. Some lichens, such as Myriospora smaragdula (Wahlenb.) Nägeli ex Uloth (≡ Acarospora smaragdula (Wahlenb.) A. Massal.) can accumulate Pb, along with other metals such as Zn, Cu, Sn, Fe, and Al, in their thalli (Purvis et al., 2000).
According to the model presented in this study (Figure 6A), Rehmia furfurosa is likely to have a wider distribution. In particular, the northern part of the Carpathian Mountains shows moderately suitable areas, indicating that there may be more potential sites for this species in Poland than currently known. In general, the highest probability of occurrence is on mountain massifs, where heavy metals are typically found as oxides in ores and rocks (Figure 6B). However, in Europe, especially in both historical and modern industrial and mining regions, metal concentrations are often elevated because of human activities (Ciepał, 1999; Tóth et al., 2016). For example, in the Colorful Lakelets, metal concentrations have increased as a result of anthropogenic activities linked to pyrite mining, which began in 1785. Mining operations took place at the ‘Hoffnung’ pyrite mine, where the Purple lakelet (also known as Orange or Red Lake) and the Yellow lakelet (also known as Yellow Pond) are now located; at ‘Neues Glück’, where the Blue lakelet (also known as Emerald, Turquoise, Azure) is situated, and at ‘Gustav Grube’, in the area of the Green lakelet (also known as Green Pond, Black Lake). Metallurgical activity in the Landeshut Ridge, where this region is located, dates back to the 14th century (Dziekoński, 1972). Kierczak et al. (2013) identified three types of slag in the Landeshut Ridge: porous slag and two massive slags, equilibrated and undercooled, which contain high concentrations of metals and metalloids such as Cu, Zn, Pb, and As. These elements can leach into the environment and cause metal contamination in soils, stream sediments, and surface waters. This region is also characterized by higher concentrations of Si and Al.
The occurrence of Rehmia furfurosa is restricted to Europe, but it was recently reported from Turkey by Yazici & Aslan (2016), confirming its presence in Asia. However, the description and accompanying photograph of the specimen in their publication suggest that it may be a different species. In detail, one of the two specimens was found on calcareous rock, which is unusual as the genus Rhizocarpon s.l. typically grows only on siliceous rock (Fryday et al., 2024). In addition, the specimens were not tested using thin-layer chromatography methods, and the medulla reacted positively with PD, turning yellow rather than orange, as would be expected if stictic acid were present (Elix, 2014; Fryday et al., 2024). In light of the discrepancies between the specimens from Turkey and our findings, the occurrence of this species outside Europe appears to be doubtful. Therefore, it warrants further verification through additional methods, particularly considering the potentially greater morphological variability.
. Conclusion
Here, we present the first record of Rehmia furfurosa in Poland (Western Sudetes, Colourful Lakelets), together with the ITS sequences, a DNA marker that serves as a universal barcode for fungi. Some of the collected specimens were characterized by muriform ascospores. Despite variations in spore size and greater septation, the specimens belong to the same genetic lineage as Nordic populations, which exhibit submuriform, smaller ascospores, as described in the protologue. In addition, because of the lack of nomenclatural type of Rhizocarpon obscuractum f. granulosum (synonym), a lectotype is here designed. None of the examined specimens produced norstictic acid, suggesting the need for further investigations. Moreover, the underlying reasons for the complete development of ascospores remain unclear, necessitating further examination. The occurrence of this species outside Europe also requires verification, particularly in light of an anomalous report from calcareous rocks in Turkey. It is likely that this species has a wider distribution, possibly extending into the Carpathian Mountains in Poland.
. Taxonomy
Rehmia furfurosa (H. Magn. & Poelt) E .J. Möller, Timdal, Haugan, Bendiksby, Fungal Syst. Evol. 16: 224 (2025);
≡ Rhizocarpon furfurosum H. Magn. & Poelt;
in Poelt, Verh. zool.-bot. Ges. Wien 95: 110 (1955); type: AUSTRIA: Hohe Tauern: Hänge an der Schwarzen Wand bei Hüttschlag in der Großarl [auf kupferhaltigem Gestein], ca. 1600 m, July 1955, J. Poelt (M 0024899! – holotype). Chemistry: stictic acid.
= Rhizocarpon obscuratum f. granulosum Schade, Abh. naturwiss. Ges. Isis: 158–159 (1933); lectotype: [GERMANY] Erzgeb.: Freiberg, auf Haldensteinen beim Gottvertrauten-Daniel-Erbstollen, April 1933, Dr. A. Schade (Schade, Stolle, Riehmer, Lich. sax. exs. No. 501.). (GLM L-0021351! lectotype, selected here). Chemistry: stictic acid and cryptostictic (trace).
MycoBank Typification MBT 10028165
Thallus: crustose, dull ochre-brown to brown, often with violet tinged blastidia; apothecia: lecideine, infrequent, up to 1 mm, flat or weakly convex, sometimes umbonate; disc: reddish-brown to black; proper margin: persistent, thick, internally dark brown and pale brown in the inner part; epihymenium: yellowish brown to dark brown, K+ yellow or partly K+ red-brown; hymenium: hyaline, up to 150 µm; hypothecium: reddish-brown to brown, K- or partly K+ red-brown, HNO3+ orange-brown, HCL+ reddish-brown, with Arnoldiana-brown; paraphyses: with brown caps reacting K+ blackish; ascospores: 8 per ascus, submuriform (up to 8 cells) to muriform (up to 18 cells), 10–29 × 6–17 µm, hyaline.
Chemistry
Stictic acid (major compound), cryptostictic acid (trace), and sometimes constictic acid (trace) in thallus.
Notes
Schade (1933) did not indicate a type specimen of Rhizocarpon obscuratum f. granulosum. Nevertheless, because the name was published before January 1, 1958, the publication is valid (Turland et al., 2018). According to Poelt (1969), forma granulosum is a synonym of R. furfurosa, but the author did not provide any information about the tested specimens. Schade (1933, 1935) included information about the localities of the taxon; unfortunately, most of his collections were destroyed during World War II. After the war, Schade started to rebuild his collection, which is now deposited in the Natural History Museum at Görlitz (part of the Senckenberg Museum) (Kärnefelt et al., 2012). However, in 1920s, he published exsiccates, which included specimens of R. obscuratum f. granulosum (Schade, F. Alvin, Stolle, Emil & Riehmer, Ernst, Lichenes Saxonici Exsiccati. Dec. I-LV, nos. 1–550, collected 1923–1925). Only two specimens with numbers 501 (labelled as Rhizocarpon obscuratum (Ach.) Mass. nov. fo. granulosum Schade) have been found: in FH (Farlow Reference Library and Herbarium of Cryptogamic Botany, Harvard University) and GLM (Senckenberg Gesellschaft für Naturforschung: Senckenberg Museum für Naturkunde Görlitz).
In the protologue, Schade (1933) provided information on the locations where he found R. obscuratum f. granulosum, including Mld. [Muldenland] Gottvertrauter Daniel-Erbstolln (sehr zahlreich, wird ausgegeben in Schade, Stolle, Riehmer Lich. Sax. Exs. N. 501) or U. Ezg. [Unteres Erzgebirge] Reicheltschacht (“loc. class.” Sehr zahlreich). Unfortunately, a specimen with a locus classicus was not found; therefore, an exsiccate number from the GLM herbarium was selected as a lectotype.
Additional exsiccata specimens examined
Poelt, Steiner; Lich. Alp. [München (Munich)]: No. 225, Ullrich (M-0330570)
Specimens examined
Czechia: Central Bohemia – distr. Rakovník, Křivoklátsko Protec. Landsc. Area; Skřivaříš – Valachov Natural Monument, W-exposed forested slopes above Tyterský brook, area of main drift mine called “Valachovské jeskyně” cave, alt. 350–380 m, on siliceous iron-rich rock, 07.02.2009, J. Malíček 1546, (Herb. Malíček); Central Bohemia – distr. Příbram, Dobříš region; Nový Knín – W-exposed rocky slopes above “Václavova štola” gallery, in Kocába Brook valley 1 km NW of town, alt. 290–320 m, on iron-rich siliceous rock, 10.04.2009, J. Malíček 1648 (Herb. Malíček); Silesia – dist. Opava, Vítkov – Zálužně: old schist quarry 600 m NNW of settlement, 49.827222°N, 17.717778°E, alt 430–450 m, on Fe-enriched slate rock, 28.04.2010, J. Malíček 3181 et al. (Herb. Malíček); Central Bohemia – distr. Rakovník, Křivoklátsko Protected Landscape Area, Roztoky – U Eremita Nature Reserve, 50.016472°N, 13.860639°E, alt. ca. 250 m, on Fe-enriched schistose rock, 05.06.2010, J. Malíček 3215 et al. (Herb. Malíček); France: Counozouls: au niveau de la petite cascade, alt. 1081 m, sur schistes non calcaires, métalifères, paroi quasi verticale, 13.09.2016, S. Poumarat, (Herb. Poumarat – SP 2016-85); Germany: Unt.[eres] Erzgeb.[irge], Brand-Erbisdorf auf Granitblöcken an der N[ord]-Seite der Halde am Vertrau auf Gott Schacht, 500 m, 29.04.1934, W. Flößner (M-0330569); Norway: Akershus, Eidsvoll: base of Mt Byrudshøgda along road 33, ca. 900 m, N of Byrud, 60.421333°N, 11.192833°E, alt. 170 m, on rust-stained rock by the road, 14.06.2012, M. Bendiksby, E. Timdal & M. Westberg 12516 (O L-179440); Akershus, Eidsvoll: c. 1 km N of Byrud, 60.4214°N, 11.1884°E, alt. 180 m, 31.10.1986, J. Holtan-Hartwig, E. Timdal 4816 (O L-152570); Hedmark, Alvdal: Mt Tronfjellet, the old copper mine Gruvvangen, 62.14°N, 10.656167°E, alt. 750 m, on rock wall, 27.07.2012, S. Rui & E. Timdal 12711 (O L-179949); Hedmark, Alvdal: Mt Tronfjellet, the old copper mine Gruvvangen, 62.14°N, 10.656167°E, alt. 750 m, on rock wall, 27.07.2012, S. Rui & E. Timdal 12709 (O L-179947); Hedmark, Alvdal: Strømmen, site of the old copper smelting works Strøms hytte, 62.1968°N, 10.6251°E, alt. 485 m, on slag in slag heap, 05.08.2023, E. Timdal 20713 (O L-239319); Nordland, Saltdal: Evenesdalen, along the old road, 66.944667°N, 15.396667°E, alt. 190 m, rock wall facing N, 08.08.2010, E. Timdal (O L-163730); Oppland, Sel: Åsårgruva historical copper mine (in operation in the 17th century), 61.7912°N, 9.4347°E, alt. 335 m, rock in an old mining heap, 03.06.2023, R. Haugan, E. Möller, O. J. Nørstegård, A. Simon, E., E. Timdal, R. Zane 2309662 (O L-243115); Oppland, Vågå: Storrvikhøygarden; 61.85°N, 8.85°E, alt. 400–500 m, iron-rich rock close to an old road, 07.10.1997, R. Haugan 5955 (O L-166436); Oppland, Østre Toten: N of Jonsrud, 60.6095°N, 11.121°E, alt. 140 m, iron-stained rock wall along the road, 27.05.2011, R. Haugan & E. Timdal 9908 (O L-173244); Oppland, Østre Toten: along road 33 at the W shore of Lake Mjøsa, N of Jonsrud, 60.6095°N, 11.121°E, alt. 130 m, on a rust-colored rock wall by the road, 27.05.2011, R. Haugan & E. Timdal 11856 (O L-169764); Oppland, Østre Toten: along road 33 at the W shore of lake Mjøsa, S of Langsletta, 60.6094°N, 11.1213°E, alt. 130 m, on shady rock wall, 04.06.2023, E. Timdal 20479 (O L-239088); Oppland, Østre Toten: along road 33 at the W shore of lake Mjøsa, S of Langsletta, 60.6195°N, 11.114833°E, alt. 140 m, 27.05.2011, R. Haugan & E. Timdal 11858 (O L-169766); Oppland, Østre Toten: along road 33 between Jonsrud and Langsletta, 60.61°N, 11.120833°E, alt. 150 m, steep, iron-stained rock wall by the road, 16.09.2012, E. Timdal 12802 (O L-182027); Sogn og Fjordane, Luster: Gaupne, Meinsete, 61.3908°N, 7.3256°E, alt. 15 m, mineral rich rock in roadcut, 22.07.2024, R. Haugan, E. Möller 2417720 (O L-244052); Sogn og Fjordane, Luster: Gaupne, Meinsete, 61.3908°N, 7.3256°E, alt. 15 m, mineral rich rock in roadcut, 22.07.2024, R. Haugan, E. Möller 2417722 (O L-244114); Sør-Trøndelag, Oppdal: Drivdalen, NW of Drivstoggosætra, 62.3571°N, 9.6322°E, alt. 750 m, steep rock along the road, 09.07.2011, R. Haugan 10302 (O L-173852); Poland: Rudawy Janowickie, S of Wieściszowice, Colourful Lakelets (Polish: Kolorowe Jeziorka), artificial ponds formed in place of former mines at the slope of Wielka Kopa Mt., outcrop by Purple Lakelet (Polish: Purpurowe Jeziorko), alt. 560 m, 50.828514°N, 15.973539°E, mining site with metalliferous rocks, 27.04.2023, M. Kukwa 24664 & 24677 (UGDA L-64403 & 64416); ibidem, rocky slope by Blue Lakelet (Polish: Niebieskie Jeziorko), alt. 645 m, 50.823519°N, 15.972457°E, mining site with metalliferous rocks, 27.03.2023, M. Kukwa 24682 & 24683 (UGDA L-64421 & 64422); ibidem, former mine excavation N of Blue Lakelet (Polish: Niebieskie Jeziorko), alt. 645 m, 50.824061°N, 15.972842°E, mining site with metalliferous rocks, 27.04.2023, M. Kukwa 24703 & 24707 (UGDA L-64442 & 64446); ibidem, artificial boulder scree in the place of mine excavation S of Red Pond (Polish: Purpurowe Jeziorko), alt. 575 m, 50.826807°N, 15.973366°E, 27.03.2023, R. Szymczyk s.n. (UGDA L-65531); ibidem, by Colorful Lakelets, alt. ca. 550 m, on rock, 25.09.2022, K. Szczepańska 1354 (herb. Szczepańska).
. Supplementary material
The following supplementary material is available for this article:
Supplement S1. Phylogenetic Trees, Posterior Estimates, MCMC Diagnostics, and Species – Metal Response Curves.
Acknowledgements
We would like to thank the Curators of M and GLM herbarium, Jiří Malíček and Serge Poumarat for the loan of Rehmia furfurosa specimens, and Magdalena Kosecka for her assistance in identifying an appropriate molecular isolation method and for her valuable suggestions. We are also grateful to the two anonymous reviewers for their valuable comments.
