. Introduction
Impatiens parviflora DC., commonly known as small balsam, is an invasive plant native to the mountains of central Asia. Intentionally introduced for botanical reasons, this species turned out to be one of Central Europe’s most widespread invasive plants (Langmaier & Lapin, 2020; Tanner et al., 2015a). Despite the fact that the factors affecting its spread are still debatable and unclear, it was found that it is able to penetrate into species-rich habitats, which makes it a potential threat to biodiversity (Adamowski & Bomanowska, 2016; Florianova & Münzbergova, 2018).
Most scientific reports concern the invasive species Impatiens glandulifera Royle, known as Himalayan balsam, which is invasive throughout Europe and can also seriously reduce native plant diversity (Ab Razak et al., 2023; Coakley & Petti, 2021; Tanner et al., 2015a; Tanner & Gange, 2020). It negatively affects whole ecosystems where the native biodiversity of invertebrates and microbial communities is disturbed (Coakley & Petti, 2021).
Much less is said about Impatiens parviflora, although it has been noted in 34 European countries, as reported already in 2008 by Lambdon et al. (2008). It has been observed at many sites in European temperate broadleaf forests (Langmaier & Lapin, 2020), such as the Czech Republic (Florianová & Münzbergová, 2017; Hejda, 2012) and Slovakia (Jarčuška et al., 2016) or Lithuania (Krokaitė et al., 2022).
The first localities of I. parviflora in Poland were recorded in 1850 in northern regions near Gdańsk (Krawiecowa, 1951), and since then, this species has spread very expansively (Piskorz & Klimko, 2006). It occurs, among others, in Polish legally protected areas such as Wigry National Park (Pusz et al., 2020), Kampinos National Park (Adamowski & Bomanowska, 2014), Wielkopolski National Park (Piskorz & Klimko, 2006, 2007), or Białowieża National Park with Białowieża Primeval Forest (Adamowski & Keczyński, 1998; Chmura & Sierka, 2007). However, it should be expected in the future that the rate of invasion of I. parviflora may decrease in our country as a result of occupation of almost the entire ecological niche.
The ability of small balsam to tolerate less favorable conditions under a dense tree canopy gives it an advantage over other species (Langmaier & Lapin, 2020; Lanta et al., 2022), although, as noted by Bobulská et al. (2016) I. parviflora can also develop in diverse conditions because sunlight and soil moisture do not have a significant impact on its population. It may, therefore, be a threat to the Impatiens noli-tangere, which is more sensitive to conditions and native to Europe, which was noted in Belgium (Godefroid & Koedam, 2010; Vervoort & Jacquemart, 2012). Glushakova et al. (2015) found that I. parviflora leads to a serious decrease in biodiversity of the plant species. It also influences the structure of soil microbiocenoses (Chmura & Gucwa-Przepióra, 2012).
Mechanical control of small balsam is tedious (Adamowski & Bomanowska, 2016). There is some hope for stopping the expansion of invasive Impatiens by using pathogens or pests that limit their populations. A possible biological weapon against small balsam may be rust fungus Puccinia komarovii Tranzschel ex P. Syd. & Syd. (Piskorz & Klimko, 2006; Tanner et al. 2015b; Rolnikuj.pl, 2017) which has been a parasite of I. parviflora and Impatiens amphorata in Central Asia and the northern Himalayas. It probably came to Europe with I. parviflora seeds in the early 20th century. In Poland, where it is considered a common species, this rust was reported for the first time in the area of Silesia and Beskidy in 1934, while in other European countries is treated as invasive (Majewski, 1979; Piskorz & Klimko, 2006). According to the observations of Tanner et al. (2015b), in Europe, it infects I. parviflora mainly in wooded habitats, and the percentage of infected plants may reach even 90% (Piskorz & Klimko, 2006).
Puccinia komarovii is an autoecious rust and had been known to be host-specific rust, infecting only Impatiens parviflora in both its native and European ranges, and was described originally from this Impatiens species (Piskorz & Klimko, 2006; Tanner et al., 2015a). It turned out that I. glandulifera can also be its host (Tanner et al., 2015b). However, there is host specificity. Hence, isolates infecting I. parviflora are referred to as P. komarovii var. komarovii, unlike P. komarovii var. glanduliferae inhabiting I. glandulifera (Tanner et al., 2015a).
Due to the above facts, observations of infection severity of I. parviflora by P. komarovii var. komarovii in two Polish national parks - Drawa National Park (DNP) and Wigry National Park (WNP) were carried out. Also, molecular analyses of pathogen isolates were performed to confirm the species identification of the rust fungus, diagnosed on the basis of morphology and symptoms on small balsam as P. komarovii var. komarovii and preliminary characterization of the population of this fungus based on comparative analysis of rDNA region sequences.
. Material and methods
. Field research
The subjects of this investigation comprised I. parviflora specimens grown within Drawa National Park (DNP), situated in northwestern Poland, at the confluence of Greater Poland, Lubusz, and West Pomeranian Voivodeships and specimens from Wigry National Park (WNP), which is situated in the northeastern region of Poland within Podlaskie Voivodeship (Figure 1). At both study locations, 20 plots measuring 10 × 10 m were selected within mixed forests (Table 1) characterized by dense I. parviflora coverage. From each of these locations, 10 plants were chosen for observation, and the dynamics of rust occurrence were monitored at 4-week intervals, commencing from May to June, depending on the growth rate of I. parviflora, and continuing until September. These observations spanned three plant vegetation seasons (2020–2022) in WPN and two seasons (2022–2023) in DNP. Monthly assessments of the percentage of leaf coverage affected by etiological symptoms of Puccinia komarovii var. komarovii were conducted based on an average of 10 plants per plot. Descriptive statistics and graphical representations depicting the dynamics of rust occurrence were generated using Tableau (2020.2.4 Professional Edition).
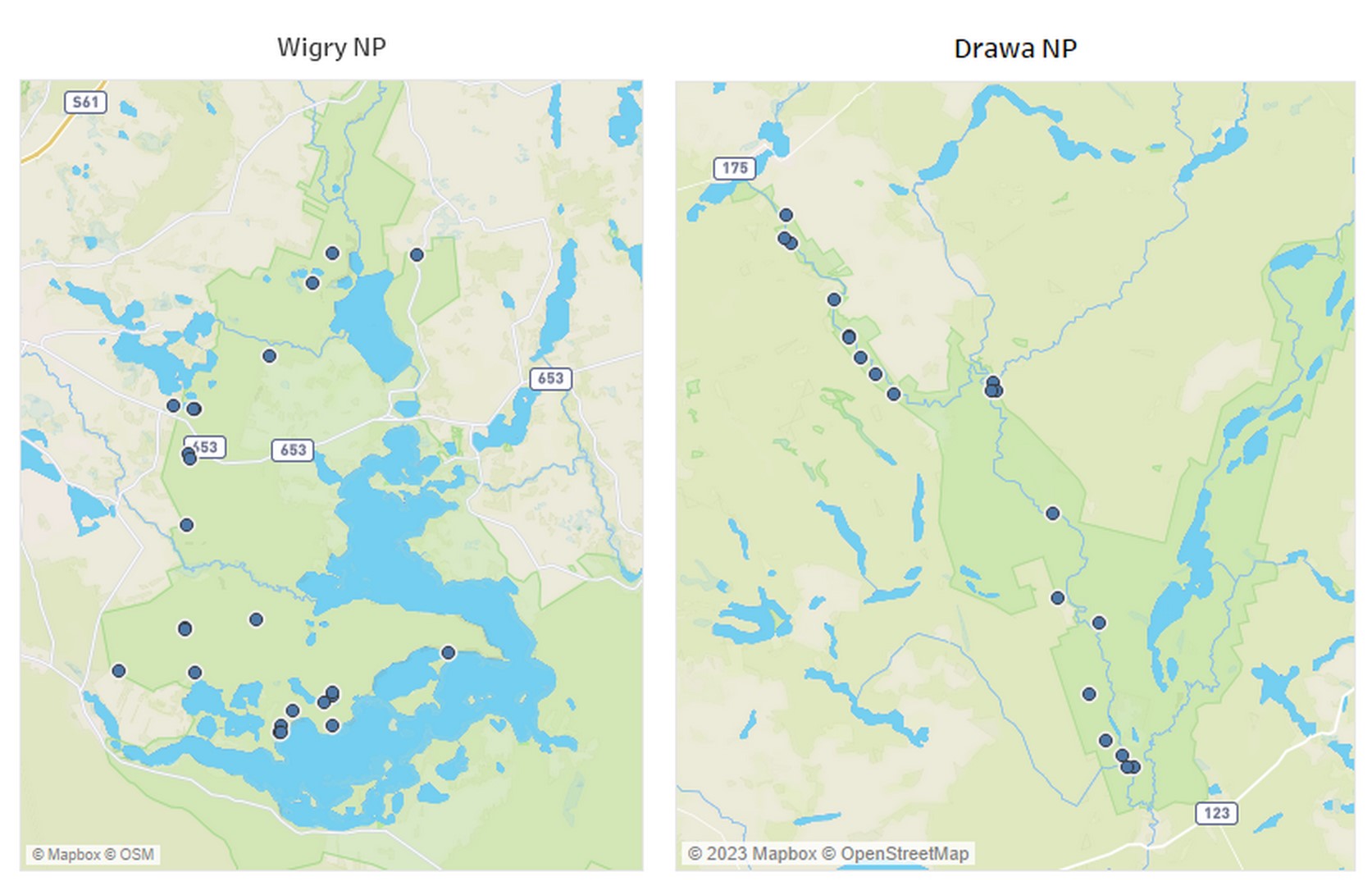
Table 1
Characteristics of sample locations.
. Molecular analyses
The research material for molecular analyses were leaves of I. parviflora with clear disease symptoms indicating infection with the fungus P. komarovii. Material was taken in August because, during this period, the most pronounced symptoms caused by uredinia and telia were observed. Leaves were collected from 14 small balsam sites in 2022, from 11 sites in 2023 in DNP, and from six sites in 2022 in WNP. In this way, 31 samples were obtained, from which 31 DNA samples were obtained for analyses of rDNA regions of P. komarovii var. komarovii (Table 2).
Table 2
A list of 31 P. komarovii var. komarovii isolates originated from Impatiens parviflora in Wigry National Park (WNP) in 2022 and in Drawa National Park (DNP) in 2022 and 2023 with accession numbers of their ITS and LSU sequences deposited in GenBank NCBI.
. DNA extraction and PCR assays
Leaf fragments with disease symptoms were freeze-dried in a CoolSafe device (ScanVac, Denmark). From symptomatic leaves collected at 31 sites, small fragments measuring 25–49 mm2 with numerous clusters of spores were cut out and homogenized to fine powder with quartz beads in a MagnaLyser homogenizer (Roche, Switzerland). DNA extracted from forty-gram samples using the Bead-Beat Micro AX Gravity kit (A&A Biotechnology) was measured fluorometrically on a Quantus device (Promega, USA) and diluted to 10 ng⋅µL−1 in ddH2O for further analyses.
Amplifications of the ITS (internal transcribed spacer) and 28S (LSU - large subunit) of rDNA were done in 37.5 µL w an Eppendorf EP Mastercycler (Eppendorf, Germany). The final concentration of reagents was as follows: 1× PCR Mix Plus (A&A Biotechnology, Poland), DNA of individual samples at a concentration of 4 ng, and rust-specific primers, respectively ITS5-u/ITS4-u (Pfunder & Schürch, 2001) and LRust1R/LR6 (Beenken et al., 2012; White et al., 1990), each at a concentration of 0.8 pM.
The PCR began with an initial denaturation step of 5 min at 95 °C, followed by 35 cycles of 95 °C for 1 min, an annealing temperature of 53 °C for 50 s and 72 °C for 50 s (ITS) or 90 s (LSU), followed by 10 min at 72 °C for a final extension.
The presence of amplicons was verified by electrophoretic separation of 2 µL of the postreaction mixture in tris-borate-EDTA (TBE) buffer on a 1.2% agarose gel (Pronadisa, Spain) stained with SimplySafe (EURX, Poland) and visualized using the INTAS set (Germany) consisting of a transilluminator, camera and computer with software.
. Sequencing of PCR products and data analysis
Amplification products were purified and sequenced by Genomed (Poland). Both strands were sequenced. FinchTV 1.4 (Geospiza) was used to analyze the obtained sequences.
Alignments were carried out using ClustalW through the Molecular Evolutionary Genetics Analysis Version 11 (MEGA11) Toolbar (Tamura et al., 2021). For species identification based on ITS and LSU sequences, the Basic Local Alignment Search Tool (BLAST) in the NCBI (The National Center for Biotechnology Information) database was used (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome).
The sequences of the analyzed isolates were compared with the P. komarovii sequences available in GenBank NCBI. In the case of the ITS regions, there were 16 sequences of isolates from I. parviflora (seven Polish and nine Chinese), 91 Indian sequences from I. brachycentra and I. glandulifera (respectively seven and 84) and eight Pakistani ones from Impatiens sp.
For comparative analyses of the LSU region, 20 sequences of isolates from I. parviflora (one from the UK, 12 - from Hungary, seven- from Poland), seven from I. brachycentra (all from India), seven from I. glandulifera (all from India) and seven from Impatiens sp. (Pakistan) were taken.
Dendrograms were constructed to visualize the genetic diversity between the 31 P. komarovii var. komarovii isolates tested and the sequences of 44 or 41 P. komarovii isolates for ITS and LSU, respectively, from GenBank NCBI. For technical reasons, the number of Indian sequences of the ITS region obtained from I. glandulifera was limited to 13 differentiated representatives.
The dendrograms were constructed in MEGA11 using the Maximum Likelihood algorithm. The Tamura 3-parameter model was applied (Tamura, 1992).
. Results
. Field research
During field studies, the highest incidence of rust infection on I. parviflora leaves was recorded in DNP, where the median blade infestation for 2022–2023 was 51% of leaf coverage. In contrast, after three years of observation in WNP, the median blade infestation amounted to only 25%. These findings are supported by the yearly average infection rate of I. parviflora in DNP, where the median yearly infection for 2022 was 21%, increasing to 68% in 2023. Notably, in 2022, spot imp3 in DNP showed the highest median rust infection (50%), while most other locations fell within the interquartile range of 13–40%. In 2023, DNP experienced the highest leaf blade coverage of rust in any observation year, with the interquartile range doubling from the previous year, ranging from 55% to 87%. Spot imp6, in particular, consistently exhibited a median 100% coverage of rust throughout the entire year.
In WNP, the median yearly infection never exceeded 50% and, for the years 2020–2023, constituted 14%, 34%, and 16%, respectively. The highest yearly median rust infection in WNP was observed in 2022 at spots imp2 (68%) and imp17 (66%). Infection ranges in 2020 and 2022 in WNP were similar, with interquartile ranges falling between 8–36% in 2020 and 8–43% in 2022. The peak infection in WNP was observed in 2021, with Q1 starting at 22% and Q3 finishing at 44%. This is shown in graphs (Figure 2).
Figure 2
Monthly and yearly average of I. parviflora leaf blades’ coverage by Puccinia komarovii var. komarovii in Wigry and Drawa National Parks in 2020–2023.
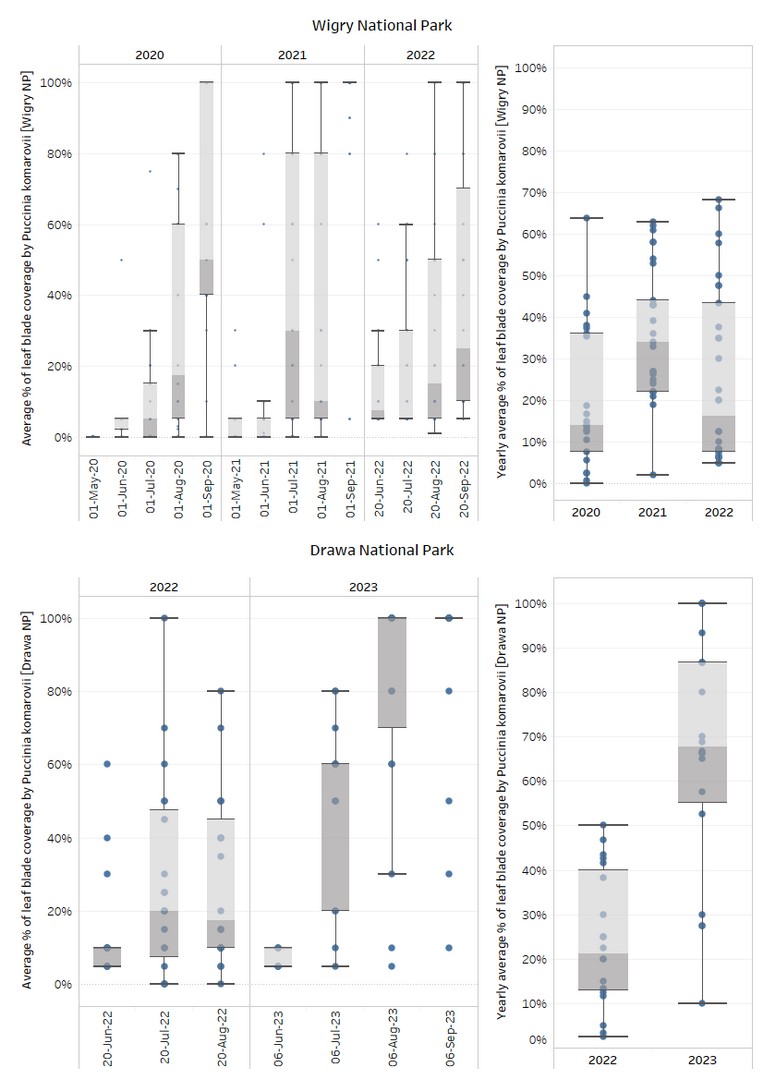
Field studies indicated that the most severe symptoms caused by P. komarovii var. komarovii on I. parviflora leaves occurred during the summer months. However, the peak of rust presence varied between years. In WNP, the median peak of rust presence was observed in September 2020 at 50%, July 2021 at 30%, and September 2022 at 25%. In DNP, the maximum rust presence was observed in July 2022 at 20% and in August 2023 at 100%. In some growing seasons, premature death of I. parviflora was observed, with entire plant populations in certain locations perishing by September. Such instances were documented in both WNP in 2021 and DNP in 2023.
. Molecular analyses
Comparative analysis of the ITS and LSU sequences of the tested samples clearly showed the presence of the rust fungus P. komarovii var. komarovii on the leaves of I. parviflora collected in 31 locations. The sequences of the fungal isolates found in individual samples were deposited in GenBank NCBI. Their accession numbers are listed in Table 2. Access to the sequences in the GenBank database is available at https://www.ncbi.nlm.nih.gov/nucleotide/.
The ITS sequences of the 31 P. komarovii var. komarovii isolates obtained over two years from the DPN and WPN did not differ from each other. Comparative analysis with sequences from I. parviflora available in GenBank NCBI showed that all our sequences showed 100% identity with the sequences of seven isolates - three from central Poland with accession numbers KC430771 KC430772 and KC430775 and four sequences KC430842, KC430839 KC430838 and KC4 30837 from China. One nucleotide distinguished them from the Polish sequences KC430769, KC430773 and KC430774 and the Chinese KC430840, which meant an identity level of 99.84%. They differed by two nucleotides from KC430841, KC430843, and KC430844 from China, and the Polish KC430770 (identity 99.69%), and by four from the Chinese KC430845 (identity 99.37%).
Comparing our sequences with seven sequences obtained from Impatiens brachycentra (spurless balsam), it was found that they differed from single isolates by one (KC430763), three (KC430759) or four nucleotides (KC430758), which corresponded to an identity level of 99.84, 99.53 and 99.37%, respectively. Our isolates differed much more from the remaining four isolates obtained from this species of Impatiens, i.e., by 8–10 nucleotides, with an identity level of 98.76–98.29%.
The similarity of P. komarovii sequences obtained from I. glandulifera to the ITS sequences of our isolates was even lower. The sequences differed by at least six nucleotides and by a maximum of 14 nucleotides, which resulted in an identity of 99.07–97.83%.
However, the greatest differences were found in Pakistani P. komarovii isolates from the unknown Impatiens species. Our isolates were similar to them only by approximately 88%.
Dendrograms based on ITS region sequences show relationships between our P. komarovii isolates and GenBank NCBI isolates from I. parviflora (Figure 3) and between our isolates and P. komarovii isolates from Impatiens species other than I. parviflora (Figure 4).
Figure 3
Dendrogram based on ITS sequences showing the relationships between the studied 31 Puccinia komarovii var. komarovii isolates and 16 P. komarovii var. komarovii isolates from GenBank NCBI derived from I. parviflora.

Figure 4
Dendrogram based on ITS sequences showing the relationships between the studied 31 Puccinia komarovii var. komarovii isolates and 28 P. komarovii isolates from Impatiens species other than I. parviflora.

Also, the sequences of our 31 isolates encoding LSU did not differ from each other. However, they did not show 100% identity with any Polish isolate of I. parviflora from Greater Poland Voivodeships available in GenBank NCBI. They were identical to the sequences of the KC460259 isolate from Hungary and LC494104 from the United Kingdom.
The tested isolates differed from the sequences of the Polish isolate KC460243 and the Hungarian KC460260 by one nucleotide, which was equivalent to a degree of identity of 99.91%, or by two nucleotides in KC460254 and KC460256 from Hungary and KC460239 from Poland, with a degree of identity of 99.81%. In the case of the remaining Polish sequences in GenBank, the diversity was greater and concerned three (KC460240) to seven (KC460244) nucleotides, and in the case of the remaining Hungarian sequences ranged from three (KC466553 and KC460255) to ten (KC460263) nucleotides. Therefore, for the LSU region, greater diversity between Polish isolates was found in the ITS regions.
Comparisons of our LSU sequences with P. komarovii isolates derived from other species Impatiens than I. parviflora showed a large range of diversity. Our isolates differed by two nucleotides from only one sequence of P. komarovii (KC460250) originating from the Indian I. glandulifera. From the remaining six sequences derived from this Impatiens species, available in GenBank - by four or even seven nucleotides. Comparing our isolates with sequences of Indian isolates derived from I. brachycentra, the differences were even greater and ranged from four to 12 nucleotides. Pakistani isolates originating from Impatiens sp. differed from our isolates by three nucleotides and even 14 ones (identity 98.71%).
The relationships between the LSU sequences described above are graphically presented in dendrograms, which, similarly to the ITS sequences, show relationships between our P. komarovii isolates and I. parviflora isolates from GenBank NCBI (Figure 5) and between our isolates and P. komarovii isolate derived from Impatiens species other than I. parviflora (Figure 6).


. Discussion
Most scientific reports concern the invasive species Impatiens glandulifera (Ab Razak et al., 2023; Coakley & Petti, 2021; Tanner et al., 2015a), which is infected by P. komarovii var. glanduliferae. This rust fungus causes significant damage of infected plants at the seedling stage (that usually leads to plant death) and leaves of maturing plants (Varia et al., 2016).
Evans and Ellison (2005) emphasize that understanding the biology of a pathogen that may be a potential biological control agent before its large-scale introduction into an ecosystem is an essential element of any weed biocontrol program. It is equally important to prove its specificity for the unwanted, problematic plant species that is our target (Barton, 2012). According to Tanner et al. (2015a), in the P. komarovii population, there are several pathotypes specific to a particular plant species. I. glandulifera shows immunity to inoculations with P. komarovii obtained from I. parviflora and vice versa. Therefore, host specificity predisposes P. komarovii var. glanduliferae to the role of fungal agent for the biological control of Himalayan balsam (Tanner et al., 2015a). For this reason, this rust fungus has been introduced into the UK for biological control of this weed, where was the subject of an intensive biological control research program (Currie et al., 2020; Varia et al., 2016). Ellison et al. (2020) revealed the first field results that clearly showed the potential for successful biological control of Impatiens glandulifera.
Similar hopes are associated with P. komarovii var. komarovii in relation to I. parviflora (Pusz et al., 2020). Already 17 years ago, Piskorz and Klimko (2006) reported on the rapid reproduction of Impatiens in Poland and other Central European countries. This is facilitated by its high adaptability to temperature, humidity, soil reaction, and nutrients (Reczyńska et al., 2015). Observations carried out in the Czech Republic by Florianová and Münzbergová (2018) showed an almost twofold increase in the intensity of the occurrence of I. parviflora on the observed plots in the period 2011–2016. These studies confirmed that I. parviflora tolerates acidic soil reactions and is tolerant (except for seedlings) to herb layer cover. This is consistent with the work of Lanta et al. (2022), and Langmaier and Lapin (2020), who summarized that the successful reproduction of I. parviflora is due to tolerating less favorable conditions under a dense tree canopy and thus avoids competition by this shade-tolerant plant species. However, seedlings are more sensitive, which also results in susceptibility to pathogens. Bacigálová et al. (1998) reported up to 100% mortality of this weed seedlings infected with aeciospores of P. komarovii var. komarovii. P. komarovii var. komarovii holds promise concerning its relationship with I. parviflora, as previously suggested by Pusz et al. (2020). Our observations have revealed a substantial infection of small balsam leaf blades by this rust, occasionally resulting in symptoms manifesting on 100% of the leaf surface, leading to premature I. parviflora mortality and population decline by September. Unfortunately, we did not observe an annual escalation in the occurrence of rust driven by inoculum growth in the environment. The percentage of leaf coverage by P. komarovii var. komarovii exhibited fluctuations, depending on the year, implying an influence of environmental conditions that stabilized the infection each year. As a result, a consistent increase in its occurrence or a fixed peak of infection was not observed; both parameters varied annually and varied on every spot. Therefore there appears the need to understand the genet diversity of P. komarovii var. komarovii population occurring in Poland, which might predispose its usage as a biological agent against small balsam.
As noted by Pollard et al. (2021) on the occasion of research on the biological control of Himalayan balm, for successful control, it is essential that plant biotypes are matched to the most virulent rust strains. Strains from the native range may be useful, among which it is reasonable to distinguish those to which local biotypes will not be resistant. Although in the case of I. glandulifera control in the UK, the most effective strain was Puccinia komarovii var. glanduliferae from India (Pollard et al., 2019), local strains can also potentially naturally limit weeds, whose expansiveness can be particularly dangerous in legally protected areas, where the possibilities of human intervention aimed at controlling the spread of plants are very limited. For this reason, it is advisable to monitor the dynamics of the development of P. komarovii var. komarovii in Poland, observing its genetic variability and impact on the I. parviflora population.
Due to the host-specificity described by Tanner et al. (2015a), there is no danger that this rust will pose a threat to touch-me-not balsam (Impatiens noli-tangere) native in Poland. No symptoms of P. komarovii have been observed on I. noli-tangere during three years of research conducted by us in DNP and WNP (data not published). Also, there is no information about such infections in the available literature.
The P. komarovii var. komarovii isolates analyzed in this study did not show any genetic differentiation within the analyzed rDNA regions, but they differed from the sequences of isolates from the Great Poland Voivodeship available in GenBank NCBI. In the case of ITS regions, this difference was a maximum of two nucleotides, while in the case of LSU - even seven ones. Greater differences were found between our isolates and isolates derived from I. glanduliferae and I. brachycentra, an Asian species which, unlike I. parviflora and I. glandulifera, do not occur in Poland (POWO, 2023). This is consistent with the observations of Tanner et al. (2015a), who observed variation in both ITS and LSU sequences of the rust-infecting I. glandulifera compared to accessions on other hosts.
Our research indicates some differentiation of P. komarovii var. komarovii in a relatively small area, i.e., in Poland. This genetic diversity probably results from sexual reproduction and may enable rapid adaptation of the pathogen to the region or host plant. Adaptation of pathogens to plant species is often due to the generation of novel allele combinations (Covo, 2020). Or it may indicate certain “entry points” to Poland where the isolates arrived from their native place of occurrence. While in the case of crop plant pathogens, genetic diversity means a threat because it may be related to the mentioned adaptive abilities, in the case of invasive plant pathogens, the development of which we would like to limit, genome instability seems to be desirable, because the pathogen has a greater chance of overcoming the possible resistance of the host plant and adaptation to the environment.
