. Introduction
Allelopathy is the phenomenon by which plants release chemical compounds into the environment that positively or negatively affect the growth and population of neighboring plants through various processes (leaching, root exudation, volatilization, residue decomposition, etc.) (Koocheki et al., 2013; Rice, 1984). A detailed understanding of the mechanisms of plant-to-plant allelopathic interactions in ecosystems can offer many implications and applications that help in developing new environmentally safe biological control strategies for sustainable farming (Loydi et al., 2015; Meiners et al., 2012). Currently, crop performance, which is suppressed by allelopathic weeds, has received more attention in order to control weeds in farming systems. In comparison to crops, weeds have the ability to grow faster through their rapid reproduction strategies and production of large amounts of biomass in a short period; weeds are also capable of tolerating environmental stress and can modify seed characteristics with respect to seedling establishment (Qasem & Foy, 2001).
Food security is an important issue for all countries, and increased attention to crop protection and weed management is required for sustainable development. Many agricultural lands in developing countries are infested with various native plants and invasive weeds (Akobundu, 1992), and weed control is a major problem for Asian farmers. Weed–crop interactions include the release of allelochemicals by weeds, which may affect seed germination and crop growth (Rafiqul Hoque et al., 2003).
In Asian countries, rice is the predominant staple food crop and provides a high amount of carbohydrates in the diet (Boers et al., 2015). Farmers in West Bengal are interested in cultivating the popular aromatic rice cultivar ‘Gobindobhog’ for its good demand from both domestic and international markets. However, many of the crops of indigenous aromatic varieties are facing weed incursions that may lead to their extinction as a result of the cultivation of only the commercially viable varieties. Weed infestation of rice crops is a critical problem that not only decreases the rate of production, but also affects both Indian and international market values. Members of the Lamiaceae family contain phytochemicals that affect the growth of other plant species in the environment (Lovett & Weerakoon, 1983). Lamiaceous weeds are a constant constraint on rice production, leading to yield reduction (Zoschke, 1990). Various weeds are found in Oryza sativa ‘Gobindobhog’ fields, but weeds such as Hyptis suaveolens and Leucas cephalotes are those that are most frequently found.
Hyptis suaveolens (pignut; Lamiaceae) is an annual herb and, because of its rapid rate of emergence, this invasive weed has negative effects on native plant communities and impedes the growth of vulnerable or threatened species (Padalia et al., 2014). Moreover, the germination and seedling growth of several crops are affected by its allelopathic influence (Arzoo et al., 2016; Mominul Islam & Kato-Noguchi, 2013a; Poornima et al., 2015; Rao & Singh, 2015; Rodrigues et al., 2012).
Leucas cephalotes (Roth) Spreng. (guma; Lamiaceae) is an annual herb that generally grows in the rainy season and is commonly found by the roadside and in wasteland and fields (Chauhan, 1999; Khanam & Hasan, 2005). It is also used as an edible vegetable and herbal remedy by tribal people and is mostly found in the fields of kharif (monsoon) crops in different states of India (Sinha & Lakra, 2007).
The aim of the present investigation was to study the allelopathic potential of aqueous root and leaf extracts of H. suaveolens and L. cephalotes on seed germination and early growth of the economically important aromatic Oryza sativa cultivar ‘Gobindobhog.’
. Material and Methods
. Plant Material
Plant material was sampled in rice fields in the region of Santiniketan (23.68° N, 87.68° E), Birbhum District, West Bengal, India. Fresh samples of leaves and roots of H. suaveolens and L. cephalotes were collected during their flowering stage, as these plants release more allelochemicals in the flowering stage than in the vegetative stage (Ahmed & Wardle, 1994). Plants were brought into the laboratory, and fresh roots and mature leaves were immediately separated out, washed several times with tap water, and cut into 2–3-cm pieces. Both leaves and roots were dried in shade at room temperature (20–25 °C) and then ground into powder with a mortar and pestle. Plant samples were stored in autoclaved plastic bags under refrigeration (4 °C) until use in the bioassay. Healthy uniform seeds of Oryza sativa ‘Gobindobhog’ were obtained from Bidhan Chandra Krishi Viswavidyalaya (BCKV), Mohanpur, West Bengal, India. Rice seeds were surface-sterilized with 4% sodium hypochlorite for 5 min and then rinsed with distilled water several times to remove excess chemicals (Jefferson & Pennacchio, 2003) before the germination tests were performed.
. Preparation of Aqueous Leaf and Root Extracts
Dry leaf and root powders of H. suaveolens and L. cephalotes (5 g) were soaked for 24 hr in 100 mL double-distilled water at room temperature (20–25 °C) in dark conditions, and the solution was filtered through one layer of filter paper to remove debris and centrifuged at 1,411 g for 15 min. The supernatant was then filtered through a two-layer of Whatman No. 1 filter paper. The stock solution (5%) was diluted to prepare solutions with concentrations of 4%, 2.5%, 1%, and 0.5% (Ashrafi et al., 2008; Yu et al., 2003). In order to avoid nonrelevant results due to pH imbalance, the pH of the different concentrations was adjusted to 6.5 by adding 1(N) NaOH or 1(N) HCl, as necessary (Uddin & Robinson, 2017).
. Bioassay
Oryza sativa ‘Gobindobhog’ was chosen as a bioassay sample for this study. Healthy and uniform seeds were presoaked in distilled water for 2 hr, and then 10 seeds were randomly placed on two layers of filter paper in each sterilized Petri dish (9 cm). The filter papers were moistened with 10 mL of 5%, 4%, 2.5%, 1%, and 0.5% (w/v) aqueous leaf and root extracts and 10 mL of distilled water was used as a control treatment. Treatments were arranged in a completely randomized design with three replicates. The Petridishes were placed in a growth chamber (25 °C and 70% relative humidity) as suggested by Mutlu and Atici (2009). Seeds were considered germinated after 2 mm of radicle emergence (Haugland & Brandsaeter, 1996), and germination was determined by counting the number of germinated seeds over a period of 5 days.
. Data Collection
The following parameters were used to study the allelopathic effect during the experiment. The germination percentage of seeds was calculated using the following formula (Saxena et al., 1996):
The lengths of roots and shoots of seedlings were measured using a centimeter ruler. The seed vigor index (SVI) was calculated as suggested by Abdul-Baki and Anderson (1973):
where SL = shoot length, and RL = root length.
To calculate the percentage inhibitory effect on seed germination, root length, shoot length, and SVI, the following formula was used (Surendra & Pota, 1978):
where I = percentage inhibition, ET = response of test plant, and EC = response of control plant.
. Statistical Analysis
Analysis of variance (ANOVA) was used to test the significance of the treatment effects on seed bioassay, and an unpaired “t test” was used to determine whether a significant difference (p < 0.05) occurred between treatments and the control. Germination percentage, root, and shoot length at different concentration of the aqueous extracts of roots and leaves of two lamiaceous weeds were evaluated using linear regression analysis for the combined data set, and a heat map representation of the percentage inhibitory effects was generated using GraphPad Prism version 7.00 for Windows (GraphPad Software, California, USA). Regression analysis was performed by data analysis from all treatments in relation to seedling growth parameters and SVI.
. Results
. Effect of Aqueous Leaf and Root Extracts of H. suaveolens and L. cephalotes on Rice Seed Germination
The inhibitory response to aqueous extracts of H. suaveolens on seed germination differed between the leaf and root parts of the bioassay species. Seed germination was inhibited in all treatments, and a significant reduction in values (p < 0.01) was recorded for percentage seed germination at all concentrations of leaf and root extracts, except for 0.5% of aqueous root extracts (p < 0.05; Table 1), with germination declining progressively with increasing concentrations of aqueous leaf and root extracts. Five percent of aqueous leaf and root extracts reduced the seed germination of rice by 57.77% and 31.42%, respectively (Figure 1).
Table 1
Effect of aqueous leaf and root extracts from Hyptis suaveolens on seed germination and early growth of Oryza sativa ‘Gobindobhog.’
| Plant parts | Treatments (%) | Germination percentage (%) | Root length (cm) | Shoot length (cm) |
|---|---|---|---|---|
| Control | 98.66 ± 0.66 | 5.97 ± 0.02 | 6.08 ± 0.03 | |
| Leaf extracts | 0.5 | 92.33 ± 0.33c | 5.16 ± 0.17b | 5.36 ± 0.03c |
| 1.0 | 81.00 ± 1.73c | 4.54 ± 0.06d | 4.40 ± 0.02d | |
| 2.5 | 68.33 ± 0.88d | 4.13 ± 0.03d | 3.68 ± 0.02d | |
| 4.0 | 57.33 ± 1.45d | 3.24 ± 0.03d | 3.09 ± 0.02d | |
| 5.0 | 41.66 ± 1.66d | 2.05 ± 0.03d | 2.83 ± 0.02d | |
| Root extracts | 0.5 | 95.00 ± 1.15a | 5.57 ± 0.05b | 5.87 ± 0.01b |
| 1.0 | 87.00 ± 1.52b | 5.14 ± 0.03d | 5.12 ± 0.03d | |
| 2.5 | 80.00 ± 1.15c | 4.90 ± 0.02d | 4.88 ± 0.02d | |
| 4.0 | 76.00 ± 2.00c | 4.34 ± 0.04d | 4.77 ± 0.03d | |
| 5.0 | 67.66 ± 1.45d | 3.77 ± 0.03c | 4.11 ± 0.02d |
Figure 1
Heat map representation of percentage inhibitory effect on germination percentage (GP), root length (RL) and shoot length (SL) of different treatments of leaf and root extract from Hyptis suaveolens and Leucas cephalotes, compared to the control.
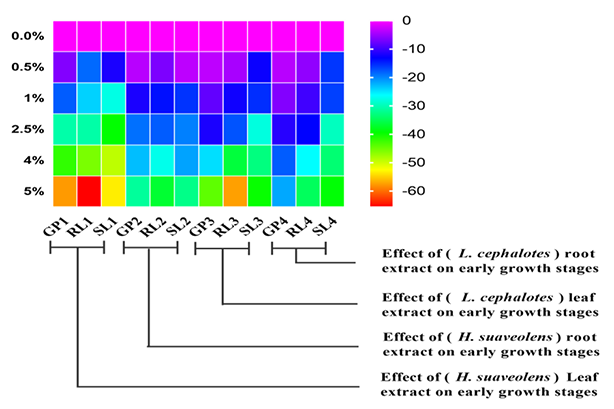
Leaf and root extracts of L. cephalotes significantly (p < 0.05) inhibited seed germination of rice as compared to the control (Table 2). The percentage inhibition was higher for leaf extracts than for root extracts and at increasing extract concentrations; at 5% concentration of aqueous leaf and root extracts it was 44.25% and 21.8%, respectively. The germination percentage declined with increasing concentrations of aqueous leaf and root extracts. The aqueous leaf extracts at low concentrations (0.5% and 1%) showed significant (p < 0.05) inhibitory effects of 3.37% and 8.10%, respectively, on rice germination, and the root extracts at 0.5% and 1% concentrations showed significant (p < 0.01) effects of 3.70% and 6.41%, respectively (Figure 1).
Table 2
Effect of aqueous leaf and root extracts from Leucas cephalotes on seed germination and early growth of Oryza sativa ‘Gobindobhog.’
| Plant parts | Treatments (%) | Germination percentage (%) | Root length (cm) | Shoot length (cm) |
|---|---|---|---|---|
| Control | 98.66 ± 0.66 | 5.97 ± 0.02 | 6.08 ± 0.03 | |
| Leaf extracts | 0.5 | 95.33 ± 0.33b | 5.70 ± 0.02b | 5.31 ± 0.03d |
| 1.0 | 90.66 ± 2.96a | 5.23 ± 0.04c | 5.14 ± 0.02d | |
| 2.5 | 87.00 ± 1.15c | 4.93 ± 0.02d | 4.38 ± 0.02d | |
| 4.0 | 74.33 ± 2.33c | 3.80 ± 0.05d | 4.08 ± 0.01d | |
| 5.0 | 55.00 ± 2.08d | 2.54 ± 0.06d | 3.66 ± 0.02d | |
| Root extracts | 0.5 | 95.00 ± 0.57b | 5.63 ± 0.03b | 5.11 ± 0.06b |
| 1.0 | 92.33 ± 1.45b | 5.38 ± 0.03d | 5.08 ± 0.01d | |
| 2.5 | 87.66 ± 1.45b | 5.18 ± 0.04c | 4.28 ± 0.01d | |
| 4.0 | 81.00 ± 0.57d | 4.40 ± 0.03d | 4.04 ± 0.01d | |
| 5.0 | 77.66 ± 2.18c | 3.90 ± 0.02d | 3.70 ± 0.02d |
. Effect of Aqueous Leaf and Root Extracts of H. suaveolens and L. cephalotes on Early Growth of Rice Seedlings
Leaf and root extracts at all concentrations exhibited significant (p < 0.01) inhibition in rice root and shoot length. After treating the seeds with aqueous leaf and root extracts of H. suaveolens, the root and shoot lengths of rice seedlings were significantly different from those in the controls (Table 1). Similarly, a gradual increase in the concentrations induced a significant decrease in the root and shoot lengths of rice seedlings. The maximum reduction of 65.66% in root length and 53.45% in shoot length was recorded after treatment with 5% aqueous leaf extracts from H. suaveolens (Figure 1). The 5% aqueous root extract showed a lower percentage inhibitory effect on root and shoot length (36.85% and 32.40%, respectively; Figure 1). Seed vigor indices significantly decreased (p < 0.01) compared to the control with increasing concentrations of leaf and root extracts from H. suaveolens (Figure 2).
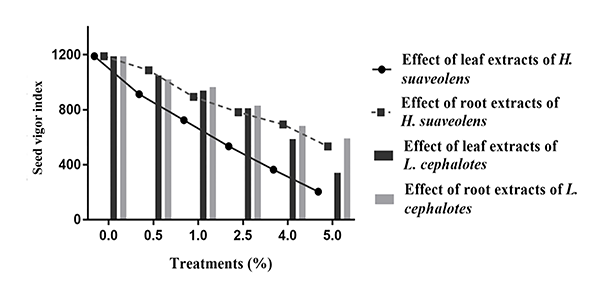
Figure 2
Effect of aqueous leaf and root extracts from Hyptis suaveolens and Leucas cephalotes on seed vigor indices.
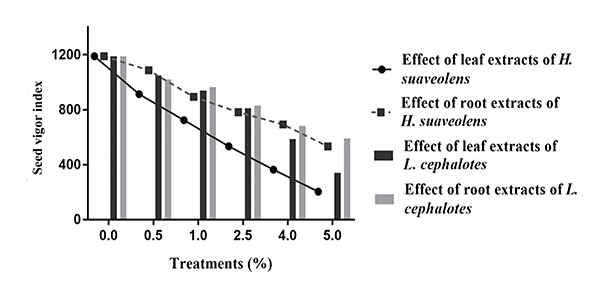
The aqueous leaf and root extracts at 5% concentrations showed an inhibitory effect on SVI (203.30 and 533.16, respectively; Figure 2). The value of SVI for the control treatment was 1,188.85.
Regression analysis revealed that under the aqueous leaf extracts, the rice SVIs were significantly related to germination percentage and root length (p < 0.001). Treatment with aqueous root extracts showed that SVIs were significantly related to root length (p < 0.001).
Rice shoot and root length differed significantly (p < 0.01) between controls and treatments with aqueous leaf and root extracts from L. cephalotes (Table 2). Compared to the control, aqueous extracts from L. cephalotes leaves showed a greater inhibitory effect on the rice root/shoot length than the L. cephalotes root extracts. At a lower concentration (0.5%), both leaf and root (L. cephalotes) extracts showed an inhibitory effect on rice root and shoot length. At 5% concentration, the rice root length was inhibited by 57.45% under L. cephalotes leaf aqueous extracts and by 34.67% under L. cephalotes root aqueous extracts, compared to the control (Figure 1). The shoot length was reduced by 39.80% and 39.14%, respectively.
Seed vigor indices significantly decreased (p < 0.001) compared to the control with increasing concentrations of leaf and root extracts from L. cephalotes (Figure 2). The SVIs in the treatments with L. cephalotes leaf extract were 585.72 (at 4% concentration) and 341 (at 5% concentration), while in the treatments with aqueous root extract, they were 683.64 (at 4% concentration) and 590 (at 5% concentration).
The regression analysis revealed that with aqueous leaf extracts, the SVIs were significantly related to germination percentage, root length, and shoot length (p < 0.001). Treatment with L. cephalotes aqueous root extracts showed that SVIs were significantly related to root length and shoot length (p < 0.001).
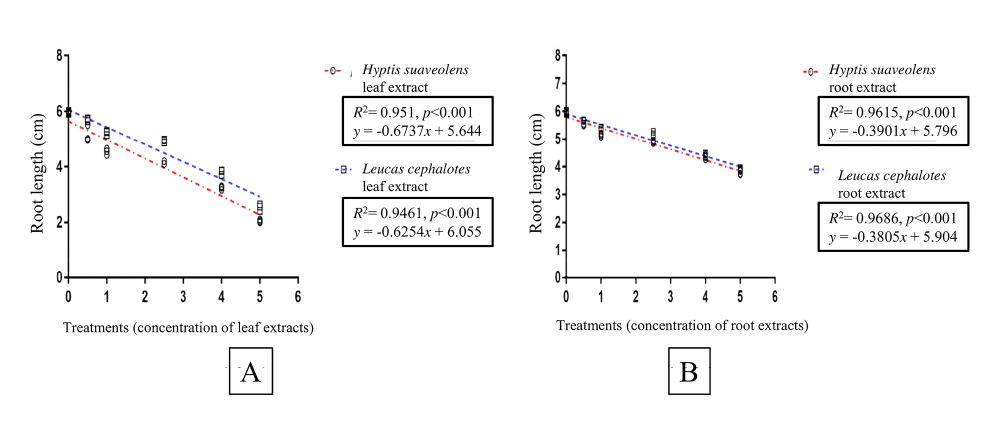
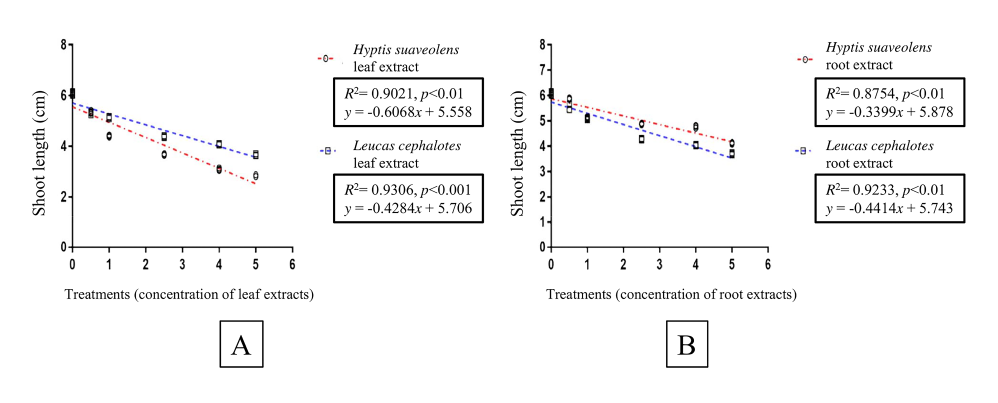
The regression analysis revealed that the H. suaveolens leaf extracts were responsible for 98% (R2 = 0.98), and the leaf extracts from L. cephalotes for 91% (R2 = 0.91) of the variation in rice seed germination (Figure 3A). The concentration of root extracts from H. suaveolens was responsible for 95% (R2 = 0.95), and from L. cephalotes for 99% (R2 = 0.99) of the variation in germination of the test species (Figure 3B). Regression analysis between treatments with leaf and root extracts from H. suaveolens or L. cephalotes showed that the root length of rice seedlings declined significantly (p < 0.001) with increasing concentration of extracts (Figure 4A,B). The shoot length of rice seedlings also declined significantly (p < 0.01) with increasing concentrations of aqueous leaf and root extracts from H. suaveolens and L. cephalotes (Figure 5A,B). A high reduction was indicated by a higher slope of the regression line in all cases.
Figure 3
Linear regression analysis showing variation in seed germination percentage with concentration of aqueous leaf (A) and root (B) extracts from Hyptis suaveolens and Leucas cephalotes.
. Discussion
The results of allelopathic studies thus far are inconsistent in the context of the stimulatory or inhibitory effects of allelopathic plants (Yasmeen et al., 2013). The present study demonstrated that the aqueous leaf and root extracts from H. suaveolens and L. cephalotes inhibited seed germination and seedling growth of Oryza sativa ‘Gobindobhog.’
Allelopathic activity depends on the concentration and varies from species to species (Assaeed & Al-Doss, 1997; Bari & Kato-Noguchi, 2017). In addition, Sundaramoorthy & Sen (1990) reported that leachates obtained from different plant parts of Tephrosia purpurea differentially affected the growth of various crops because of differences in their allelochemical diversity. In many allelopathic studies, high concentrations of extracts generally have an inhibitory effect on seed germination and seedling growth (Rice, 1984; Synowiec & Nowicka-Połeć, 2016). Prinsloo and Du Plooy (2018) have also suggested that the allelopathic effect depends on the concentration and type of test species. For example, these authors reported that high concentrations of Amaranthus extracts significantly inhibited tomato seed germination; however, at low concentrations no such effect was observed. In contrast, low concentrations of allelochemicals can have a stimulatory effect on target species (Lovett et al., 1989); however, the present study revealed that the effect of aqueous leaf and root extracts of two lamiaceous species at low concentrations (0.5% and 1%) showed inhibitory effects on seed germination and the early growth parameters of rice seedlings. Our results support those of Poornima et al. (2015). Presumably, the mitotic activity of cells is suppressed by plant allelochemicals, which impedes the initial development of cells and consequently inhibits seed germination (Rice, 1985).
Leaves have been reported to contain more allelochemicals than other plant organs (Dorning & Cipollini, 2006). In nature, allelochemicals are mostly water-soluble (Turk et al., 2003), and almost all allelochemicals in the above-ground parts of plants are leached out into the soil (Wetzel & Howe, 1999). For example, aqueous leaf extracts of allelopathic black mustard (Brassica nigra) have a greater effect on root length than shoot length in Avena fatua (Turk & Tawaha, 2003). Generally, roots absorb allelochemicals from moist filter paper owing to their direct contact and are more affected by different concentrations than shoots (Ismail & Chong, 2002; Turk & Tawaha, 2003). The present study revealed that aqueous leaf extracts of H. suaveolens and L. cephalotes showed the highest inhibitory effect on rice root length compared to other growth parameters. Similar results were reported by Mominul Islam and Kato-Noguchi (2013b). In mustard seedlings, roots were more affected by the aqueous extracts of Helianthus annuus than by shoots (Ghafar et al., 2001).
Increasing the concentrations of aqueous leaf and root extracts increased the inhibitory effect, especially on shoot length. Similar results for the allelopathic action of Nepeta meyeri on shoot length were reported by Mutlu and Atici (2009). Tefera (2002) also reported that higher concentrations of aqueous leaf extract from Parthenium hysterophorus showed a high percentage inhibition on shoot length of teff seedlings (Poaceae), whereas under lower aqueous leaf extract concentrations, a stimulatory effect was observed.
The standard germination test does not perfectly represent the field performance of seeds. Therefore, many researchers recommend seed vigor testing. In the control treatments, high SVIs were observed, which indicated normal seed germination, as might be expected in normal field conditions. Under higher concentrations of aqueous extracts, the SVIs were poor, which may indicate that seedlings may not survive in the field. Lower seed vigor was observed with aqueous leaf extracts of H. suaveolens than with aqueous leaf extracts of L. cephalotes. The present findings are in accordance with Das et al. (2012), who found that the aqueous leachates of leaf litter of seven tree species significantly (p < 0.01) reduced the SVI of gram (chickpea) seeds. Alqarawi et al. (2018) reported a strong inhibitory effect of aqueous leaf extracts of Rhazya stricta on SVI of Salsola villosa, while Oraon and Mondal (2020) reported that the SVI of chickpea decreased significantly (p < 0.001) with increased concentrations of aqueous leaf extracts of Putranjiva roxburghii.
The effect was more pronounced with leaf extracts than with root extracts, indicating that the leaf contains many allelochemicals that can act together as inhibitory agents. The effect of allelochemicals from various plant parts cannot be attributed to a single chemical compound. If allelochemicals act as a mixture; therefore, understanding the allelopathic interaction between plants in their environment is not easy (Inderjit, 2005). The allelopathic potential of weeds can adversely affect the establishment of native species.
Aqueous leaf and root extracts of H. suaveolens showed greater allelopathic effects than leaf and root extracts of L. cephalotes on seed germination and early growth of aromatic Oryza sativa ‘Gobindobhog.’ Both the weeds showed inhibitory effects on seed germination and early growth, and this should be taken into consideration prior to cultivation of Oryza sativa ‘Gobindobhog.’
